


Deciphering Winter Sprouting Potential of Erianthus procerus Derived Sugarcane Hybrids under Subtropical Climates

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Weather Statistics of the Experimental Site
2.2. Significance of Variation Sources
2.3. Mean Performance
2.3.1. Agro-Morphological and Quality Traits
2.3.2. Physiological Traits
Flavonoid and Anthicyanin Indices
Exploring Fibre Content in Procerus-Derived Hybrids
2.4. Correlation Studies
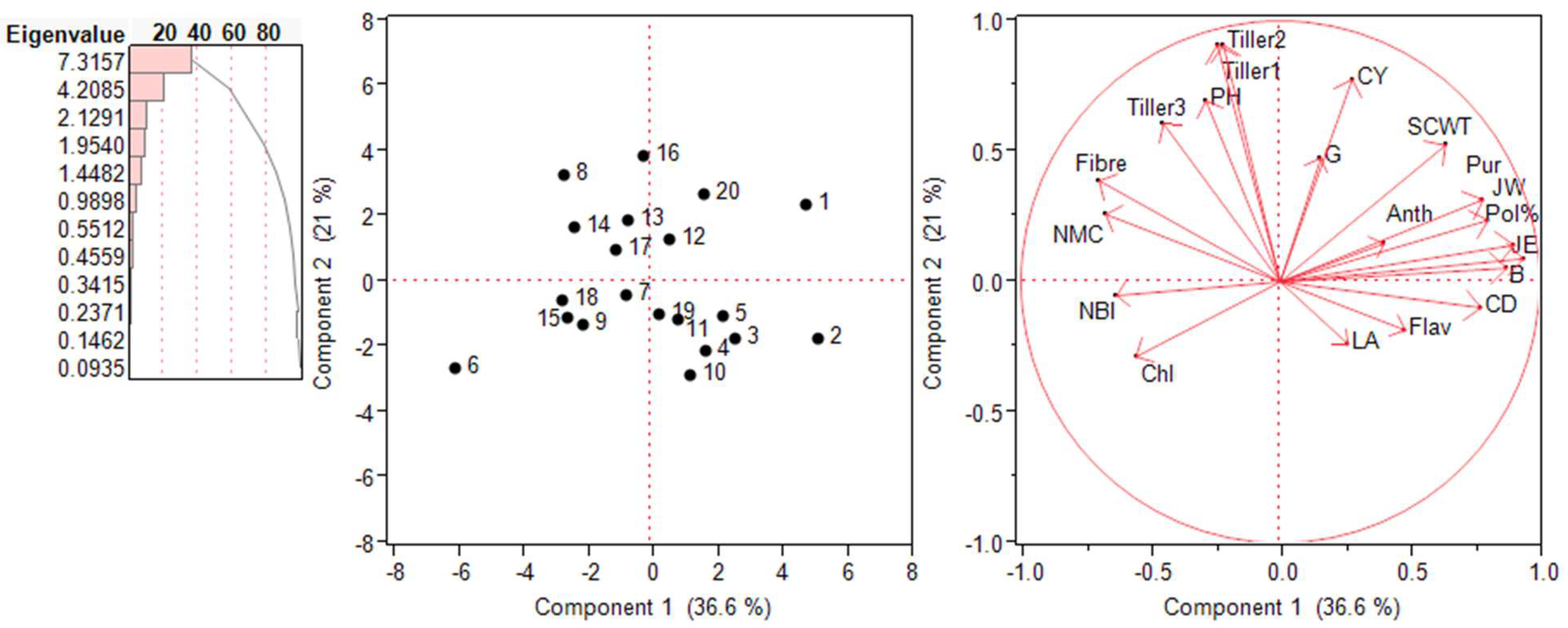
2.5. Principal Component Analysis (PCA)—Biplot Analysis
2.6. Winter Sprouting and Red Rot
2.6.1. Winter Sprouting Potential in Erianthus Procerus Vis-a-Vis Standard Checks
2.6.2. Resistance of Erianthus procerus-Derived Hybrids against the Red Rot
3. Discussion
3.1. Variability and Mean Performance for Various Economic Traits
3.2. Winter Sprouting Potential
3.3. Red Rot Resistance
3.4. Fibre and Flavonoids in Erinathus procerus-Derived Hybrids
4. Materials and Methods
4.1. Experimental Site
4.2. Plant Material and Experimental Design
4.3. Red Rot Evaluation
4.4. Measurement of Physiological Parameters
4.5. Confirmation of Intergeneric Hybrids
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Glossary
| Term | Details |
| PCA: Principal component analysis | Statistical method used for reducing the dimensionality of large datasets into smaller ones |
| Juice extraction (%) | Percentage of juice from a given quantity of cane by mechanical extraction |
| Brix value (%) | Measurement of total dissolved solids content in a solution |
| Pol (%) | Measurement of sucrose content in percent |
| Purity (%) | Sucrose content as ratio of the total dissolved solids and expressed in percent |
| Fibre content (%) | Amount of fibre present in sugarcane |
| FLAV: Flavonoid index | A diverse group of naturally occurring compounds in sugarcane |
| ANTH: Anthocyanin index | Quantitative assessments of anthocyanins in a sample |
| SCWT: Single cane weight (kg) | Weight of an individual mature sugarcane stalk |
| Ger: Germination (%) | Percentage of cane bud setts in a given sample that have successfully begun to emerge out |
| NBI: Nitrogen balance index | The ratio between chlorophyll and flavonoid contents and indicator of crop N status for crop growth |
| 60-day tillers (‘000/ha) | Population of tillers after 60 days of a specific crop’s growth |
| 90-day tillers (‘000/ha) | Population of tillers after 90 days of a specific crop’s growth |
| 120-day tillers (‘000/ha) | Population of tillers after 120 days of a specific crop’s growth |
| PH: Plant height (cm) | Vertical growth of cane from the base to the top of the stem, where it easily breaks by hand |
| CD: Cane diameter (cm) | Measurement of the diameter of a cane stalk |
| NMCs: Number of millable canes (‘000/ha) | The population of mature cane which is ready to crush in the sugar factory |
| Leaf area (cm2) | The surface area of a leaf and expressed in square centimetres. Leaf area (cm2) = length (cm) × width (cm) |
| CHLORO: Chlorophyll (µg/cm2) | Amount of chlorophyll present in a unit area of a cane leaf surface |
| CY: Cane yield (t/ha) | Amount of sugarcane (in weight) harvested from a specific area |
| WSI: Winter sprouting index | WSI = percentage of sprouted clumps × number of shoots per clumps/100 |
| SPAD: Soil plant analysis development | Measurement of the relative chlorophyll content in leaves in a non-destructive way |
| TVD: Top visual dewlap | Uppermost fully expanded leaf with visible dewlap or distinct collar, a band between the leaf sheath and leaf blade |
| PCR: Polymerase chain reaction | Used to amplify a specific segment of DNA in millions to billions of copies |
| UDL: Upper decision limit | The threshold set in a measurement which indicates the maximum acceptable value for a particular parameter |
| LDL: Lower decision limit | The threshold set in a measurement which indicates the minimum acceptable value for a particular parameter |
| BC: Back-cross | Mating of a hybrid organism (offspring of genetically unlike parents) with one of its parents |
References
- Manochio, C.; Andrade, B.R.; Rodriguez, R.P.; Moraes, B.S. Ethanol from biomass: A comparative overview. Renew. Sustain. Energy Rev. 2017, 80, 743–755. [Google Scholar] [CrossRef]
- Kim, M.; Day, D.F.L. Composition of sugarcane, energy cane, and sweet sorghum suitable for ethanol production at Louisiana sugar mills. J. Ind. Microbiol. Biotechnol. 2011, 38, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Chatwachirawong, P.; Thumkrasair, S.; Srisink, S. Sugarcane breeding final report: Research development design and engineering project BT-B-01-PG-11-4924. NSTDA 2009. [Google Scholar]
- Cursi, D.E.; Hoffmann, H.P.; Barbosa, G.V.S.; Bressiani, J.A.; Gazaffi, R.; Chapola, R.G.; Fernandes, J.A.R.; Balsalobre, T.W.A.; Diniz, C.A.; Santos, J.M. History and current status of sugarcane breeding, germplasm development and molecular genetics in Brazil. Sugar Tech. 2022, 24, 112–133. [Google Scholar] [CrossRef]
- Govindaraj, P.; Amalraj, V.A.; Mohanraj, K.; Nair, N.V. Collection, characterization and phenotypic diversity of Saccharum spontaneum L. from arid and semiarid zones of northwestern India. Sugar Tech. 2014, 16, 36–43. [Google Scholar] [CrossRef]
- Mukherjee, S.K. Origin and distribution of Saccharum. Bot. Gaz. 1957, 119, 55–61. [Google Scholar] [CrossRef]
- D’hont, A.; Rao, P.; Feldmann, P.; Grivet, L.; Islam-Faridi, N.; Taylor, P.; Glaszmann, J.C. Identification and characterization of sugarcane intergeneric hybrids, Saccharum officinarum × Erianthus arundinaceus, with molecular markers and DNA in situ hybridization. Theor. Appl. Genet. 1995, 91, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Aitken, K.; Deng, H.; Chen, X.W.; Fu, C.; Jackson, P.A.; McIntyre, C.L. Verification of the introgression of Erianthus arundinaceus germplasm into sugarcane using molecular markers. Plant Breed. 2005, 124, 322–328. [Google Scholar] [CrossRef]
- Fukuhara, S.; Terajima, Y.; Irei, S.; Sakaigaichi, T.; Ujihara, K. Identification and characterization of intergeneric hybrid of commercial sugarcane (Saccharum spp. hybrid) and Erianthus arundinaceus (Retz.) Jeswiet. Euphytica 2013, 189, 321–327. [Google Scholar] [CrossRef]
- Mohanraj, K.; Nair, N.V. Biomass potential of novel interspecific hybrids involving improved clones of Saccharum. Ind. Crops Prod. 2014, 53, 128–132. [Google Scholar] [CrossRef]
- Amalraj, V.A.; Balasundaram, N. On the taxonomy of the members of “Saccharum complex”. Genet. Resour. Crop Evol. 2006, 53, 35–41. [Google Scholar] [CrossRef]
- Nair, N.V.; Mohanraj, K.; Sunadaravelpandian, K.; Suganya, A.; Selvi, A.; Appunu, C. Characterization of an intergeneric hybrid of Erianthus procerus × Saccharum officinarum and its backcross progenies. Euphytica 2017, 213, 267. [Google Scholar] [CrossRef]
- Verma, R.S. Sugarcane Ratoon Management; International Book Distributing Co.: Lucknow, India, 2002; 266p. [Google Scholar]
- Ram, B.; Karuppaiyan, R.; Meena, M.R.; Kumar, R.; Kulshreshtha, N. Winter sprouting index of sugarcane genotypes is a measure of winter ratooning ability. Int. J. Dev. Res. 2017, 7, 15385–15391. [Google Scholar]
- Hemaprabha, G. Sugarcane Varieties for Abiotic Stress Tolerance. In Training manual on “Sugarcane Cultivation in Biotic and Abiotic Stress Conditions”; Nair, V., Gopalasundaram, P., Shanty, T.R., Prathap, P.D., Eds.; ICAR-Sugarcane Breeding Institute: Tamil Nadu, India, 2008. [Google Scholar]
- Tsuruta, S.I.; Srithawong, S.; Sakuanrungsirikul, S.; Ebina, M.; Kobayashi, M.; Terajima, Y.; Tippayawat, A.; Ponragdee, W. Erianthus germplasm collection in Thailand: Genetic structure and phylogenetic aspects of tetraploid and hexaploid accessions. BMC Plant Biol. 2022, 22, 45. [Google Scholar] [CrossRef]
- Kandel, R.; Yang, X.; Song, J.; Wang, J. Potentials, challenges, and genetic and genomic resources for sugarcane biomass improvement. Front. Plant Sci. 2018, 9, 151. [Google Scholar] [CrossRef]
- Meena, M.R.; Kumar, R.; Ramaiyan, K. Biomass potential of novel interspecific and intergeneric hybrids of Saccharum grown in subtropical climates. Sci. Rep. 2020, 10, 21560. [Google Scholar] [CrossRef]
- Medeiros, K.L.S.; Reis, V.R.; Santos, J.R.M. Evaluation of biomass productivity and energy potential of energy cane varieties in Brazil. Ind. Crops Prod. 2019, 140, 111625. [Google Scholar] [CrossRef]
- Li, S.J.; Bai, Y.C.; Li, C.L.; Yao, H.P.; Chen, H.; Zhao, H.X.; Wu, Q. Anthocyanins accumulate in tartary buckwheat (Fagopyrum tataricum) sprout in response to cold stress. Acta Physiol. Plantarum. 2015, 37, 159. [Google Scholar] [CrossRef]
- Pachakkil, B.; Terajima, Y.; Ohmido, N.; Ebina, M.; Irei, S.; Hayashi, H.; Takagi, H. Cytogenetic and agronomic characterization of intergeneric hybrids between Saccharum spp. hybrid and Erianthus arundinaceus. Sci. Rep. 2019, 9, 1748. [Google Scholar] [CrossRef]
- Sobhakumari, V.P.; Mohanraj, K.; Nair, N.V.; Mahadevaswamy, H.K.; Ram, B. Cytogenetic and molecular approaches to detect alien chromosome introgression and its impact in three successive generations of Erianthus procerus × Saccharum. Cytologia 2020, 85, 341–346. [Google Scholar] [CrossRef]
- Parihar, R. Character association and path coefficient analysis for cane yield and quality characters in fourth clonal generation (C4) of Sugarcane (Saccharum sp. complex). J. Crop. Weed. 2020, 16, 256–260. [Google Scholar] [CrossRef]
- Doule, R.B.; Balasundaram, N. Genetic variability in fibre and related characters for selection of sugarcane. Sugar Tech. 2004, 6, 73–75. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, D. Correlation and Path Coefficient Analysis in Sugarcane Germplasm under Subtropics. Afr. J. Agri. Res. 2014, 9, 148–153. [Google Scholar] [CrossRef]
- Kohler, U.; Luniak, M. Data inspection using biplots. Stata J. 2005, 5, 208–223. [Google Scholar] [CrossRef]
- Kumar, R.; Meena, M.R.; Dhansu, P.; Karuppaiyan, R.; Appunu, C.; Kulshreshtha, N.; Kaushik, P.; Ram, B. Winter Tolerance Potential of Genetically Diverse Sugarcane Clones under Subtropical Climate of Northern India. Sustainability 2022, 14, 11757. [Google Scholar] [CrossRef]
- Jain, R.; Shrivastava, A.K.; Solomon, S.; Yadav, R.L. Low temperature stress-induced biochemical changes affect bud sprouting in sugarcane (Saccharum spp. hybrid). Plant Growth Regul. 2007, 53, 17–23. [Google Scholar] [CrossRef]
- Sahi, B.K.; Ram, B.; Kumar, P. Evaluation of sugarcane clones for ratoonability during winter month. Indian J. Sugarcane Technol. 2002, 17, 1–4. [Google Scholar]
- Sujie, L.I.; Xianhong, W.; Qinghui, Y. Evaluation of Cold Tolerance in Wild Sugarcane Germplasm Resources under Field Conditions. Agric. Biotechnol. 2017, 6, 2164–4993. [Google Scholar]
- Ram, B.; Sreenivasan, T.; Sahi, B.; Singh, N. introgression of low temperature tolerance and red rot resistance from Erianthus in sugarcane. Euphytica 2001, 122, 145–153. [Google Scholar] [CrossRef]
- Viswanathan, R. Red rot of sugarcane (Colletotrichum falcatum Went). CAB Rev. 2021, 16, 23–57. [Google Scholar] [CrossRef]
- Mohanraj, K.S.A. Broadening the genetic base of sugarcane: Introgression of red rot resistance from the wild relative Erianthus procerus. In Proceedings of the XII International Society of Sugar Cane Technologists Pathology Workshop, Coimbatore, India, 3–7 September 2018; ICAR-SBI: Coimbatore, India, 2018; Volume 18. [Google Scholar]
- Viswanathan, R. Severe Red Rot Epidemics in Sugarcane in Sub-tropical India: Role of Aerial Spread of the Pathogen. Sugar Tech. 2023, 25, 1275–1277. [Google Scholar] [CrossRef]
- Srinivasan, K.V.B.N.R. Red rot of sugarcane—Criteria for grading resistance. J. Indian Bio. Soc. 1961, 40, 566–577. [Google Scholar]
- Buckeridge, M.S.; De Souza, A.P.; Tavares, E.Q.P.; Cambler, A.B. Sugarcane Cell Wall Structure and Degradation: From Monosaccharide Analyses to the Glycomic Code. In Advances of Basic Science for Second Generation Bioethanol from Sugarcane; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 7–19. [Google Scholar] [CrossRef]
- Carvalho-Netto, O.V.; Bressiani, J.A.; Soriano, H.L.; Fiori, C.S.; Santos, J.M.; Barbosa, G.V.; Xavier, M.A.; Landell, M.G.A.; Pereira, G.A.G. The potential of the energy cane as the main biomass crop for the cellulosic industry. Chem. Biol. Technol. Agric. 2014, 1, 20. [Google Scholar] [CrossRef]
- Dias, M.O.S.; Junqueira, T.L.; Cavalett, O.; Cunha, M.P.; Jesus, C.D.F.; Rossell, C.E.V.; Maciel Filho, R.; Bonomi, A. Integrated versus stand-alone second generation ethanol production from sugarcane bagasse and trash. Bioresour. Technol. 2012, 103, 152–161. [Google Scholar] [CrossRef]
- Hoang, N.V.; Furtado, A.; Botha, F.C.; Simmons, B.A.; Henry, R.J. Potential for genetic improvement of sugarcane as a source of biomass for biofuels. Front. Bioeng. Biotechnol. 2015, 3, 182. [Google Scholar] [CrossRef] [PubMed]
- Colombo, R.; Lanças, F.M.; Yariwake, J.H. Determination of flavonoids in cultivated sugarcane leaves, bagasse, juice and in transgenic sugarcane by liquid chromatography-UV detection. J. Chromatogr. A 2006, 1103, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Sun, Q.; Li, H.; Li, X.; Cao, Y.; Zhang, H.; Li, S.; Zhang, L.; Qi, Y.; Ren, S.; et al. Melatonin improved anthocyanin accumulation by regulating gene expressions and resulted in high reactive oxygen species scavenging capacity in cabbage. Front. Plant Sci. 2016, 7, 197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.Q.; Yang, L.T.; Li, Y.R. Physiological and biochemical characteristics related to cold resistance in sugarcane. Sugar Tech. 2015, 17, 49–58. [Google Scholar] [CrossRef]
- Chen, J.C.; Chou, C.C. Cane Sugar Handbook; Wiley: Hoboken, NJ, USA, 1993. [Google Scholar]
- Thangavelu, S.C.R.K. Comparison of Rapi pol extractor and Cutex cane shredder methods for direct determination of fibre in Saccharum clones. Proc. Annu. Conv. Sugar Technol. Assoc. India. 1982, 46, 15–21. [Google Scholar]
- LeBauer, D.; Newcomb, M.; Herrit, M.; Babaeian, E. Flavonol, Anthocyanin, and Chlorophyll Indices Using a Force-A Dualiex Scientific+. 2019. Available online: https://web.archive.org/web/20200303042750id_/https://www.protocols.io/view/flavonol-anthocyanin-and-chlorophyll-indices-using-u6yezfw.pdf (accessed on 23 November 2019).
- Meade, G.P.; Chen, J.C.P. Cane Sugar Hand Book, 10th ed.; John Wiley & Sons: New York, NY, USA, 1977. [Google Scholar]
- Babu, C.; Koodalingam, K.; Natarajan, U.; Shanthi, R.; Govindaraj, P. Genetic enhancement of sugarcane (Saccharum sp. hybrids) for resistance to red rot disease and economic traits. J. Agric. Sci. 2010, 4, 97. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; Wiley: Hoboken, NJ, USA, 1984. [Google Scholar]
- SAS Institute. The SAS System for Windows; Release 9.3; SAS Institute: Independence, KS, USA, 2011. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Main Effects | Interaction Effects | ||||
|---|---|---|---|---|---|---|
| Genotypes (19) | Years (1) | Genotypes × Years (19) | ||||
| MSS | p Value | MSS | p Value | MSS | p Value | |
| Germination | 216.4 | 0.0145 * | 15,359.9 | <0.0001 ** | 121.2 | 0.3211 |
| 60-day tillers | 9.47 × 108 | <0.0001 ** | 2.61 × 109 | 0.1845 | 3.18 × 108 | 0.9997 |
| 90-day tillers | 3.21 × 109 | <0.0001 ** | 1.11 × 1011 | <0.0001 ** | 2.02 × 109 | 0.0003 ** |
| 120-day tillers | 3.30 × 109 | 0.0005 ** | 2.16 × 1011 | <0.0001 ** | 2.72 × 109 | 0.0034 ** |
| Plant height | 9256.8 | <0.0001 ** | 0.0 | 1.000 | 0.0 | 1.000 |
| Cane diameter | 0.7 | <0.0001 ** | 0.0 | 1.000 | 0.0 | 1.000 |
| Juice weight | 2.5 | <0.0001 ** | 0.0 | 1.000 | 6.95 × 1029 | 1.000 |
| Juice extraction | 397.7 | <0.0001 ** | 0.0 | 1.000 | 4.21 × 1029 | 1.000 |
| Brix value | 15.1 | <0.0001 ** | 6.8 | 0.0755 | 1.4 | 0.8373 |
| Pol% | 21.9 | <0.0001 ** | 13.7 | 0.0272 * | 2.5 | 0.5523 |
| Purity | 105.2 | <0.0001 ** | 64.7 | 0.0969 | 24.5 | 0.4001 |
| Single cane weight | 0.2 | <0.0001 ** | 0.01 | 0.6407 | 0.02 | 0.3455 |
| Fibre content | 49.9 | <0.0001 ** | 45.6 | 0.0142 * | 16.1 | 0.0076 ** |
| Number of millable canes | 1.42 × 109 | <0.0244 * | 4.50 × 108 | 0.4372 | 2.93 × 108 | 0.9871 |
| Cane yield | 1495.1 | <0.0001 ** | 689.8 | 0.1095 | 97.5 | 0.9913 |
| Nitrogen balance index | 213.7 | <0.0001 ** | 70.9 | 0.1433 | 33.6 | 0.4334 |
| Chlorophyll concentration | 128.3 | <0.0001 ** | 42.5 | 0.2199 | 26.5 | 0.5204 |
| Flavonoid index | 0.15 | 0.1072 | 0.2 | 0.1474 | 0.11 | 0.3385 |
| Anthocyanin index | 0.0008 | 0.2709 | 0.0004 | 0.4417 | 0.0006 | 0.5011 |
| Leaf area | 3020.1 | 0.748 | 1035.2 | 0.6119 | 1618.4 | 0.9852 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1.00 | 0.066 | −0.45 ** | −0.59 ** | −0.01 | 0.12 | 0.17 | 0.21 | 0.17 | 0.06 | 0.05 | −0.00 | −0.37 * | −0.10 | −0.08 | −0.20 | −0.17 | 0.23 | −0.13 | −0.18 |
| 2 | 1.00 | 0.57 ** | 0.40 * | 0.66 ** | −0.28 | 0.06 | −0.09 | −0.02 | 0.02 | 0.12 | 0.45 ** | 0.32 * | 0.04 | 0.57 ** | −0.04 | −0.18 | −0.24 | 0.02 | −0.05 | |
| 3 | 1.00 | 0.88 ** | 0.29 | −0.15 | 0.00 | −0.03 | 0.07 | 0.122 | 0.215 | 0.149 | 0.356 * | 0.295 | 0.395 * | 0.054 | −0.090 | −0.168 | 0.121 | −0.007 | ||
| 4 | 1.00 | 0.207 | −0.19 | −0.09 | −0.14 | 0.02 | 0.03 | 0.01 | 0.00 | 0.33 * | 0.32 * | 0.23 | 0.14 | 0.08 | −0.21 | 0.11 | −0.08 | |||
| 5 | 1.00 | −0.14 | 0.13 | −0.27 | −0.31 * | −0.20 | −0.09 | 0.43 ** | 0.43 ** | −0.01 | 0.51 ** | 0.32 * | 0.26 | −0.25 | −0.02 | −0.22 | ||||
| 6 | 1.00 | 0.85 ** | 0.81 ** | 0.50 ** | 0.51 ** | 0.47 ** | 0.28 | −0.41 ** | −0.18 | 0.20 | −0.20 | −0.17 | 0.09 | 0.12 | 0.16 | |||||
| 7 | 1.00 | 0.86 ** | 0.66 ** | 0.67 ** | 0.58 ** | 0.50 ** | −0.34 * | −0.27 | −0.36 * | −0.19 | −0.15 | 0.00 | 0.07 | 0.08 | ||||||
| 8 | 1.00 | 0.77 ** | 0.79 ** | 0.70 ** | 0.44 ** | −0.50 ** | −0.32 * | 0.27 | −0.40 * | −0.37 * | 0.12 | 0.17 | 0.12 | |||||||
| 9 | 1.00 | 0.97 ** | 0.77 * | 0.29 | −0.57 ** | −0.37 * | 0.07 | −0.35 * | −0.27 | 0.09 | 0.09 | 0.06 | ||||||||
| 10 | 11.00 | 0.89 ** | 0.34 * | −0.54 ** | −0.34 * | 0.17 | −0.35 * | −0.31 | 0.09 | 0.14 | 0.09 | |||||||||
| 11 | 1.00 | 0.40 * | −0.36 * | −0.21 | 0.35 * | −0.29 | −0.36 * | 0.08 | 0.021 | 0.13 | ||||||||||
| 12 | 1.00 | 0.123 | −0.53 ** | 0.84 ** | −0.26 | −0.29 | −0.00 | 0.33 * | 0.07 | |||||||||||
| 13 | 1.00 | 0.35 * | 0.35 * | 0.34 * | 0.19 | −0.23 | −0.10 | 0.08 | ||||||||||||
| 14 | 1.00 | −0.01 | 0.26 | 0.04 | 0.00 | −0.12 | −0.11 | |||||||||||||
| 15 | 1.00 | −0.10 | −0.27 | −0.01 | 0.24 | 0.05 | ||||||||||||||
| 16 | 1.00 | 0.84 ** | −0.47 ** | −0.57 ** | −0.01 | |||||||||||||||
| 17 | 1.00 | −0.19 | −0.52 ** | 0.02 | ||||||||||||||||
| 18 | 1.00 | 0.31 * | −0.09 | |||||||||||||||||
| 19 | 1.00 | −0.15 | ||||||||||||||||||
| 20 | 1.00 |
| SN | Genotype | Winter Sprouting Index (WSI) | |||
|---|---|---|---|---|---|
| 2018–2019 | 2019–2020 | 2020–2021 | Mean | ||
| 1 | GU 12—19 | 8.0 | 12.2 | 11.6 | 10.6 |
| 2 | GU 12—22 | 6.1 | 9.9 | 9.5 | 8.5 |
| 3 | GU 12—21 | 5.3 | 7.2 | 9.1 | 7.2 |
| 4 | GU 12—29 | 3.5 | 8.8 | 8.4 | 6.9 |
| 5 | GU 12—33 | 4.0 | 8.6 | 7.3 | 6.6 |
| 6 | GU 12—23 | 4.8 | 7.3 | 6.4 | 6.2 |
| 7 | GU 12—27 | 3.9 | 8.7 | 5.5 | 6.0 |
| 8 | GU 12—34 | 4.2 | 6.9 | 6.3 | 5.8 |
| 9 | GU 04(28)EO—2 | 6.0 | 6.0 | 5.3 | 5.8 |
| 10 | GU 12—30 | 5.0 | 5.4 | 6.5 | 5.6 |
| 11 | GU 12—31 | 3.1 | 5.3 | 6.9 | 5.1 |
| 12 | GU 12—16 | 3.7 | 5.5 | 5.3 | 4.8 |
| 13 | GU 12—38 | 2.5 | 4.1 | 5.7 | 4.1 |
| 14 | GU 12—26 | 2.9 | 4.2 | 3.9 | 3.7 |
| 15 | GU 12—28 | 2.9 | 4.0 | 3.3 | 3.4 |
| 16 | Co 0238 (Check) | 2.4 | 3.6 | 2.5 | 2.8 |
| 17 | CoJ 64 (Check) | 1.7 | 2.9 | 2.3 | 2.3 |
| 18 | Co 06027 (Check) | 0.5 | 3.1 | 2.0 | 1.9 |
| 19 | CoS 8436 (Check) | 1.0 | 2.5 | 1.7 | 1.7 |
| 20 | CoS 767 (Check) | 0.9 | 1.9 | 1.4 | 1.4 |
| Erinathus procerus | 2018–2019 and 2019–2020 | 2018–2019 and 2019–2020 | ||
|---|---|---|---|---|
| Coimbatore (Tropical Isolates) | Karnal (Subtropical Isolates) | |||
| Clone name | Cf671 | Cf671 + Cf94012 | Cf08 | Cf09 |
| GU04 (28) EO-2 | R | R | MR | MR |
| GU12—16 | MR | R | S | MR |
| GU12—19 | R | MR | MR | MR |
| GU12—21 | R | R | MR | R |
| GU12—22 | MR | MR | MR | MR |
| GU12—23 | MR | R | - | MS |
| GU12—26 | MR | MR | MR | MR |
| GU12—27 | MR | R | MR | R |
| GU12—28 | MR | MR | S | MR |
| GU12—29 | MR | R | S | MR |
| GU12—30 | MR | MR | MR | MR |
| GU12—31 | MR | R | MR | MR |
| GU12—33 | R | MR | MS | MR |
| GU12—34 | R | MR | MS | S |
| GU12—38 | S | HS | S | S |
| Co 06027 | MR | S | S | MR |
| SN | Clone | Generation | Female | Male | Remarks |
|---|---|---|---|---|---|
| 1 | GU 04(28) EO—2 | F1 | IND 90-776 | PIO 96-435 | GU 04 (28) EO—2 is an intergeneric hybrid between Erianthus procerus and S. officinarum |
| 2 | GU 12—16 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 3 | GU 12—19 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 4 | GU 12—21 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 5 | GU 12—22 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 6 | GU 12—23 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 7 | GU 12—26 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 8 | GU 12—27 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 9 | GU 12—28 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 10 | GU 12—29 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 11 | GU 12—30 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 12 | GU 12—31 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 13 | GU 12—33 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 14 | GU 12—34 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 15 | GU 12—38 | BC1 | GU 04(28) EO-2 | Co 06027 | BC1 hybrid |
| 16 | Co 06027 | - | CoC 671 | IG 91-1100 | Commercial Variety Tropical |
| 17 | Co 0238 | - | Co LK 8102 | Co 775 | Commercial Variety Subtropical |
| 18 | CoJ 64 | - | Co 976 | Co 617 | Commercial Variety Subtropical |
| 19 | CoS 767 | - | Co 419 | Co 319 | Commercial Variety Subtropical |
| 20 | CoS 8436 | - | MS 68/47 | Co 1148 | Commercial Variety Subtropical |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meena, M.R.; Mohanraj, K.; Kumar, R.; Kumar, R.A.; Chhabra, M.L.; Kulshreshtha, N.; Krishnappa, G.; Swamy, H.K.M.; Suganya, A.; Govindaraj, P.; et al. Deciphering Winter Sprouting Potential of Erianthus procerus Derived Sugarcane Hybrids under Subtropical Climates. Plants 2024, 13, 1023. https://doi.org/10.3390/plants13071023
Meena MR, Mohanraj K, Kumar R, Kumar RA, Chhabra ML, Kulshreshtha N, Krishnappa G, Swamy HKM, Suganya A, Govindaraj P, et al. Deciphering Winter Sprouting Potential of Erianthus procerus Derived Sugarcane Hybrids under Subtropical Climates. Plants. 2024; 13(7):1023. https://doi.org/10.3390/plants13071023
Chicago/Turabian StyleMeena, Mintu Ram, K. Mohanraj, Ravinder Kumar, Raja Arun Kumar, Manohar Lal Chhabra, Neeraj Kulshreshtha, Gopalareddy Krishnappa, H. K. Mahadeva Swamy, A. Suganya, Perumal Govindaraj, and et al. 2024. "Deciphering Winter Sprouting Potential of Erianthus procerus Derived Sugarcane Hybrids under Subtropical Climates" Plants 13, no. 7: 1023. https://doi.org/10.3390/plants13071023
APA StyleMeena, M. R., Mohanraj, K., Kumar, R., Kumar, R. A., Chhabra, M. L., Kulshreshtha, N., Krishnappa, G., Swamy, H. K. M., Suganya, A., Govindaraj, P., & Hemaprabha, G. (2024). Deciphering Winter Sprouting Potential of Erianthus procerus Derived Sugarcane Hybrids under Subtropical Climates. Plants, 13(7), 1023. https://doi.org/10.3390/plants13071023