

Anthriscus sylvestris—Noxious Weed or Sustainable Source of Bioactive Lignans?


 ,
,  ,
, 
Abstract
:1. Introduction
1.1. Tradicional Medicine
1.2. Official Medicine
1.3. Nutritional Use
2. Composition
2.1. Root Composition
2.2. Aerial Parts Composition
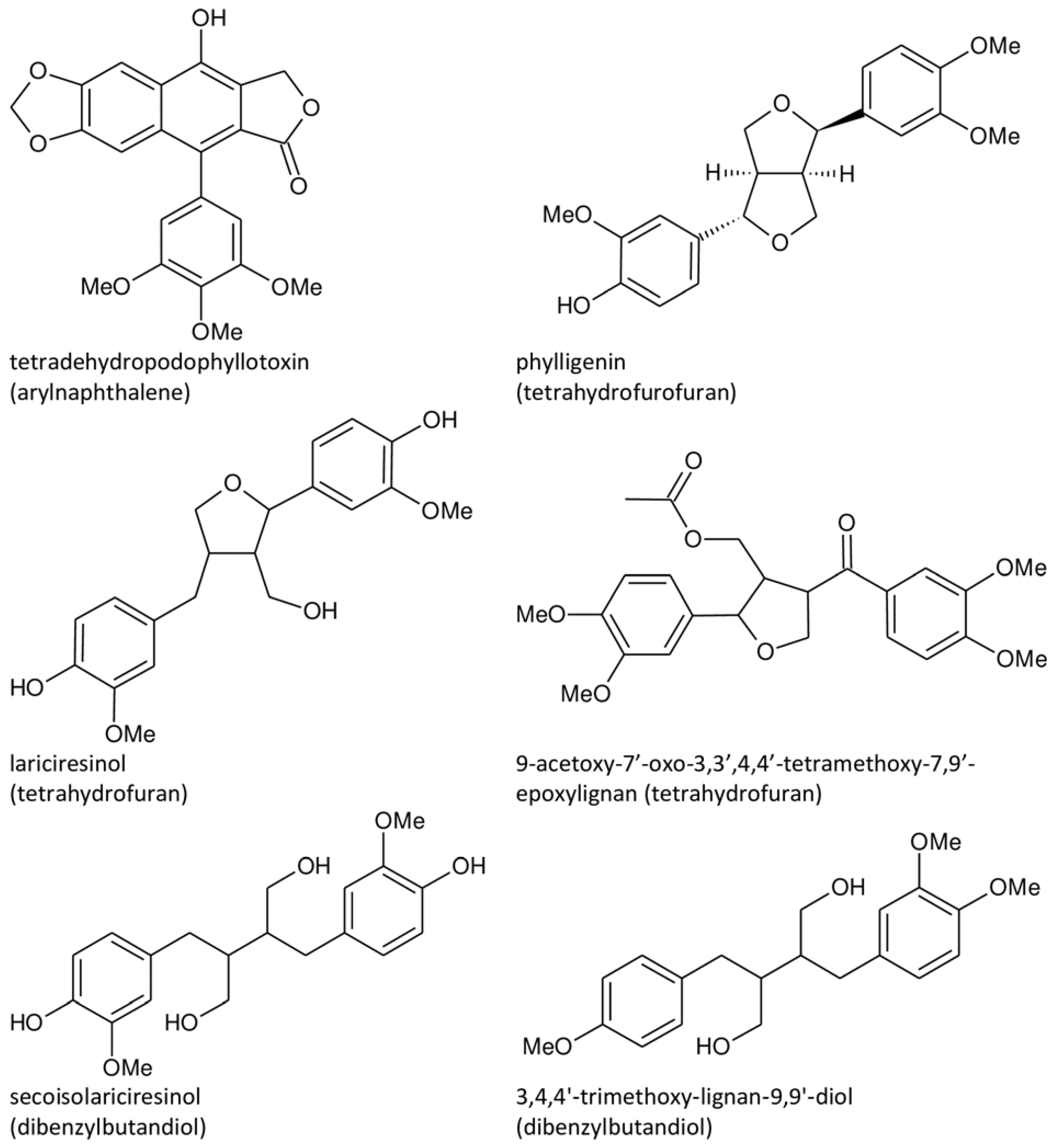
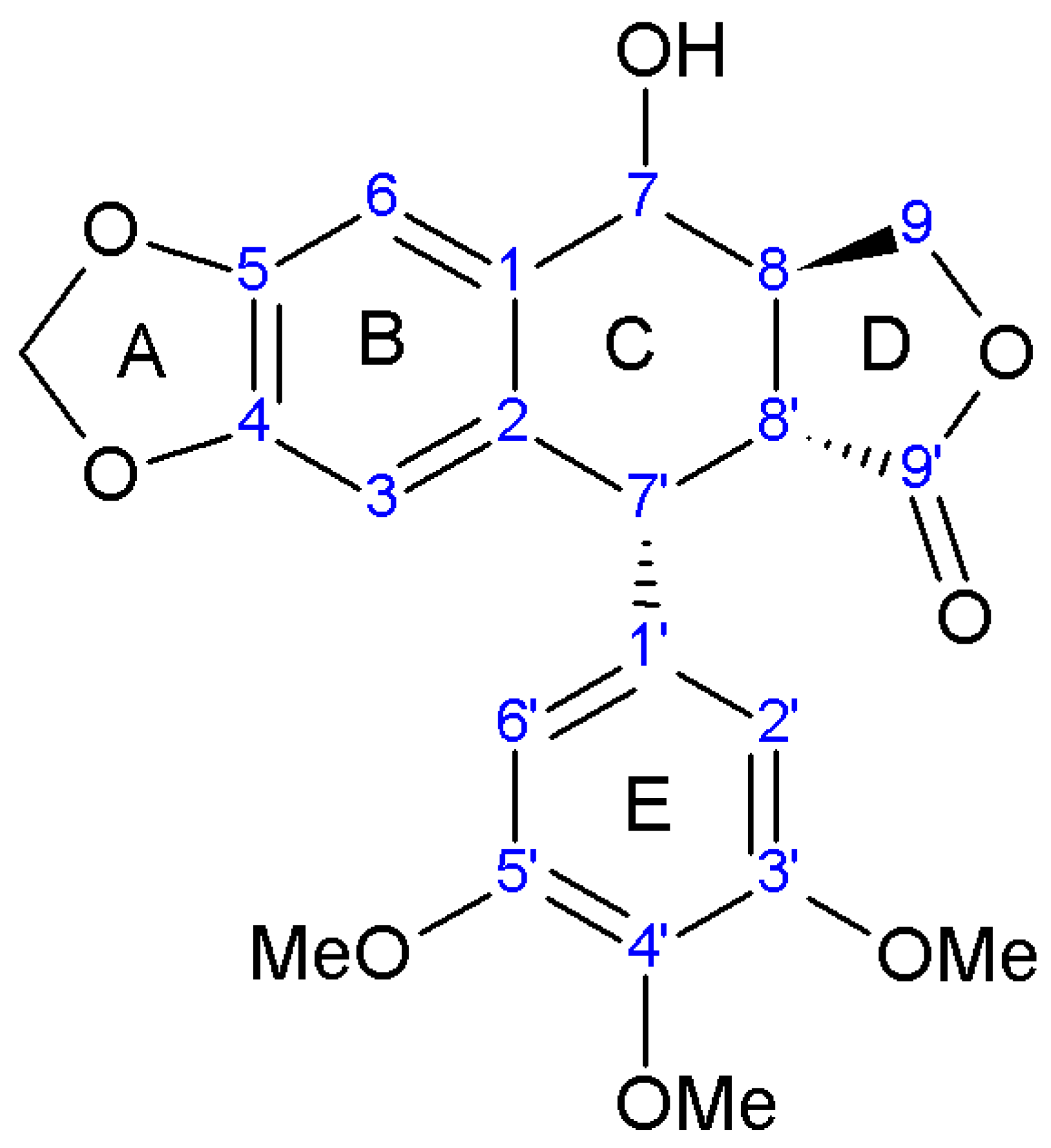
3. Lignan Structure
4. Biological Activity
4.1. Anti-Inflamatory Activity
4.1.1. Cyclooxigenase Inhibition
4.1.2. Antiasthmatic Activity
4.1.3. Cardiovascular Effects
4.1.4. Hyperpigmentation Treatment
4.1.5. Antiallergic Activity
4.2. Antiproliferative Activity
4.2.1. Antimitotic Activity
4.2.2. Topoisomerase II Inhibition
4.2.3. Caspase-3 Activity
4.2.4. Protein Kinase B (Akt) Inhibition
4.3. Antimicrobial Activity
4.3.1. Antiviral Activity
4.3.2. Antibacterial Activity
4.4. Antioxidant Activity
4.5. Other Activities
4.5.1. Cytochrome P450 (CYP) Inhibition
4.5.2. Neurotoxic Effect
4.5.3. Protective Activity
4.5.4. Immunomodulatory Activity
4.6. Toxicity towards Animals
5. Structure–Activity Relationship
5.1. Ring A Modifications
5.2. Ring B Modifications
5.3. Ring C Modifications
5.4. Ring D Modifications
5.5. Ring E Modifications
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| A. sylvestris | Anthriscus sylvestris |
| A-549 | human lung carcinoma cells |
| AGS | human gastric adenocarcinoma cells |
| AIF | apoptosis-inducing factor. |
| Akt | protein kinase B |
| AMPK | adenosine-monophosphate-activated protein kinase |
| APT | angeloyl podophyllotoxin |
| ATG | arctigenin |
| B16 | mouse melanoma cells |
| B16F10 | murine melanoma cell line |
| BGC-823 | human gastric adenocarcinoma cells |
| BMMC | bone-marrow-derived mast cells |
| CBS | colchicine-binding site |
| Cdk | cyclin-dependent kinase |
| Colo205 | colorectal adenocarcinoma cells |
| COX | cyclooxygenase |
| DDPT | 4′-O-demethyl-4-deoxypodophyllotoxin |
| DPP | deoxypicropodophyllin |
| DPT | deoxypodophyllotoxin |
| EGFR | epidermal growth factor receptor |
| ESCC | esophageal squamous cell carcinoma |
| ET | etoposide |
| H460 | nonsmall-cell lung cancer cells |
| HASMC | human aortic smooth muscle cells |
| HCC827GR | human lung adenocarcinoma cells |
| HeLa | immortalized cervical cancer cell line |
| Hepa 1c1c7 | mouse hepatoma cells |
| HepG2 | human liver cancer cell line |
| HIV | human immunodeficiency virus |
| HL-60 | human promyelocytic leukemia cells |
| HLM | human liver microsomes |
| HO-8910 | human ovarian carcinoma cells |
| HPV | human papilloma virus |
| HSV | herpes simplex viruses |
| HT-29 | human colorectal adenocarcinoma cells |
| HUVECs | human umbilical vein endothelial cells |
| IFV | influenza A virus |
| IgE | immunoglobulin E |
| IGF1R | insulin-like growth factor 1 receptor |
| IL | interleukin |
| ILC2s | type 2 innate lymphoid cells |
| iNOS | inducible nitric oxide synthase |
| IRF | interferon regulatory factor |
| JeG-3 | human choriocarcinoma cells |
| K562 | human leukemia cells |
| L5178Y | mouse leukemic cells |
| LOX | lipoxygenase |
| LPS | lipopolysaccharide |
| LT | leukotriene |
| MAPKs | mitogen-activated protein kinases |
| MAT | matairesinol |
| MCF-7 | human breast adenocarcinoma cells |
| MCMV | murine cytomegalovirus |
| MDA-MB-231 | human breast adenocarcinoma cells |
| MDR | multidrug resistance |
| MEL-28 | human melanoma cells |
| MET | Mesenchymal–epithelial transition |
| MG-63 | human osteosarcoma cells |
| MH60 | murine hepatoma cells |
| MK-1 | human epithelial cell line |
| MMP | matrix metalloproteinase |
| NF-κB | nuclear factor kappa B |
| NSAID | nonsteroidal anti-inflammatory drugs |
| NSCLC | nonsmall-cell lung cancer |
| OA | osteoarthritis |
| OVA | ovalbumin |
| P-388 | mouse leukemic cells |
| PANC-1 | pancreatic cancer cell line |
| PARP-1 | poly(ADP-ribose) polymerase 1. |
| PCA | passive cutaneous anaphylaxis |
| PG | prostaglandin |
| PI3K | phosphoinositide 3-kinase |
| PLK | polo-like kinase |
| PPT | picropodophyllotoxin |
| PT | podophyllotoxin |
| RAW264.7 cells | murine macrophage cell line |
| ROS | reactive oxygen species |
| SF126 | human glioblastoma cell line |
| SGC-7901 | human gastric adenocarcinoma cells |
| SiHa | human cervical carcinoma cells |
| SK-MEL-2 | human melanoma cells |
| SK-OV-3 | human ovarian adenocarcinoma cells |
| Th2 | T helper cell type 2 |
| TKIs | tyrosine kinase inhibitors |
| TNF-α | tumor necrosis factor alpha |
| U-87 MG | human glioblastoma cells |
| YAT | yatein |
References
- Plunkett, G.M.; Soltis, D.E.; Soltis, P.S. Evolutionary Patterns in Apiaceae: Inferences Based on MatK Sequence Data. Syst. Bot. 1996, 21, 477–495. [Google Scholar] [CrossRef]
- Hultén, E.; Fries, M. Atlas of North European Vascular Plants North of the Tropic of Cancer; Koeltz Scientific Books: Konigstein, Germany, 1984. [Google Scholar]
- Walton, D.W.H. European Weeds and Other Alien Species in the Subantarctic. Weed Res. 1975, 15, 271–282. [Google Scholar] [CrossRef]
- Olaru, O.T.; Niţulescu, G.M.; Orţan, A.; Dinu-Pîrvu, C.E. Ethnomedicinal, Phytochemical and Pharmacological Profile of Anthriscus sylvestris as an Alternative Source for Anticancer Lignans. Molecules 2015, 20, 15003–15022. [Google Scholar] [CrossRef] [PubMed]
- Townsend, C.C. One New and One Disjunct Variety of Umbelliferae from East Africa. Kew Bull. 1984, 39, 603–605. [Google Scholar] [CrossRef]
- Webb, C.J.; Sykes, W.R.; Garnock-Jones, P.J.; Given, D.R.; Brownsey, P.J. Checklist of Dicotyledons, Gymnosperms, and Pteridophytes Naturalised in New Zealand: Additional Records and Corrections. N. Z. J. Bot. 1989, 27, 139–162. [Google Scholar] [CrossRef]
- Hiroe, M. Umbelliferae of World; Ariake Book Company: Tokyo, Japan, 1979. [Google Scholar]
- Coulter, J.M.; Rose, J.N. Monograph of the North American Umbelliferae; Biodiversity Heritage Library: Washington, DC, USA, 1900; Volume 7, pp. 9–256. [Google Scholar]
- Heywood, V.H. Systematic Survey of Old World Umbelliferae. In The Biology and Chemistry of the Umbelliferae; Heywood, V.H., Ed.; Academic Press: London, UK, 1971; pp. 31–41. [Google Scholar]
- Tutin, T.G. Umbellifers of the British Isles. BSBI Handbook No. 2.; Botanical Society of Britain & Ireland: London, UK, 1980; ISBN 9780901158024. [Google Scholar]
- Spalik, K.; Downie, S.R. The Utility of Morphological Characters for Inferring Phylogeny in Scandiceae Subtribe Scandicinae (Apiaceae). Ann. Missouri Bot. Gard. 2001, 88, 270–301. [Google Scholar] [CrossRef]
- Tekin, M.; Civelek, S. A Taxonomic Revision of the Genus Anthriscus (Apiaceae) in Turkey. Phytotaxa 2017, 302, 001–026. [Google Scholar] [CrossRef]
- POWO Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 23 November 2023).
- Cannon, J.F.M. Anthriscus. In Flora Europaea: Volume 2 Rosaceae to Umbelliferae; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; p. 326. [Google Scholar]
- Hruška, K. Considerazioni Ecologiche, Fitosociologiche e Morfologiche Sul Genere Anthriscus pers. G. Bot. Ital. 1982, 116, 175–187. [Google Scholar] [CrossRef]
- Hedge, I.C.; Lamond, J.M. Umbelliferae. In Flora of Turkey and the East Aegean Islands; Davis, P.H., Chamberlain, D.F., Matthews, V.A., Eds.; Edinburgh University Press: Edinburgh, UK, 1972. [Google Scholar]
- Hand, R. Euro+Med PlantBase. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 23 November 2023).
- Muckensturm, B.; Diyani, F.; Reduron, J.-P. Grilactone and Other Terpenoids from Anthriscus nitida. Biochem. Syst. Ecol. 1995, 23, 875–876. [Google Scholar] [CrossRef]
- Deforce, K. The Historical Use of Ladanum. Palynological Evidence from 15th and 16th Century Cesspits in Northern Belgium. Veg. Hist. Archaeobot. 2006, 15, 145–148. [Google Scholar] [CrossRef]
- Hadač, E. Ruderal Vegetation of the Broumov Basin, NE. Bohemia. Folia Geobot. Phytotaxon. 1978, 13, 129–163. [Google Scholar] [CrossRef]
- Derbyshire, S.J.; Hoeg, R.; Haverkort, J. The Biology of Canadian Weeds. 111. Anthriscus sylvestris (L.) Hoffm. Can. J. Plant Sci. 1999, 79, 671–682. [Google Scholar] [CrossRef]
- Hansson, M.L.; Persson, T.S. Anthriscus sylvestris—A Growing Conservation Problem? Ann. Bot. Fenn. 1994, 31, 205–213. [Google Scholar]
- Spalik, K.; Woodel, S.R.J. Regulation of Pollen Production in Anthriscus sylvestris, an Andromonoecious Species. Int. J. Plant Sci. 1994, 155, 750–754. [Google Scholar] [CrossRef]
- Tamamschian, S. Materials for the Karyosystematics of the Cultivated and Wild Growing Species of the Family Umbelliferae. Tr. Prikl. Bot. Genet. Sel. Ser. 2 1933, 2, 137–164. [Google Scholar]
- Chatterjee, A.; Ghosh, S.; Roy, S.C. A Cytological Survey of Eastern Himalayan Plants III. Cell Chromosom. Res. 1989, 12, 22–29. [Google Scholar]
- Koulman, A.; Bos, R.; Medarde, M.; Pras, N.; Quax, W.J. A Fast and Simple GC MS Method for Lignan Profiling in Anthriscus sylvestris and Biosynthetically Related Plant Species. Planta Med. 2001, 67, 858–862. [Google Scholar] [CrossRef] [PubMed]
- Loike, J.D.; Brewer, C.F.; Sternlicht, H.; Gensler, W.J.; Horwitz, S.B. Structure-Activity Study of the Inhibition of Microtubule Assembly in vitro by Podophyllotoxin and Its Congeners. Cancer Res. 1978, 38, 2688–2693. [Google Scholar] [PubMed]
- Canel, C.; Moraes, R.M.; Dayan, F.E.; Ferreira, D. Podophyllotoxin. Phytochemistry 2000, 54, 115–120. [Google Scholar] [CrossRef]
- Koulman, A.; Kubbinga, M.E.; Batterman, S.; Woerdenbag, H.J.; Pras, N.; Woolley, J.G.; Quax, W.J. A Phytochemical Study of Lignans in Whole Plants and Cell Suspension Cultures of Anthriscus sylvestris. Planta Med. 2003, 69, 733–738. [Google Scholar] [CrossRef]
- Rios, J.L.; Giner, R.M.; Prieto, J.M. New Findings on the Bioactivity of Lignans. Stud. Nat. Prod. Chem. 2002, 26, 183–292. [Google Scholar] [CrossRef]
- Slanina, J.; Glatz, Z. Separation Procedures Applicable to Lignan Analysis. J. Chromatogr. B 2004, 812, 215–229. [Google Scholar] [CrossRef]
- Ingram, D.; Sanders, K.; Kolybaba, M.; Lopez, D. Case-Control Study of Phyto-Oestrogens and Breast Cancer. Lancet 1997, 350, 990–994. [Google Scholar] [CrossRef] [PubMed]
- Pietinen, P.; Stumpf, K.; Männistö, S.; Kataja, V.; Uusitupa, M.; Adlercreutz, H. Serum Enterolactone and Risk of Breast Cancer: A Case-Control Study in Eastern Finland. Cancer Epidemiol. Biomarkers Prev. 2001, 10, 339–344. [Google Scholar] [PubMed]
- Kilkkinen, A.; Virtamo, J.; Vartiainen, E.; Sankila, R.; Virtanen, M.J.; Adlercreutz, H.; Pietinen, P. Serum Enterolactone Concentration Is Not Associated with Breast Cancer Risk in a Nested Case-Control Study. Int. J. Cancer 2004, 108, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Vanharanta, M.; Voutilainen, S.; Rissanen, T.H.; Adlercreutz, H.; Salonen, J.T. Risk of Cardiovascular Disease-Related and All-Cause Death According to Serum Concentrations of Enterolactone: Kuopio Ischaemic Heart Disease Risk Factor Study. Arch. Intern. Med. 2003, 163, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Podwyssotzki, V. Pharmakologische Studien Über Podophyllum Peltatum. Arch. Exp. Pathol. Pharmakol. 1880, 13, 29–52. [Google Scholar] [CrossRef]
- Noguchi, T.; Kawanami, M. Studies on the Constituents of Anthriscus sylvestris Hoffm. Yakugaku Zasshi 1940, 60, 629–636. [Google Scholar]
- Sackett, D.L. Podophyllotoxin, Steganacin and Combretastatin: Natural Products That Bind at the Colchicine Site of Tubulin. Pharmacol. Ther. 1993, 59, 163–228. [Google Scholar] [CrossRef]
- Hartwell, J.L.; Schrecker, A.W. The Chemistry of Podophyllum. Fortschritte Chem. Org. Naturstoffe 1958, 15, 83–166. [Google Scholar] [CrossRef]
- Kozawa, M.; Baba, K.; Matsuyama, Y.; Kido, T.; Sakai, M.; Takemoto, T. Components of the Root of Anthriscus sylvestris Hoffm. II. Insecticidal Activity. Chem. Pharm. Bull. 1982, 30, 2885–2888. [Google Scholar] [CrossRef]
- Lee, S.H.; Son, M.J.; Ju, H.K.; Lin, C.X.; Moon, T.C.; Choi, H.; Son, J.K.; Chang, H.W. Dual Inhibition of Cyclooxygenases-2 and 5-Lipoxygenase by Deoxypodophyllotoxin in Mouse Bone Marrow-Derived Mast Cells. Biol. Pharm. Bull. 2004, 27, 786–788. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.; Kwon, O.; Park, B.; Oh, S.; Ahn, K.; Chang, M.; Oh, W.K.; Kim, J.; Min, B.; Kim, Y.; et al. Lignans and Coumarins from the Roots of Anthriscus sylvestris and Their Increase of Caspase-3 Activity in HL-60 Cells. Biol. Pharm. Bull. 2007, 30, 1340–1343. [Google Scholar] [CrossRef] [PubMed]
- Milovanovic, M.; Banjac, N.; Vucelic-Radovic, B. Functional Food: Rare Herbs, Seeds and Vegetable Oils as Sources of Flavors and Phytosterols. J. Agric. Sci. Belgrade 2009, 54, 80–93. [Google Scholar] [CrossRef]
- Allen, D.E.; Hatfield, G. Medicinal Plants in Folk Tradition: An Ethnobotany of Britain and Ireland; Timber Press, Inc.: Kingston, WA, USA, 2004; ISBN 0881926388. [Google Scholar]
- Wahida, B.; Amor, M.; Nabil, C. An Inventory of Ethnomedicinal Plants Used in Tunisia. In Ethnomedicinal Plants: Revitalization of Traditional Knowledge of Herbs; Rai, M., Charya, D., Luis Rios, J., Eds.; Taylor and Francis Group, LLC: Abingdon, UK, 2011; pp. 333–360. ISBN 9781439853627. [Google Scholar]
- Gross, G. Clinical Aspects and Therapy of Anogenital Warts and Papillomavirus-Associated Lesions. Hautarzt 2001, 52, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Carlström, K.; Hedin, P.; Jönsson, L.; Lerndal, T.; Lien, J.; Weitoft, T.; Axelson, M. Endocrine Effects of the Podophyllotoxine Derivative Drug CPH 82 (Reumacon®) in Patients with Rheumatoid Arthritis. Scand. J. Rheumatol. 2000, 29, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Özqeltk, H. Notes on Economic Plants. Econ. Bot. 1994, 48, 214–221. [Google Scholar] [CrossRef]
- Orčić, D.; Berežni, S.; Mimica-Dukić, N. Quantitative HPLC-UV Study of Lignans in Anthriscus sylvestris. Molecules 2022, 27, 6072. [Google Scholar] [CrossRef] [PubMed]
- Kozawa, M.; Morita, N.; Hata, K. Chemical Components of the Roots of Anthriscus sylvestris Hoffm. I. Structures of an Acyloxycarboxylic Acid and a New Phenylpropanoidester, Anthriscusin. Yakugaku Zasshi 1978, 98, 1486–1490. [Google Scholar] [CrossRef]
- Ikeda, R.; Nagao, T.; Okabe, H.; Nakano, Y.; Matsunaga, H.; Katano, M.; Mori, M. Antiproliferative Constituents in Umbelliferae Plants. III. Constituents in Teh Root and the Ground Part of Anthriscus sylvestris Hoffm. Chem. Pharm. Bull. 1998, 46, 871–874. [Google Scholar] [CrossRef]
- Lim, Y.; Leem, M.; Shin, D.; Chang, H.; Hong, S.; Moon, E.; Lee, D.; Yoon, S.; Woo, W. Cytotoxic Constituents from the Roots of Anthriscus sylvestris. Arch. Pharm. Res. 1999, 22, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Sakakibara, N.; Umezawa, T.; Shimada, M. Survey and Enzymatic Formation of Lignans of Anthriscus sylvestris. J. Wood Sci. 2002, 48, 536–541. [Google Scholar] [CrossRef]
- Hendrawati, O.; Woerdenbag, H.J.; Michiels, P.J.A.; Aantjes, H.G.; van Dam, A.; Kayser, O. Identification of Lignans and Related Compounds in Anthriscus sylvestris by LC-ESI-MS/MS and LC-SPE-NMR. Phytochemistry 2011, 72, 2172–2179. [Google Scholar] [CrossRef]
- Seegers, C.L.C.; Tepper, P.G.; Setroikromo, R.; Quax, W.J. Cytotoxic Deoxypodophyllotoxin Can Be Extracted in High Purity from Anthriscus sylvestris Roots by Supercritical Carbon Dioxide. Planta Med. 2018, 84, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Kozawa, M.; Morita, N.; Hata, K. Structure of Anthriscusin, a New Phenylpropanoid Ester from the Roots of Anthriscus sylvestris Hoffm. Chem. Pharm. Bull. 1978, 26, 1337–1338. [Google Scholar] [CrossRef]
- Kurihara, T.; Kikuchi, M.; Suzuki, S.; Hisamichi, S. Studies on the Constituents of Anthriscus sylvestris Hoffm. I. On the Components of the Radix. Yakugaku Zasshi 1978, 98, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Lei, B.; Ning, N.; Fan, J.; Zhang, X.; Ma, C.; Jiang, H. A New Phenylpropanoid Ester from the Roots of Anthriscus sylvestris and Its Chemotaxonomic Significance. Biochem. Syst. Ecol. 2020, 93, 104144. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, Y.; Niu, Y.; Zheng, Y.; Chen, X.; Ren, Y.; Fan, X.; Li, X.; Ma, X.; Zheng, X.; et al. Diarylpentanoids and Phenylpropanoids from the Roots of Anthriscus sylvestris (L.) Hoffm. Phytochemistry 2023, 216, 113865. [Google Scholar] [CrossRef] [PubMed]
- Bos, R.; Koulman, A.; Woerdenbag, H.J.; Quax, W.J.; Pras, N. Volatile Components from Anthriscus sylvestris (L.) Hoffm. J. Chromatogr. A 2002, 966, 233–238. [Google Scholar] [CrossRef]
- Kramer, M.; Mühleis, A.; Conrad, J.; Leitenberger, M.; Beifuss, U.; Carle, R.; Kammerer, D.R. Quantification of Polyacetylenes in Apiaceous Plants by High-Performance Liquid Chromatography Coupled with Diode Array Detection. Z. Naturforsch 2011, 66, 319–327. [Google Scholar] [CrossRef]
- Dall’Acqua, S.; Giorgetti, M.; Cervellati, R.I.; Nnocenti, G. Deoxypodophyllotoxin Content and Antioxidant Activity of Aerial Parts of Anthriscus sylvestris Hoffm. Z. Naturforsch 2006, 61, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Milovanovic, M.; Stefanovic, M.; Djermanovic, V.; Milovanovic, J. Some Chemical Constituents of Anthriscus sylvestris. J. Herbs. Spices Med. Plants 1996, 4, 17–22. [Google Scholar] [CrossRef]
- Milovanovic, M.; Picuric-Jovanovic, K.; Vucelic-Radovic, B.; Vrbaski, Z. Antioxidant Effects of Flavonoids of Anthriscus sylvestris in Lard. J. Am. Oil Chem. Soc. 1996, 73, 773–776. [Google Scholar] [CrossRef]
- Ikeda, R.; Nagao, T.; Okabe, H.; Nakano, Y.; Matsunaga, H.; Katano, M.; Mori, M. Antiproliferative Constituents in Umbelliferae Plants. IV. Constituents in the Fruits of Anthriscus sylvestris Hoffm. Chem. Pharm. Bull. 1998, 46, 875–878. [Google Scholar] [CrossRef] [PubMed]
- Janković, M.; Berežni, S.; Orčić, D. Quantitative Analysis of Lignans from the Fruits of Wild Chervil (Anthriscus sylvestris (L.)Hoffm.). FACTA Univ. Ser. Physics, Chem. Technol. 2023, 21, 39–46. [Google Scholar] [CrossRef]
- Janković, M.; Berežni, S.; Orčić, D. Lignan Profile in Fruits of Wild Chervil (Anthriscus sylvestris (L.) Hoffm.). FACTA Univ. Ser. Physics, Chem. Technol. 2023, 21, 27–37. [Google Scholar] [CrossRef]
- Kurihara, T.; Kikuchi, M. Studies on the Constituents of Anthriscus sylvestris Hoffm. II. On the Components of the Flowers and Leaves. Yakugaku Zasshi 1979, 99, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Borg-Karlson, A.; Valterová, I.; Anders Nilsson, L. Volatile Compounds from Flowers of Six Species in the Family Apiaceae: Bouquets for Different Pollinators? Phytochemistry 1994, 35, 111–119. [Google Scholar] [CrossRef]
- Sólyomváry, A.; Béni, S.; Boldizsár, I. Dibenzylbutyrolactone Lignans—A Review of Their Structural Diversity, Biosynthesis, Occurrence, Identification and Importance. Mini-Rev. Med. Chem. 2017, 17, 1053–1074. [Google Scholar] [CrossRef]
- Guerram, M.; Jiang, Z.Z.; Zhang, L.Y. Podophyllotoxin, a Medicinal Agent of Plant Origin: Past, Present and Future. Chin. J. Nat. Med. 2012, 10, 161–169. [Google Scholar] [CrossRef]
- Umezawa, T. Diversity in Lignan Biosynthesis. Phytochem. Rev. 2003, 2, 371–390. [Google Scholar] [CrossRef]
- Orčić, D.; Berežni, S.; Škorić, D.; Mimica-Dukić, N. Comprehensive Study of Anthriscus sylvestris Lignans. Phytochemistry 2021, 192, 112958. [Google Scholar] [CrossRef]
- Ayres, D.C.; Loike, J.D. Lignans Chemical, Biological and Clinical Properties; Cambridge University Press: Cambridge, UK, 1990; ISBN 9780511983665. [Google Scholar]
- Dewick, P.M.; Jackson, D.E. Cytotoxic Lignans from Podophyllum, and the Nomenclature of Aryltetralin Lignans. Phytochemistry 1981, 20, 2277–2280. [Google Scholar] [CrossRef]
- Cui, Q.; Du, R.; Liu, M.; Rong, L. Lignans and Their Derivatives from Plants as Antivirals. Molecules 2020, 25, 183. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, N.; Suzuki, S.; Umezawa, T.; Shimada, M. Biosynthesis of Yatein in Anthriscus sylvestris. Org. Biomol. Chem. 2003, 1, 2474–2485. [Google Scholar] [CrossRef] [PubMed]
- Koulman, A. Podophyllotoxin: A Study of the Biosynthesis, Evolution, Function and Use of Podophyllotoxin and Related Lignans; University of Groningen: Groningen, The Netherlands, 2003; Chapter X. [Google Scholar]
- Orčić, D. Vrste Tribusa Scandiceae (Apiaceae Lindley 1836, Subfam. Apioideae) Potencijalni Izvor Biološki i Farmakološki Aktivnih Sekundarnih Biomolekula; University of Novi Sad: Novi Sad, Serbia, 2010. [Google Scholar]
- Ragamustari, S.K.; Nakatsubo, T.; Hattori, T.; Ono, E.; Kitamura, Y.; Suzuki, S.; Yamamura, M.; Umezawa, T. A Novel O-Methyltransferase Involved in the First Methylation Step of Yatein Biosynthesis from Matairesinol in Anthriscus sylvestris. Plant Biotechnol. 2013, 30, 375–384. [Google Scholar] [CrossRef]
- Kamil, W.M.; Dewick, P.M. Biosynthetic Relationship of Aryltetralin Lactone Lignans to Dibenzylbutyrolactone Lignans. Phytochemistry 1986, 25, 2093–2102. [Google Scholar] [CrossRef]
- Jackson, D.E.; Dewick, P.M. Biosynthesis of Podophyllum Lignans—II. Interconversions of Aryltetralin Lignans in Podophyllum Hexandrum. Phytochemistry 1984, 23, 1037–1042. [Google Scholar] [CrossRef]
- Lin, C.X.; Son, M.J.; Ju, H.K.; Moon, T.C.; Lee, E.; Kim, S.H.; Kim, M.; Son, J.K.; Lee, S.H.; Chang, H.W. Deoxypodophyllotoxin, a Naturally Occurring Lignan, Inhibits the Passive Cutaneous Anaphylaxis Reaction. Planta Med. 2004, 70, 474–476. [Google Scholar] [CrossRef]
- Lin, C.X.; Lee, E.; Jin, M.H.; Yook, J.; Quan, Z.; Ha, K.; Moon, T.C.; Kim, M.J.; Kim, K.J.; Lee, S.H.; et al. Deoxypodophyllotoxin (DPT) Inhibits Eosinophil Recruitment into the Airway and Th2 Cytokine Expression in an OVA-Induced Lung Inflammation. Planta Med. 2006, 72, 786–791. [Google Scholar] [CrossRef]
- Jin, M.; Moon, T.C.; Quan, Z.; Lee, E.; Kim, Y.K.; Yang, J.H.; Suh, S.J.; Jeong, T.C.; Lee, S.H.; Kim, C.H.; et al. The Naturally Occurring Flavolignan, Deoxypodophyllotoxin, Inhibits Lipopolysaccharide-Induced INOS Expression through the NF-κB Activation in RAW264.7 Macrophage Cells. Biol. Pharm. Bull. 2008, 31, 1312–1315. [Google Scholar] [CrossRef] [PubMed]
- Yong, Y.; Shin, S.Y.; Lee, Y.H.; Lim, Y. Antitumor Activity of Deoxypodophyllotoxin Isolated from Anthriscus sylvestris: Induction of G2/M Cell Cycle Arrest and Caspase-Dependent Apoptosis. Bioorg. Med. Chem. Lett. 2009, 19, 4367–4371. [Google Scholar] [CrossRef] [PubMed]
- Quan, G.H.; Chin, Y.W.; Lee, H.K.; Oh, S.R. Preparative Isolation and Purification of Deoxypodophyllotoxin from the Rhizomes of Anthriscus sylvestris by High-Speed Counter-Current Chromatography. J. Korean Soc. Appl. Biol. Chem. 2010, 53, 110–113. [Google Scholar] [CrossRef]
- Jung, C.H.; Kim, H.; Ahn, J.; Jung, S.K.; Um, M.Y.; Son, K.H.; Kim, T.W.; Ha, T.Y. Anthricin Isolated from Anthriscus sylvestris (L.) Hoffm. Inhibits the Growth of Breast Cancer Cells by Inhibiting Akt/mTOR Signaling, and Its Apoptotic Effects Are Enhanced by Autophagy Inhibition. Evid.-Based Complement. Altern. Med. 2013, 2013, 385219. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.J.; Choi, J.M.; Kim, H.M.; Choi, K.; Ku, J.; Park, K.W.; Kim, J.; Lee, S. Antibacterial Activity and Protective Effect against Gastric Cancer by Anthriscus sylvestris Fractions. Hortic. Environ. Biotechnol. 2013, 54, 326–330. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, H.Z.; Li, Y.C.; Wei, G.Q.; Geng, Y.; Ma, C.Y. Antitumor Constituents from Anthriscus sylvestris (L.) Hoffm. Asian Pacific J. Cancer Prev. 2014, 15, 2803–2807. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lee, A.Y.; Moon, B.C.; Kim, W.J.; Choi, G. Ultrasonic-Assisted Extraction Process and Method Validation for Deoxypodophyllotoxin from the Roots of Anthriscus sylvestris: Application of Response Surface Methodology and UPLC–PDA–QDa. Acta Chromatogr. 2019, 31, 126–132. [Google Scholar] [CrossRef]
- Velescu, B.Ş.; Anuţa, V.; Nițulescu, G.M.; Olaru, O.T.; Orțan, A.; Ionescu, D.; Ghica, M.V.; Drăgoi, C.M.; Pîrvu, C.E.D. Pharmaceutical Assesment of Romanian Crops of Anthriscus sylvestris (Apiaceae). Farmacia 2017, 65, 824–831. [Google Scholar]
- Lee, S.A.; Moon, S.; Han, S.H.; Hwang, E.J.; Park, B.; Kim, J.; Kim, D.K.; Kim, C.S. Chondroprotective Effects of Aqueous Extract of Anthriscus sylvestris Leaves on Osteoarthritis in vitro and in vivo through MAPKs and NF-κB Signaling Inhibition. Biomed. Pharmacother. 2018, 103, 1202–1211. [Google Scholar] [CrossRef]
- Lee, S.A.; Moon, S.M.; Han, S.H.; Hwang, E.J.; Hong, J.H.; Park, B.R.; Choi, M.S.; Ahn, H.; Kim, J.S.; Kim, H.J.; et al. In vivo and in vitro Anti-Inflammatory Effects of Aqueous Extract of Anthriscus sylvestris Leaves. J. Med. Food 2018, 21, 585–595. [Google Scholar] [CrossRef]
- Vane, J.R. Inhibition of Prostaglandin Synthesis as a Mechanism of Action for Aspirin-like Drugs. Nat. New Biol. 1971, 231, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Whittle, B.J.R. Arachidonic Acid Metabolites and the Gastro-Intestinal Toxicity of Anti-Inflammatory Agents. Prostaglandins 1981, 21, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Schneider, I.; Bucar, F. Lipoxygenase Inhibitors from Natural Plant Sources. Part 2: Medicinal Plants with Inhibitory Activity on Arachidonate 12-Lipoxygenase, 15-Lipoxygenase and Leukotriene Receptor Antagonists. Phyther. Res. 2005, 19, 263–272. [Google Scholar] [CrossRef] [PubMed]
- León, B.; Ballesteros-Tato, A. Modulating Th2 Cell Immunity for the Treatment of Asthma. Front. Immunol. 2021, 12, 637948. [Google Scholar] [CrossRef] [PubMed]
- Drazen, J.M.; Arm, J.P.; Austen, K.F. Sorting out the Cytokines of Asthma. J. Exp. Med. 1996, 183, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Chanez, P.; Lacoste, J.Y.; Barneon, G.; Ghavanian, N.; Enander, I.; Venge, P.; Ahlstedt, S.; Simony-Lafontaine, J.; Godard, P.; et al. Eosinophilic Inflammation in Asthma. N. Engl. J. Med. 1990, 323, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Maarsingh, H.; Dekkers, B.G.J.; Zuidhof, A.B.; Bos, I.S.T.; Menzen, M.H.; Klein, T.; Flik, G.; Zaagsma, J.; Meurs, H. Increased Arginase Activity Contributes to Airway Remodelling in Chronic Allergic Asthma. Eur. Respir. J. 2011, 38, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Hamad, A.M.; Knox, A.J. Mechanisms Mediating the Antiproliferative Effects of Nitric Oxide in Cultured Human Airway Smooth Muscle Cells. FEBS Lett. 2001, 506, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of Nitric Oxide in Inflammatory Diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Kim, S.B.; Lee, A.Y.; Chun, J.M.; Lee, A.R.; Kim, H.S.; Seo, Y.S.; Moon, B.C.; Kwon, B.I. Anthriscus sylvestris Root Extract Reduces Allergic Lung Inflammation by Regulating Interferon Regulatory Factor 4-Mediated Th2 Cell Activation. J. Ethnopharmacol. 2019, 25, 165–175. [Google Scholar] [CrossRef]
- An, M.; Oh, M.; Park, K.T.; Seon, K.H.; Jo, J.E.; Lee, S.K.; Kim, J.K.; Shin, K.S.; Koh, J.H.; Lim, Y.H. Anti-Asthma and Antitussive Effects of a Fermented Extract of a Mixture of Ramulus Mori, Anthriscus sylvestris, and Salvia plebeian. Food Sci. Biotechnol. 2021, 30, 1257–1268. [Google Scholar] [CrossRef]
- Suh, S.J.; Kim, J.R.; Jin, U.H.; Choi, H.S.; Chang, Y.C.; Lee, Y.C.; Kim, S.H.; Lee, I.S.; Moon, T.C.; Chang, H.W.; et al. Deoxypodophyllotoxin, Flavolignan, from Anthriscus sylvestris Hoffm. Inhibits Migration and MMP-9 via MAPK Pathways in TNF-α-Induced HASMC. Vascul. Pharmacol. 2009, 51, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Farina, A.R.; Mackay, A.R. Gelatinase B/MMP-9 in Tumour Pathogenesis and Progression. Cancers 2014, 6, 240–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, B.; Guerram, M.; Sun, L.; Shi, W.; Tian, C.; Zhu, X.; Jiang, Z.; Zhang, L. Deoxypodophyllotoxin Suppresses Tumor Vasculature in HUVECs by Promoting Cytoskeleton Remodeling through LKB1-AMPK Dependent Rho A Activation. Oncotarget 2015, 6, 29497–29512. [Google Scholar] [CrossRef]
- Choi, H.; Lee, J.; Shin, H.; Lee, B.; Chang, I.; Hwang, J. Deoxypodophyllotoxin Reduces Skin Pigmentation of Brown Guinea Pigs. Planta Med. 2004, 70, 378–380. [Google Scholar] [CrossRef] [PubMed]
- Briganti, S.; Camera, E.; Picardo, M. Chemical and Instrumental Approaches to Treat Hyperpigmentation. Pigment Cell Res. 2003, 16, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Muto, N.; Tomokuni, T.; Haramoto, M.; Tatemoto, H.; Nakanishi, T.; Inatomi, Y.; Murata, H.; Inada, A. Isolation of Apoptosis- and Differentiation-Inducing Substances toward Human Promyelocytic Leukemia HL-60 Cells from Leaves of Juniperus taxifolia. Biosci. Biotechnol. Biochem. 2008, 72, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; You, Y.J.; Nam, N.H.; Ahn, B.Z. 2,3-Dibenzylbutyrolactones and 1,2,3,4-Tetrahydro-2-Naphthoic Acid y-Lactones: Structure and Activity Relationship in Cytotoxic Activity. Arch. Pharm. Res. 2002, 25, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.; Singh Negi, A.S.; Kumar, J.K.; Gupta, M.M.; Khanuja, S.P.S. Plant-Based Anticancer Molecules: A Chemical and Biological Profile of Some Important Leads. Bioorg. Med. Chem. 2005, 13, 5892–5908. [Google Scholar] [CrossRef]
- Inamori, Y.; Kubo, M.; Tsujibo, H.; Ogawa, M.; Baba, K.; Kozawa, M.; Fujita, E. The Biological Activities of Podophyllotoxin Compounds. Chem. Pharm. Bull. 1986, 34, 3928–3932. [Google Scholar] [CrossRef]
- Levy, R.K.; Hall, I.H.; Lee, K.H. Antitumor Agents LXII: Synthesis and Biological Evaluation of Podophyllotoxin Esters and Related Derivatives. J. Pharm. Sci. 1983, 72, 1158–1161. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Lee, H.J.; Kim, C.Y.; Lee, S.B.; Tunsag, J.; Batsuren, D.; Nho, C.W. The Chemopreventive Effects of Saussurea salicifolia through Induction of Apoptosis and Phase II Detoxification Enzyme. Biol. Pharm. Bull. 2007, 30, 2352–2359. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Gotoh, M.; Oka, K. Natural Flavonoids and Lignans Are Potent Cytostatic Agents against Human Leukemic HL-60 Cells. Life Sci. 1994, 55, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Hosono-Nishiyama, K.; Yamada, H. Antiproliferative and Apoptotic Effects of Butyrolactone Lignans from Arctium Lappa on Leukemic Cells. Planta Med. 2006, 72, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Awale, S.; Lu, J.; Kalauni, S.K.; Kurashima, Y.; Tezuka, Y.; Kadota, S.; Esumi, H. Identification of Arctigenin as an Antitumor Agent Having the Ability to Eliminate the Tolerance of Cancer Cells to Nutrient Starvation. Cancer Res. 2006, 66, 1751–1757. [Google Scholar] [CrossRef] [PubMed]
- Takasaki, M.; Konoshima, T.; Komatsu, K.; Tokuda, H.; Nishino, H. Anti-Tumor-Promoting Activity of Lignans from the Aerial Part of Saussurea medusa. Cancer Lett. 2000, 158, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Kim, A.R.; Yoo, E.S.; Baik, K.U.; Park, M.H. Immunomodulatory Effect of Arctigenin, a Lignan Compound, on Tumour Necrosis Factor-α and Nitric Oxide Production, and Lymphocyte Proliferation. J. Pharm. Pharmacol. 1999, 51, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Hausott, B.; Greger, H.; Marian, B. Naturally Occurring Lignans Efficiently Induce Apoptosis in Colorectal Tumor Cells. J. Cancer Res. Clin. Oncol. 2003, 129, 569–576. [Google Scholar] [CrossRef]
- Ma, D.; Lu, B.; Feng, C.; Wang, C.; Wang, Y.; Luo, T.; Feng, J.; Jia, H.; Chi, G.; Luo, Y.; et al. Deoxypodophyllotoxin Triggers Parthanatos in Glioma Cells via Induction of Excessive ROS. Cancer Lett. 2016, 371, 194–204. [Google Scholar] [CrossRef]
- Chen, S.; Gao, Y.; Zhou, N.; Liu, J.; Huang, W.; Hui, L.; Jin, Y.; Jin, Y. Carbamates of 4′-Demethyl-4-Deoxypodophyllotoxin- Synthesis, Cytotoxicity and Cell Cycle Effects. Bioorg. Med. Chem. Lett. 2011, 21, 7355–7358. [Google Scholar] [CrossRef]
- Gordaliza, M.; Castro, M.A.; García-Grávalos, M.D.; Ruiz, P.; Del Corral, J.M.M.; San Feliciano, A. Antineoplastic and Antiviral Activities of Podophyllotoxin Related Lignans. Arch. Pharm. 1994, 327, 175–179. [Google Scholar] [CrossRef] [PubMed]
- San Feliciano, A.; Gordaliza, M.; Miguel Del Corral, J.M.; Castro, M.A.; Garcia-Gravalos, M.D.; Ruiz-Lazaro, P. Antineoplastic and Antiviral Activities of Some Cyclolignans. Planta Med. 1993, 59, 246–249. [Google Scholar] [CrossRef]
- You, Y.; Kim, Y.; Nam, N.; Bang, S.; Ahn, B. Alkyl and Carboxylalkyl Esters of 4′-Demethyl-4-Deoxypodophyllotoxin: Synthesis, Cytotoxic, and Antitumor Activity. Eur. J. Med. Chem. 2004, 39, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Khaled, M.; Belaaloui, G.; Jiang, Z.Z.; Zhu, X.; Zhang, L.Y. Deoxypodophyllotoxin, a Semi-Synthetic Compound from Dysosma versipellis, Induces Selective Cell Death in Human Breast Cancer Cell Lines. Med. Chem. Res. 2017, 26, 1241–1258. [Google Scholar] [CrossRef]
- Guerram, M.; Jiang, Z.Z.; Sun, L.; Zhu, X.; Zhang, L.Y. Antineoplastic Effects of Deoxypodophyllotoxin, a Potent Cytotoxic Agent of Plant Origin, on Glioblastoma U-87 MG and SF126 Cells. Pharmacol. Rep. 2015, 67, 245–252. [Google Scholar] [CrossRef]
- Gordaliza, M.; Miguel Del Corral, J.M.; Angeles Castro, M.; García-García, P.A.; San Feliciano, A. Cytotoxic Cyclolignans Related to Podophyllotoxin. Il Farm. 2001, 56, 297–304. [Google Scholar] [CrossRef]
- Moritani, S.; Nomura, M.; Takeda, Y.; Miyamoto, K. Cytotoxic Components of Bardanae Fructus (Goboshi). Biol. Pharm. Bull. 1996, 19, 1515–1517. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, E.C.; O’Boyle, N.M. Colchicine-Binding Site Inhibitors from Chemistry to Clinic: A Review. Pharmaceuticals 2020, 13, 8, Corrected in Pharmaceuticals 2020, 13, 72. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Brauer, C.; Thu, K.L.; Mason, J.M.; Blaser, H.; Bray, M.R.; Mak, T.W. Targeting Mitosis in Cancer: Emerging Strategies. Mol. Cell 2015, 60, 524–536. [Google Scholar] [CrossRef]
- Zang, X.; Wang, G.; Cai, Q.; Zheng, X.; Zhang, J.; Chen, Q.; Wu, B.; Zhu, X.; Hao, H.; Zhou, F. A Promising Microtubule Inhibitor Deoxypodophyllotoxin Exhibits Better Efficacy to Multidrug-Resistant Breast Cancer than Paclitaxel via Avoiding Efflux Transport. Drug Metab. Dispos. 2018, 46, 542–551. [Google Scholar] [CrossRef]
- Shin, S.Y.; Yong, Y.; Lee, Y.H. Effect of Deoxypodophyllotoxin Isolated from Anthriscus sylvestris Roots on the Expression of Cell Cycle-Regulatory Proteins in HeLa Cells. J. Appl. Biol. Chem. 2010, 53, 304–309. [Google Scholar] [CrossRef]
- Malumbres, M.; Barbacid, M. Mammalian Cyclin-Dependent Kinases. Trends Biochem. Sci. 2005, 30, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Khaled, M.; Jiang, Z.Z.; Zhang, L.Y. Deoxypodophyllotoxin: A Promising Therapeutic Agent from Herbal Medicine. J. Ethnopharmacol. 2013, 149, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.R.; Xu, Y.; Jiang, Z.Z.; Guerram, M.; Wang, B.; Zhu, X.; Zhang, L.Y. Deoxypodophyllotoxin Induces G2/M Cell Cycle Arrest and Apoptosis in SGC-7901 Cells and Inhibits Tumor Growth in vivo. Molecules 2015, 20, 1661–1675. [Google Scholar] [CrossRef]
- Wu, M.; Jiang, Z.; Duan, H.; Sun, L.; Zhang, S.; Chen, M.; Wang, Y.; Gao, Q.; Song, Y.; Zhu, X.; et al. Deoxypodophyllotoxin Triggers Necroptosis in Human Non-Small Cell Lung Cancer NCI-H460 Cells. Biomed. Pharmacother. 2013, 67, 701–706. [Google Scholar] [CrossRef]
- MacRae, W.D.; Towers, G.H.N. Biological of Lignans. Phytochemistry 1984, 23, 1207–1220. [Google Scholar] [CrossRef]
- Loike, J.D.; Horwitz, S.B. Effect of VP-16-213 on the Intracellular Degradation of DNA in HeLa Cells. Biochemistry 1976, 15, 5443–5448. [Google Scholar] [CrossRef]
- Grieder, A.; Maurer, R.; Stähelin, H. Effect of an Epipodophyllotoxin Derivative (VP 16-213) on Macromolecular Synthesis and Mitosis in Mastocytoma Cells in vitro. Cancer Res. 1974, 34, 1788–1793. [Google Scholar] [PubMed]
- Huang, C.C.; Hou, Y.; Wang, J.J. Effects of a New Antitumor Agent, Epipodophyllotoxin, on Growth and Chromosomes in Human Hematopoietic Cell Lines. Cancer Res. 1973, 33, 3123–3129. [Google Scholar]
- Lakhani, S.A.; Masud, A.; Kuida, K.; Porter, G.A.; Booth, C.J.; Mehal, W.Z.; Inayat, I.; Flavell, R.A. Caspases 3 and 7: Key Mediators of Mitochondrial Events of Apoptosis. Science 2006, 311, 847–851. [Google Scholar] [CrossRef]
- Ponder, K.G.; Boise, L.H. The Prodomain of Caspase-3 Regulates Its Own Removal and Caspase Activation. Cell Death Discov. 2019, 5, 56. [Google Scholar] [CrossRef]
- Nitulescu, G.M.; Margina, D.; Juzenas, P.; Peng, Q.; Olaru, O.T.; Saloustros, E.; Fenga, C.; Spandidos, D.A.; Libra, M.; Tsatsakis, A.M. Akt Inhibitors in Cancer Treatment: The Long Journey from Drug Discovery to Clinical Use (Review). Int. J. Oncol. 2016, 48, 869–885. [Google Scholar] [CrossRef]
- Luo, H.R.; Hattori, H.; Hossain, M.A.; Hester, L.; Huang, Y.; Lee-Kwon, W.; Donowitz, M.; Nagata, E.; Snyder, S.H. Akt as a Mediator of Cell Death. Proc. Natl. Acad. Sci. USA 2003, 100, 11712–11717. [Google Scholar] [CrossRef]
- Grunt, T.W.; Mariani, G.L. Novel Approaches for Molecular Targeted Therapy of Breast Cancer: Interfering with PI3K/AKT/MTOR Signaling. Curr. Cancer Drug Targets 2013, 13, 188–204. [Google Scholar] [CrossRef] [PubMed]
- Park, B.; Lee, S.A.; Moon, S.M.; Kim, C.S. Anthricin-induced Caspase-dependent Apoptosis through IGF1R-PI3K-AKT Pathway Inhibition in A549 Human Non-small Lung Cancer Cells. Oncol. Rep. 2018, 39, 2769–2776. [Google Scholar] [CrossRef]
- Baserga, R.; Peruzzi, F.; Reiss, K. The IGF-1 Receptor in Cancer Biology. Int. J. Cancer 2003, 107, 873–877. [Google Scholar] [CrossRef]
- Kwak, A.W.; Lee, M.H.; Yoon, G.; Cho, S.S.; Choi, J.S.; Chae, J.I.; Shim, J.H. Deoxypodophyllotoxin, a Lignan from Anthriscus sylvestris, Induces Apoptosis and Cell Cycle Arrest by Inhibiting the Egfr Signaling Pathways in Esophageal Squamous Cell Carcinoma Cells. Int. J. Mol. Sci. 2020, 21, 6854. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Chiara, M.; Mok, T.; Mitsudomi, T. Treatment Strategies and Outcomes for Patients with EGFR-Mutant Non-Small Cell Lung Cancer Resistant to EGFR Tyrosine Kinase Inhibitors: Focus on Novel Therapies. Lung Cancer 2022, 170, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Oh, H.N.; Kwak, A.W.; Kim, E.; Lee, M.H.; Seo, J.H.; Cho, S.S.; Yoon, G.; Chae, J.I.; Shim, J.H. Deoxypodophyllotoxin Inhibits Cell Growth and Induces Apoptosis in Gefitinib-Resistant Non-Small Lung Cancer Cells by Dual-Targeting EGFR and MET. J. Microbiol. Biotechnol. 2021, 31, 559–569. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kang, B.Y.; Jung, S.J.; Kwak, A.W.; Lee, S.O.; Park, J.W. Picropodophyllotoxin Inhibits Cell Growth and Induces Apoptosis in Gefitinib-Resistant Non-Small Lung Cancer Cells by Dual-Targeting EGFR and MET. Biomol. Ther. 2022, 10, 1–10. [Google Scholar] [CrossRef]
- Negut, I.; Grumezescu, V.; Grumezescu, A.M.; Bîrcă, A.C.; Holban, A.M.; Urzica, I.; Avramescu, S.M.; Gălăţeanu, B.; Hudiţă, A. Nanostructured Thin Coatings Containing Anthriscus sylvestris Extract with Dual Bioactivity. Molecules 2020, 25, 3866. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Kuramochi, K.; Imai, T.; Takata, K.; Takehara, M.; Kobayashi, S.; Sakaguchi, K.; Sugawara, F. Podophyllotoxin Directly Binds a Hinge Domain in E2 of HPV and Inhibits an E2/E7 Interaction in vitro. Bioorg. Med. Chem. 2008, 16, 5815–5825. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, A.; Ono, T.; Ishimoto, A.; Dowhanick, J.J.; Frizzell, M.A.; Howley, P.M.; Sakai, H. Mechanisms of Human Papillomavirus E2-Mediated Repression of Viral Oncogene Expression and Cervical Cancer Cell Growth Inhibition. J. Virol. 2000, 74, 3752–3760. [Google Scholar] [CrossRef] [PubMed]
- Bedows, E.; Hatfield, G.M. An Investigation of the Antiviral Activity of Podophyllum peltatum. J. Nat. Prod. 1982, 45, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.A.; Miguel Del Corral, J.M.; Gordaliza, M.; Gómez-Zurita, M.A.; De La Puente, M.L.; Betancur-Galvis, L.A.; Sierra, J.; San Feliciano, A. Synthesis, Cytotoxicity and Antiviral Activity of Podophyllotoxin Analogues Modified in the E-Ring. Eur. J. Med. Chem. 2003, 38, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Hammonds, T.R.; Denyer, S.P.; Jackson, D.E.; Irving, W.L. Studies to Show That with Podophyllotoxin the Early Replicative Stages of Herpes Simplex Virus Type 1 Depend upon Functional Cytoplasmic Microtubules. J. Med. Microbiol. 1996, 45, 167–172. [Google Scholar] [CrossRef] [PubMed]
- MacRae, W.D.; Hudson, J.B.N.; Towers, G.H. The Antiviral Action of Lignans. Planta Med. 1989, 55, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Charlton, J.L. Antiviral Activity of Lignans. J. Nat. Prod. 1998, 61, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Narutaki, K.; Nagaoka, Y.; Hayashi, T.; Uesato, S. Therapeutic Effect of Arctiin and Arctigenin in Immunocompetent and Immunocompromised Mice Infected with Influenza A Virus. Biol. Pharm. Bull. 2010, 33, 1199–1205. [Google Scholar] [CrossRef]
- Eich, E.; Pertz, H.; Kaloga, M.; Schulz, J.; Fesen, M.R.; Mazumder, A.; Pommier, Y. (−)-Arctigenin as a Lead Structure for Inhibitors of Human Immunodeficiency Virus Type-1 Integrase. J. Med. Chem. 1996, 39, 86–95. [Google Scholar] [CrossRef]
- Schröder, H.C.; Merz, H.; Steffen, R.; Müller, W.E.G.; Sarin, P.S.; Trumm, S.; Schulz, J.; Eich, E. Differential in vitro Anti-HIV Activity of Natural Lignans. Z. Naturforsch.—Sect. C J. Biosci. 1990, 45, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Eich, E.; Schulz, J.; Trumm, S.; Sarin, P.S.; Maidof, A.; Merz, H.; Schröder, H.C.; Muller, W.E.G. Lignanolides: Novel In vitro Anti-HIV Active. Planta Med. 1990, 56, 506. [Google Scholar] [CrossRef]
- Kim, Y.; Hollenbaugh, J.A.; Kim, D.H.; Kim, B. Novel PI3K/Akt Inhibitors Screened by the Cytoprotective Function of Human Immunodeficiency Virus Type 1 Tat. PLoS ONE 2011, 6, e21781. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.; Yamakuchi, M.; Masuda, S.; Tokioka, T.; Yamaoka, S.; Maruyama, I.; Kitajima, I. Phosphoinositide-3 Kinase-PKB/Akt Pathway Activation Is Involved in Fibroblast Rat-1 Transformation by Human T-Cell Leukemia Virus Type I Tax. Oncogene 2001, 20, 2514–2526. [Google Scholar] [CrossRef] [PubMed]
- Esteves, F.; Rueff, J.; Kranendonk, M. The Central Role of Cytochrome P450 in Xenobiotic Metabolism—A Brief Review on a Fascinating Enzyme Family. J. Xenobiotics 2021, 11, 94–114. [Google Scholar] [CrossRef] [PubMed]
- Julsing, M.K.; Vasilev, N.P.; Schneidman-Duhovny, D.; Muntendam, R.; Woerdenbag, H.J.; Quax, W.J.; Wolfson, H.J.; Ionkova, I.; Kayser, O. Metabolic Stereoselectivity of Cytochrome P450 3A4 towards Deoxypodophyllotoxin: In Silico Predictions and Experimental Validation. Eur. J. Med. Chem. 2008, 43, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Franklin, M.R. The Enzymic Formation of a Methylenedioxyphenyl Derivative Exhibiting an Isocyanide-like Spectrum with Reduced Cytochrome P-450 in Hepatic Microsomes. Xenobiotica 1971, 1, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kim, Y.; Jin, C.; Lee, S.H.; Kang, M.J.; Jeong, T.C.; Jeong, S.Y.; Kim, D.H.; Yoo, H.H. Inhibitory Effects of Deoxypodophyllotoxin from Anthriscus sylvestris on Human CYP2C9 and CYP3A4. Planta Med. 2010, 76, 701–704. [Google Scholar] [CrossRef]
- Xu, P.; Sun, Q.; Wang, X.; Zhang, S.; An, S.; Cheng, J.; Gao, R.; Xiao, H. Pharmacological Effect of Deoxypodophyllotoxin: A Medicinal Agent of Plant Origin, on Mammalian Neurons. Neurotoxicology 2010, 31, 680–686. [Google Scholar] [CrossRef]
- Djamgoz, M.B.A.; Fraser, S.P.; Brackenbury, W.J. In vivo Evidence for Voltage-Gated Sodium Channel Expression in Carcinomas and Potentiation of Metastasis. Cancers 2019, 11, 1675. [Google Scholar] [CrossRef]
- Lopez-Charcas, O.; Pukkanasut, P.; Velu, S.E.; Brackenbury, W.J.; Hales, T.G.; Besson, P.; Gomora, J.C.; Roger, S. Pharmacological and Nutritional Targeting of Voltage-Gated Sodium Channels in the Treatment of Cancers. iScience 2021, 24, 102270. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, E.A.; Golmohammadi, A.; Rehm, R.; Tetzlaff, C.; Rizzoli, S.O. High-Resolution Analysis of Bound Ca2+ in Neurons and Synapses. Life Sci. Alliance 2023, 7, e202302030. [Google Scholar] [CrossRef] [PubMed]
- Gleichmann, M.; Mattson, M.P. Neuronal Calcium Homeostasis and Dysregulation. Antioxid. Redox Signal. 2011, 14, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.P.; Kim, S.R.; Kim, Y.C. Neuroprotective Dibenzylbutyrolactone Lignans of Torreya nucifera. Planta Med. 2001, 67, 470–472. [Google Scholar] [CrossRef]
- Kim, S.H.; Jang, Y.P.; Sung, S.H.; Kim, C.J.; Kim, J.W.; Kim, Y.C. Hepatoprotective Dibenzylbutyrolactone Lignans of Torreya nucifera against CCI4-Induced Toxicity in Primary Cultured Rat Hepatocytes. Biol. Pharm. Bull. 2003, 26, 1202–1205. [Google Scholar] [CrossRef]
- Kim, K.Y.; Park, K.I.; Lee, S.G.; Baek, S.Y.; Lee, E.H.; Kim, S.C.; Kim, S.H.; Park, S.G.; Yu, S.N.; Oh, T.W.; et al. Deoxypodophyllotoxin in Anthriscus sylvestris Alleviates Fat Accumulation in the Liver via AMP-Activated Protein Kinase, Impeding SREBP-1c Signal. Chem. Biol. Interact. 2018, 294, 151–157. [Google Scholar] [CrossRef]
- Gordaliza, M.; Faircloth, G.T.; Castro, M.A.; Miguel Del Corral, J.M.; López-Vázquez, M.L.; San Feliciano, A. Immunosuppressive Cyclolignans. J. Med. Chem. 1996, 39, 2865–2868. [Google Scholar] [CrossRef]
- Park, H.J.; Hong, S. Anthriscus sylvestris-Derived Extract Induces Th1 and Th17 Cell Differentiation via the Upregulation of IL12 and IL23 Production. Anim. Cells Syst. 2014, 18, 237–243. [Google Scholar] [CrossRef]
- Inamori, Y.; Kato, Y.; Kubo, M.; Baba, K.; Ishida, T.; Nomoto, K.; Kozawa, M. The Biological Actions of Deoxypodophyllotoxin (Anthricin). I. Physiological Activities and Conformational Analysis of Deoxypodophyllotoxin. Chem. Pharm. Bull. 1985, 33, 704–709. [Google Scholar] [CrossRef]
- Russell, G.B.; Singh, P.; Fenemore, P.G. Insect-Control Chemicals from Plants. III. Toxic Lignans from Lihocedrus hidwillii. Aust. J. Biol. Sci. 1976, 29, 99–103. [Google Scholar] [CrossRef]
- Inamori, Y.; Kato, Y.; Kubo, M.; Baba, K.; Matsuyama, Y.; Sakai, M.; Kozawa, M. Mechanisms of Insecticidal Action of Deoxypodophyllotoxin (Anthricin). I.1) Distribution of Deoxypodophyllotoxin in Tissues of the 5th Instar Larvae of Silkworm, Bombyx mori LINNE.Pdf. Chem. Pharm. Bull. 1983, 31, 4464–4468. [Google Scholar] [CrossRef]
- Inamori, Y.; Kato, Y.; Kubo, M.; Waku, Y.; Hayashiya, K.; Sakai, M.; Baba, K.; Kozawa, M. Mechanisms of Insecticidal Action of Deoxypodophyllotoxin (Anthricin). II. Histopathological Studies on Tissues of Silkworm Larvae Intoxicated by Deoxypodophyllotoxin. Chem. Pharm. Bull. 1984, 32, 2015–2019. [Google Scholar] [CrossRef]
- Moss, G.P. Nomenclature of Lignans and Neolignans (IUPAC Recommendations 2000). Pure Appl. Chem. 2000, 72, 1493–1523. [Google Scholar] [CrossRef]
- Terada, T.; Fujimoto, K.; Nomura, M.; Yamashita, J.; Kobunai, T.; Takeda, S.; Wierzba, K.; Yamada, Y.; Yamaguchi, H. Antitumor Agents. I. DNA Topoisomerase II Inhibitory Activity and the Structural Relationship of Podophyllotoxin Derivatives as Antitumor Agents. Chem. Pharm. Bull. 1992, 40, 2720–2727. [Google Scholar] [CrossRef]
- Castro, A.; Del Corral, J.M.M.; Gordaliza, M.; Grande, C.; Gómez-Zurita, A.; García-Grávalos, D.; San Feliciano, A. Synthesis and Cytotoxicity of Podophyllotoxin Analogues Modified in the A Ring. Eur. J. Med. Chem. 2003, 38, 65–74. [Google Scholar] [CrossRef]
- Gordaliza, M.; Castro, M.A.; Miguel del Corral, J.M.; Lopez-Vazquez, M.L.; San Feliciano, A.; Faircloth, G.T. In vivo Immunosuppressive Activity of Some Cyclolignans. Bioorg. Med. Chem. Lett. 1997, 7, 2781–2786. [Google Scholar] [CrossRef]
- Thurston, L.S.; Imakura, Y.; Haruna, M.; Li, D.; Liu, Z.; Liu, S.; Cheng, Y.; Lee, K. Antitumor Agents. 100. Inhibition of Human DNA Topoisomerase II by Cytotoxic Ether and Ester Derivatives of Podophyllotoxin and Alpha-Peltatin. Am. Chem. Soc. 1989, 32, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Van Uden, W.; Bos, J.A.; Boeke, G.M.; Woerdenbag, H.J.; Pras, N. The Large-Scale Isolation of Deoxypodophyllotoxin from Rhizomes of Anthriscus sylvestris Followed by Its Bioconversion into 5- Methoxypodophyllotoxin β-D-Glucoside by Cell Cultures of Linum flavum. J. Nat. Prod. 1997, 60, 401–403. [Google Scholar] [CrossRef]
- Hu, H.; Wang, Z.; Liu, S.; Cheng, Y.; Lee, K. Antitumor Agents. 123. Synthesis and Human DNA Topoisomerase II Inhibitory Activity of 2′-Chloro Derivatives of Etoposide and 4&-(Arylamino)-4′-O-Demethylpodophyllotoxins. J. Med. Chem. 1992, 35, 866–871. [Google Scholar]
- Brewer, C.F.; Loike, J.D.; Horwitz, S.B. Conformational Analysis of Podophyllotoxin and Its Congerers. Structure-Activity Relationships in Microtubule Assembly. J. Med. Chem. 1979, 22, 215–221. [Google Scholar] [CrossRef]
- Kelly, M.G.; Hartwell, J.L. The Biological Effects and the Chemical Composition of Podophyllin. A Review. J. Natl. Cancer Inst. 1954, 14, 967–1010. [Google Scholar] [PubMed]
- Rithner, C.D.; Bushweller, C.H. Dynamic Nuclear Magnetic Resonance and Empirical Force Field Studies of Podophyllotoxin. J. Org. Chem. 1983, 48, 1491–1495. [Google Scholar] [CrossRef]
- Alizadeh, B.H.; Emami, S.; Dehghan, G.; Foroumadi, A.; Shafiee, A. Synthesis of Cytotoxic Isodeoxypodophyllotoxin Analogs. J. Heterocycl. Chem. 2017, 54, 539–545. [Google Scholar] [CrossRef]
- Zhu, X.; Fu, J.; Tang, Y.; Gao, Y.; Zhang, S.; Guo, Q. Design and Synthesis of Novel 4′-Demethyl-4-Deoxypodophyllotoxin Derivatives as Potential Anticancer Agents. Bioorg. Med. Chem. Lett. 2016, 26, 1360–1364. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Name | R1 | R2 | R3 | R4 | R5 | R6 | R7 | References |
| (−)-deoxypodorhizone (yatein) | OCH2O | OMe | OMe | OMe | αH | βH | [26,29,42,51,53,54,55,65,73,77,79] | |
| matairesinol | OMe | OH | OMe | OH | H | αH | βH | [53,73,77] |
| thujaplicatin | OMe | OH | OMe | OH | OH | αH | βH | [77] |
| 5-O-methylthujaplicatin | OMe | OH | OMe | OH | OMe | αH | βH | [77,80] |
| 4,5-di-O-methylthujaplicatin * (thujaplicatin-3,4-dimethylether, hernanol) | OMe | OH | OMe | OMe | OMe | αH | βH | [73,77,80] |
| dimethylthujaplicatin methyl ether (trimethylthujaplicatin) * | OMe | OMe | OMe | OMe | OMe | αH | βH | [29,73] |
| pluviatolide * | OCH2O | OMe | OH | H | αH | βH | [53,73,77] | |
| bursehernin | OCH2O | OMe | OMe | H | αH | βH | [29,52,53,73,77] | |
| arctigenin | OMe | OMe | OMe | OH | H | αH | βH | [29] |
| (−)-hinokinin | OCH2O | OCH2O | H | αH | βH | [29,53,65] | ||
| dimethylmatairesinol (methylarctigenin) | OMe | OMe | OMe | OMe | H | αH | βH | [29,73] |
| 5-methoxyguayaraol * | OH | OH | OMe | OMe | OMe | αH | βH | [73] |
 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Name | R1 | R2 | R3 | R4 | R5 | R6 | R7 | R8 | R9 | References |
| 7′-hydroxyyatein | OCH2O | OMe | OMe | OMe | αH | βH | OH | H | [29] | |
| 8-hydroxy-8-epi-yatein | OCH2O | OMe | OMe | OMe | αH | αOH | H | H | [73] | |
| (−)-podorhizol (7-hydroxyyatein) | OCH2O | OMe | OMe | OMe | αH | βH | H | βOH | [73] | |
| (+)-podorhizon (7-oxoyatein) | OCH2O | OMe | OMe | OMe | αH | βH | H | =O | [73] | |
| 8-hydroxy-8′-epi-pluviatolide | OCH2O | OMe | OH | H | βH | βOH | H | H | [73] | |
| guayadequiol | OCH2O | OMe | OMe | H | βH | βOH | H | H | [73] | |
| epiwikstromol dimethyl ether (8-hydroxy-8-epi-matairesinol) | OMe | OMe | OMe | OMe | H | βH | βOH | H | H | [73] |
| wikstromol dimethyl ether | OMe | OMe | OMe | OMe | H | βH | αOH | H | H | [73] |
| 8-hydroxy-trimethoxylignano-9,9′-lactone * | (OMe, H) | OMe | OMe | H | H | OH | H | H | [73] | |
| 3′,8-dihydroxy-3,4,4′-trimethoxylignano-9,9′-lactone * | OMe | OH | OMe | OMe | H | H | OH | H | H | [73] |
 | |||||||
|---|---|---|---|---|---|---|---|
| Name | R1 | R2 | R3 | R4 | R5 | R6 | References |
| anhydropodorhizol (nemerosin) | OCH2O | OMe | OMe | OMe | H | [26,29,42,51,53,54,55,62,73,79] | |
| isochaihulactone | OCH2O | OMe | OMe | OMe | H | [73] | |
| isosuchilactone | OMe | OMe | OCH2O | H | H | [51] | |
| jatrophan (βH in lactone) | OMe | OMe | OCH2O | H | H | [51] | |
| sylvestrin (βH in lactone) | OCH2O | OMe | OMe | OMe | H | [42] | |
| kaerophyllin | OCH2O | OMe | OMe | H | H | [51,73,79] | |
| isokaerophyllin | OCH2O | OMe | OMe | H | H | [73,79] | |
| 7′-hydroxyanhydropodorhizol | OCH2O | OMe | OMe | OMe | OH | [29] | |
| 3,4,5-trimethoxy-3′,4′-dihydroxylign-7-eno-9,9′-lactone * | OH | OH | OMe | OMe | OMe | H | [73,79] |
| (E)-3′-demethyljatrophan | OH | OMe | OCH2O | H | H | [73] | |
| (Z)-3′-demethyljatrophan | OH | OMe | OCH2O | H | H | [73] | |
| (E)-7,8-didehydro-dimethylmatairesinol | OMe | OMe | OMe | OMe | H | H | [73] |
| (Z)-7,8-didehydro-dimethylmatairesinol | OMe | OMe | OMe | OMe | H | H | [73] |
| 7,8-didehydroguayarol * | OH | OH | OMe | OMe | H | H | [73] |
| 7,8-didehydroisoarctigenin * | OMe | OH | OMe | OMe | H | H | [73] |
 | ||||||
|---|---|---|---|---|---|---|
| Name | R1 | R2 | R3 | R4 | R5 | References |
| morelensin | OMe | H | βH | αH | H | [29,52,65,73] |
| (−)-deoxypodophyllotoxin (anthricin, DPT) | OMe | OMe | βH | αH | H | [26,29,37,42,50,51,52,53,54,55,57,58,65,68,73,79,83,84,85,86,87,88,89,90,91] |
| isoanthricin | OMe | OMe | βH | αH | H | [50,90] |
| deoxypicropodophyllotoxin | OMe | OMe | αH | αH | H | [42,90] |
| α-peltatin | OH | OMe | βH | αH | OH | [54] |
| β-peltatin | OMe | OMe | βH | αH | OH | [54] |
| β-peltatin A methyl ether | OMe | OMe | βH | αH | OMe | [54] |
 | |||||
|---|---|---|---|---|---|
| Name | R1 | R2 | R3 | R4 | References |
| podophyllotoxin | OMe | βH | αH | αOH | [29,54,73,79] |
| picropodophyllotoxin | OMe | αH | αH | αOH | [42,73] |
| angeloylpodophyllotoxin | OMe | βH | αH | α-C5H7O2 | [29,42,52,54,55,73,79] |
| acetylpodophyllotoxin | OMe | βH | αH | α-CH3COO | [73] |
| 5′-demethoxypodophyllotoxin (7-hydroxymorelensin) | H | βH | αH | αOH | [73,79] |
| podophyllotoxone | OMe | βH | αH | =O | [54,55,73,79] |
| picropodophyllotoxone (picropodophyllone) | OMe | αH | αH | =O | [73,79] |
| isopicropodophyllotoxone (isopicropodophyllone) | OMe | βH | βH | =O | [54,55,73] |
| 5′-demethoxypodophyllotoxone | H | βH | αH | =O | [73] |
| 5′-demethoxypicropodophyllotoxone (5′-demethoxypicropodophyllone) | H | αH | αH | =O | [73] |
| 5′-demethoxyisopicropodophyllotoxone (5′-demethoxyisopicropodophyllone) | H | βH | βH | =O | [73] |
| 4-hydroxy-3′,4′,5-trimethoxy-7-oxo-2,7′-cyclolignano-9′,9-lactone * | H | H | H | =O | [73] |
| Compound | Cell Line | IC50 (µmol/L) | Assay | References |
|---|---|---|---|---|
| deoxypodophyllotoxin | MK-1 | 0.0025 | MTT | [51] |
| 0.055 | MTT | [65] | ||
| HeLa | 0.0013 | MTT | [51] | |
| 0.083 | MTT | [65] | ||
| 4.97 | CCK-8 | [124] | ||
| 47.03 | MTT | [90] | ||
| B16F10 | 0.0018 | MTT | [51] | |
| 0.21 | MTT | [65] | ||
| Colo205 | 0.24 | MTT | [52] | |
| K562 | 0.046 | MTT | [52] | |
| A-549 | 1.38 | MTT | [124] | |
| <0.006 | CVS | [125] | ||
| 0.0063 | CVS | [126] | ||
| 0.030 | SRB | [127] | ||
| 0.053 | SRB | [112] | ||
| SiHa | 6.01 | MTT | [124] | |
| HL-60 | 0.47 | CCK-8 | [124] | |
| HepG2 | 30.75 | MTT | [90] | |
| MG-63 | 36.10 | MTT | [90] | |
| B16 | 108.67 | MTT | [90] | |
| MCF-7 | 0.011 | MTT | [128] | |
| MDA-MB-231 | 0.020 | MTT | [128] | |
| 0.0218 | MTT | [129] | ||
| SF126 | 0.014 | MTT | [129] | |
| U-87 MG | 0.0151 | MTT | [129] | |
| SGC-7901 | 0.0197 | MTT | [129] | |
| BGC-823 | 0.0267 | MTT | [129] | |
| HO-8910 | 0.0212 | MTT | [129] | |
| SK-0V-3 | 0.0252 | MTT | [129] | |
| HT-29 | 0.0187 | MTT | [129] | |
| 0.006 | CVS | [125] | ||
| 0.0063 | CVS | [126] | ||
| JeG-3 | 0.0253 | MTT | [129] | |
| P-388 | 0.010 | CVS | [125] | |
| 0.0063 | CVS | [126] | ||
| SK-MEL-2 | 0.023 | SRB | [127] | |
| 0.011 | SRB | [112] | ||
| yatein | MK-1 | 0.75 | MTT | [51] |
| 1.85 | MTT | [65] | ||
| HeLa | 2.00 | MTT | [51] | |
| 3.15 | MTT | [65] | ||
| B16F10 | 0.87 | MTT | [51] | |
| 4.03 | MTT | [65] | ||
| A-549 | 1.1 | SRB | [112] | |
| SK-MEL-2 | 1.4 | SRB | [112] | |
| nemerosin | MK-1 | 1.76 | MTT | [51] |
| HeLa | 1.01 | MTT | [51] | |
| B16F10 | 1.76 | MTT | [51] | |
| podophyllotoxin | MK-1 | 0.014 | MTT | [51] |
| HeLa | 0.006 | MTT | [51] | |
| B16F10 | 0.0024 | MTT | [51] | |
| A-549 | 0.012 | CVS | [125,130] | |
| HT-29 | 0.024 | CVS | [125,130] | |
| P-388 | 0.012 | CVS | [125,130] | |
| hinokinin | MK-1 | 4.72 | MTT | [65] |
| HeLa | 7.29 | MTT | [65] | |
| B16F10 | 7.68 | MTT | [65] | |
| morelensin | MK-1 | 0.24 | MTT | [65] |
| HeLa | 0.14 | MTT | [65] | |
| B16F10 | 0.23 | MTT | [65] | |
| Colo205 | >50 | MTT | [52] | |
| K562 | 44.70 | MTT | [52] | |
| bursehernin | Colo205 | 44.45 | MTT | [52] |
| K562 | 1.15 | MTT | [52] | |
| angeloylpodophyllotoxin | Colo205 | 0.19 | MTT | [52] |
| K562 | 0.037 | MTT | [52] | |
| arctigenin | AGS | 85.75 | MTT | [116] |
| Hepa 1c1c7 | 311.02 | MTT | [116] | |
| HL-60 | 0.180 | MTT | [117] | |
| HepG2 | 3.49 | MTT | [131] | |
| MH60 | 1.0 | MTT | [118] | |
| matairesinol | Hepa 1c1c7 | 246.65 | MTT | [116] |
| HL-60 | 0.114 | MTT | [117] | |
| MH60 | 8.4 | MTT | [118] | |
| isoanthricin | HeLa | >377 | MTT | [90] |
| HepG2 | >377 | MTT | [90] | |
| MG-63 | 87.14 | MTT | [90] | |
| B16 | 165.13 | MTT | [90] | |
| deoxypicropodophyllotoxin | HeLa | 296.48 | MTT | [90] |
| A-549 | 0.13 | CVS | [125] | |
| 0.063 | CVS | [126] | ||
| HepG2 | 178.17 | MTT | [90] | |
| HT-29 | 0.06 | CVS | [125] | |
| 0.063 | CVS | [126] | ||
| P-388 | 0.10 | CVS | [125] | |
| 0.063 | CVS | [126] | ||
| acetylpodophyllotoxin | A-549 | 0.625 | CVS | [126] |
| HT-29 | 0.625 | CVS | [126] | |
| P-388 | 0.625 | CVS | [126] | |
| podophyllotoxon | A-549 | 1.8 | CVS | [126,130] |
| HT-29 | 1.8 | CVS | [126,130] | |
| P-388 | 1.8 | CVS | [126,130] | |
| picropodophyllotoxon | A-549 | 12.0 | CVS | [126] |
| HT-29 | 12.0 | CVS | [126] | |
| P-388 | 12.0 | CVS | [126] | |
| isopicropodophyllotoxone | A-549 | 12.1 | CVS | [130] |
| HT-29 | 12.1 | CVS | [130] | |
| P-388 | 6.0 | CVS | [130] | |
| MEL-28 | 12.1 | CVS | [130] | |
| β-peltatin A methyl ether | A-549 | 0.0097 | CVS | [126] |
| HT-29 | 0.0097 | CVS | [126] | |
| P-388 | 0.0097 | CVS | [126] | |
| picropodophyllotoxin | A-549 | 6.0 | CVS | [126,130] |
| HT-29 | 6.0 | CVS | [126,130] | |
| P-388 | 6.0 | CVS | [126,130] | |
| MEL-28 | 6.0 | CVS | [130] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berežni, S.; Mimica-Dukić, N.; Domina, G.; Raimondo, F.M.; Orčić, D. Anthriscus sylvestris—Noxious Weed or Sustainable Source of Bioactive Lignans? Plants 2024, 13, 1087. https://doi.org/10.3390/plants13081087
Berežni S, Mimica-Dukić N, Domina G, Raimondo FM, Orčić D. Anthriscus sylvestris—Noxious Weed or Sustainable Source of Bioactive Lignans? Plants. 2024; 13(8):1087. https://doi.org/10.3390/plants13081087
Chicago/Turabian StyleBerežni, Sanja, Neda Mimica-Dukić, Gianniantonio Domina, Francesco Maria Raimondo, and Dejan Orčić. 2024. "Anthriscus sylvestris—Noxious Weed or Sustainable Source of Bioactive Lignans?" Plants 13, no. 8: 1087. https://doi.org/10.3390/plants13081087
APA StyleBerežni, S., Mimica-Dukić, N., Domina, G., Raimondo, F. M., & Orčić, D. (2024). Anthriscus sylvestris—Noxious Weed or Sustainable Source of Bioactive Lignans? Plants, 13(8), 1087. https://doi.org/10.3390/plants13081087