
Fine Mapping of qAL5.2 Controlling Anther Length in Oryza sativa

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
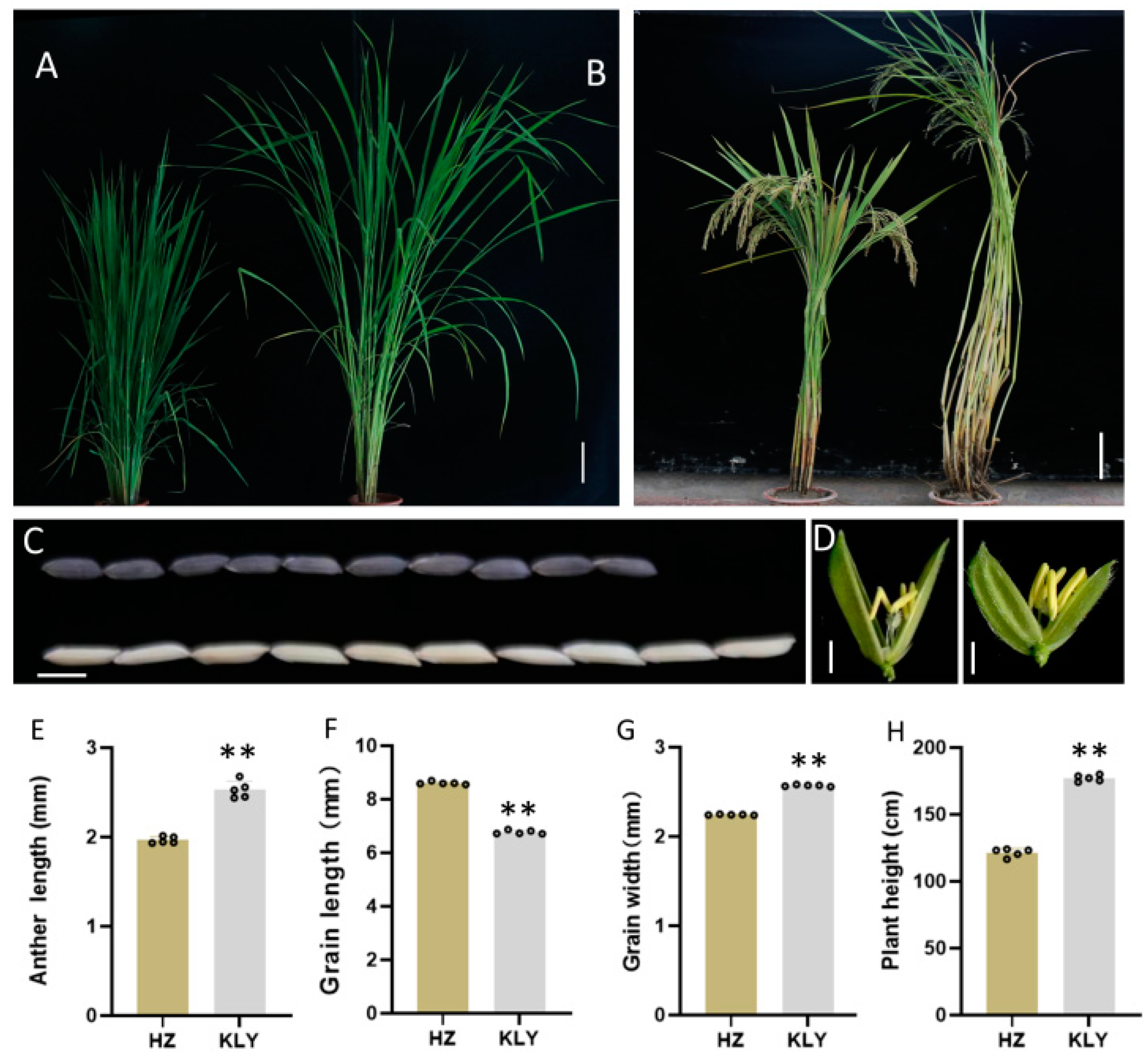
2.1. Anther Morphology of NIL-HZ and NIL-KLY
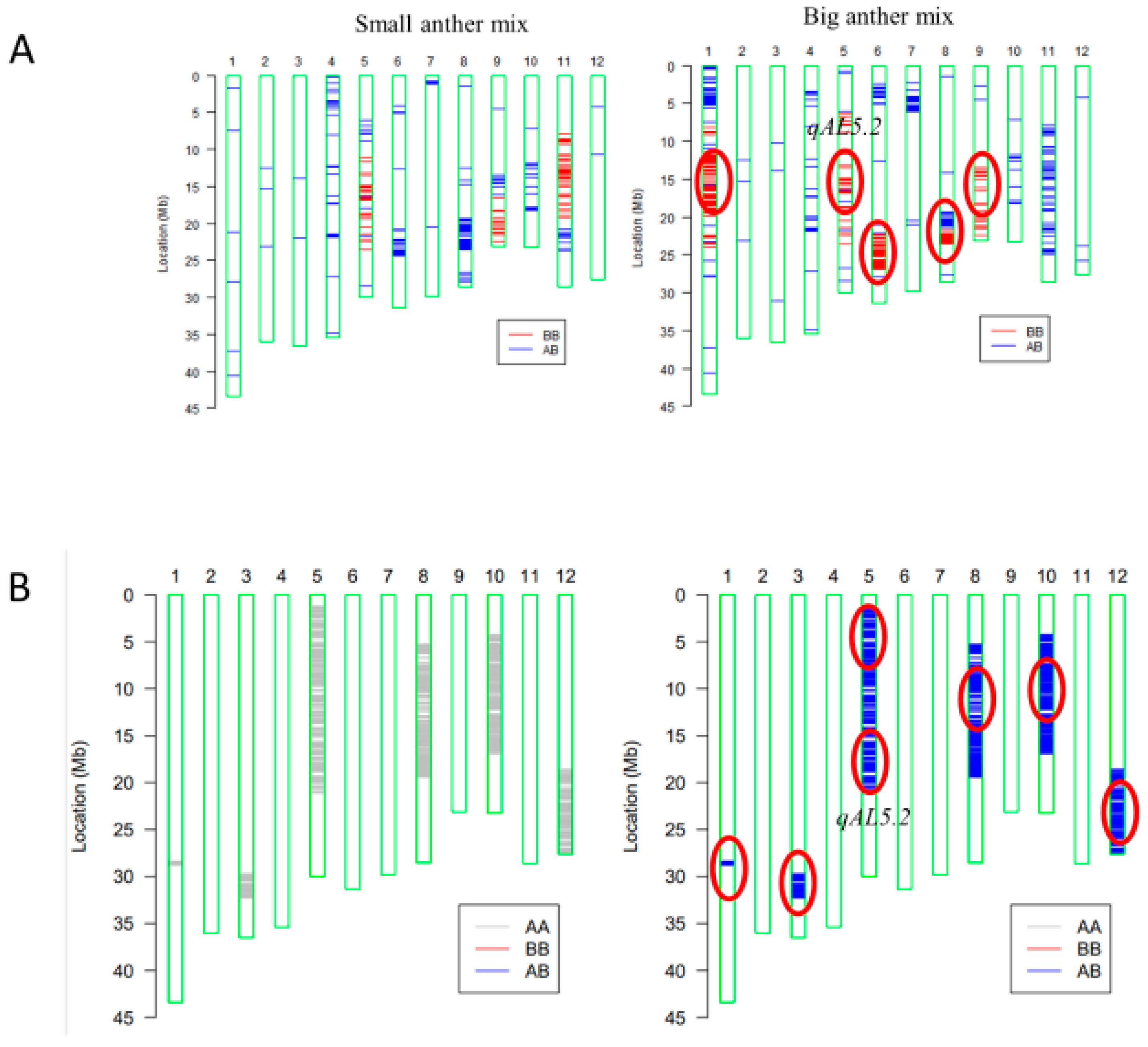
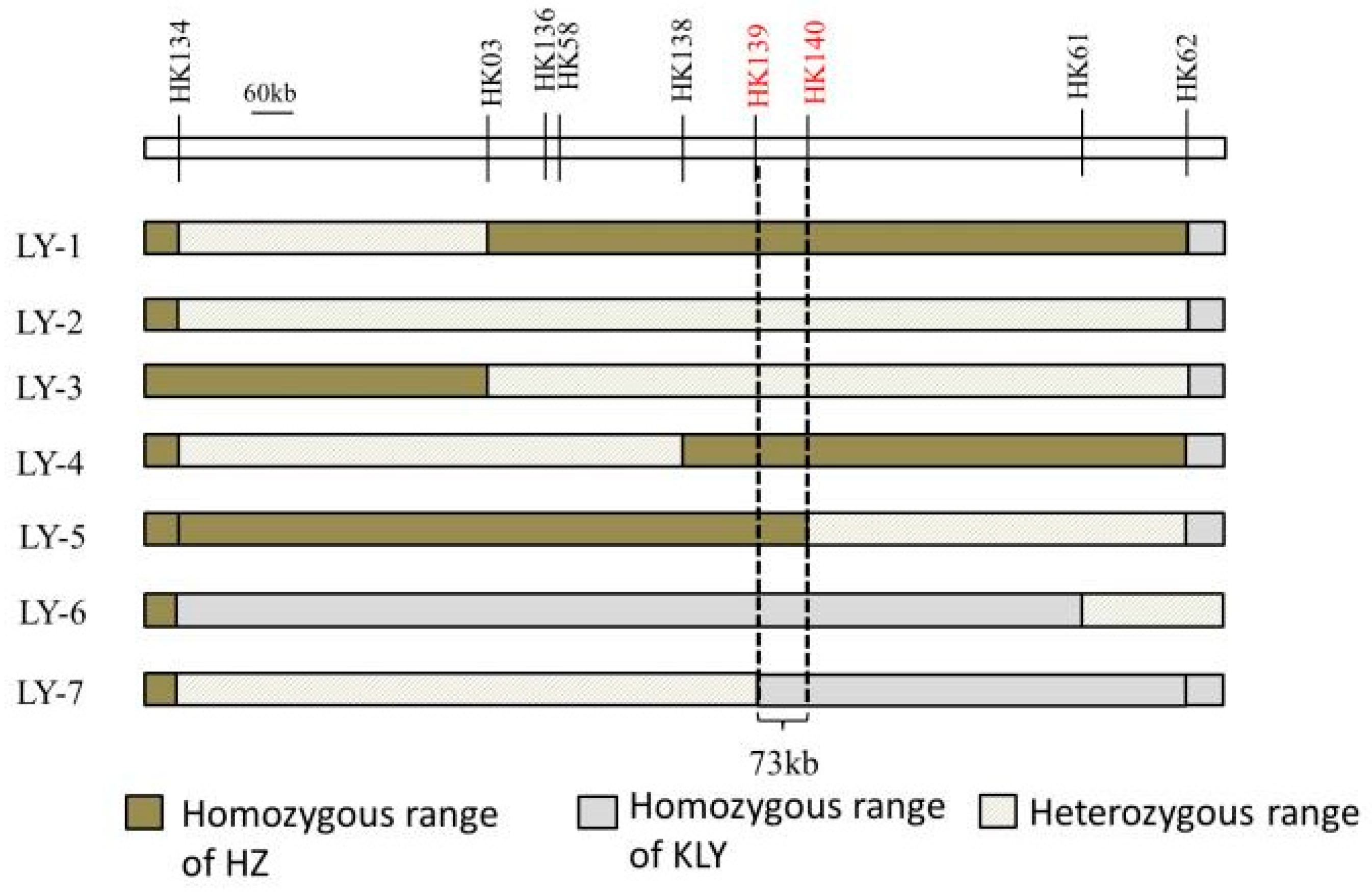
2.2. Effects of qAL5.2 for Anther Length
2.3. Enrichment Analysis of RNA-Seq
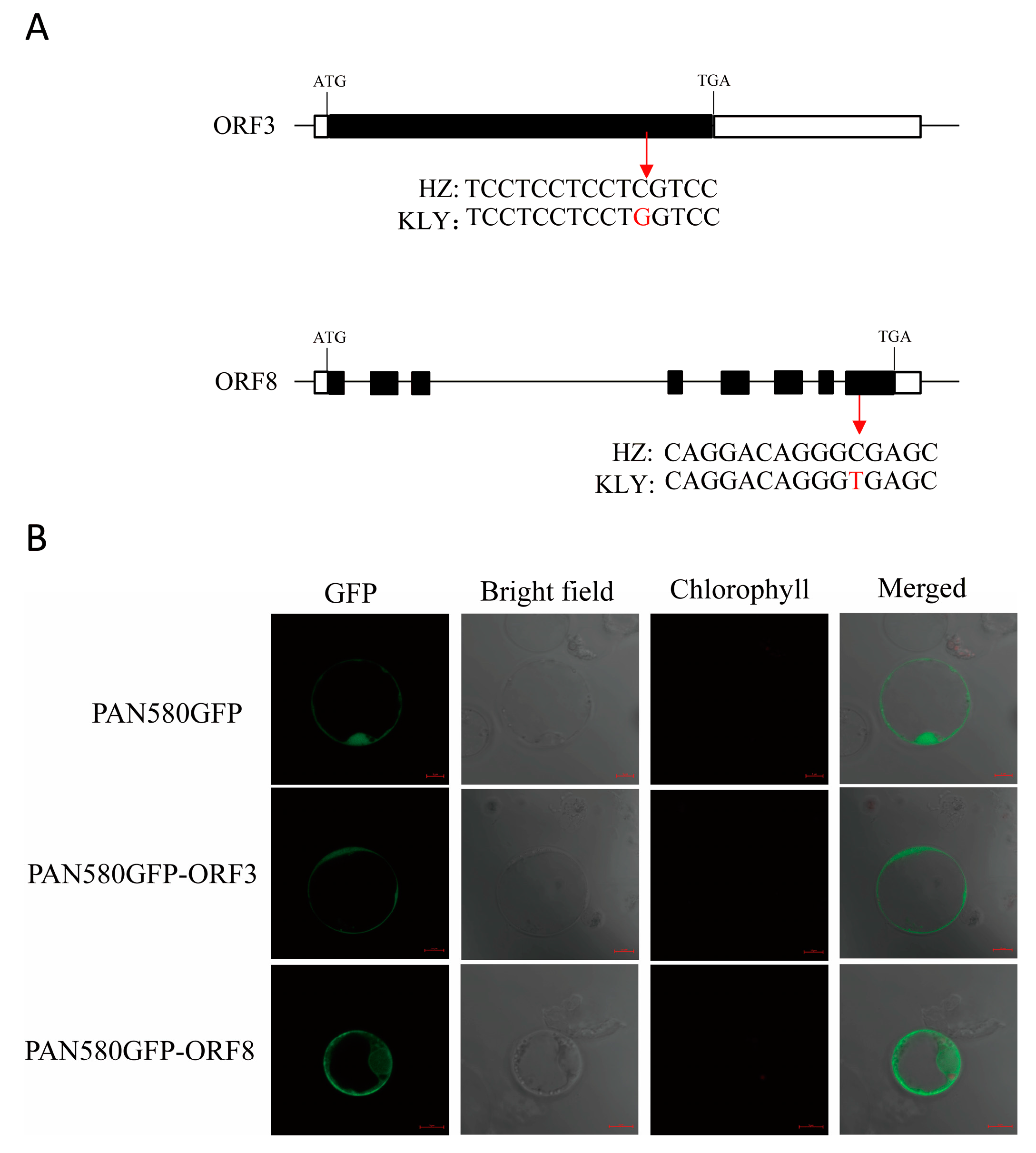
2.4. Candidate Gene Analysis of qAL5.2
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Measurement of Traits
4.3. Genotype Analysis
4.4. Genetic Mapping
4.5. Transcriptomics Analysis
4.6. Subcellular Localization of qAL5
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Qian, Q.; Zhang, F.; Xin, Y. Yuan Longping and Hybrid Rice Research. Rice 2021, 14, 101. [Google Scholar] [CrossRef]
- Liao, C.; Yan, W.; Chen, Z.; Xie, G.; Deng, X.W.; Tang, X. Innovation and development of the third-generation hybrid rice technology. Crop J. 2021, 9, 693–701. [Google Scholar] [CrossRef]
- Cheng, S.H.; Zhuang, J.Y.; Fan, Y.Y.; Du, J.H.; Cao, L.Y. Progress in Research and Development on Hybrid Rice: A Super-domesticate in China. Ann. Bot. 2007, 100, 959–966. [Google Scholar] [CrossRef]
- Luo, D.; Xu, H.; Liu, Z.; Guo, J.; Li, H.; Chen, L.; Fang, C.; Zhang, Q.; Bai, M.; Yao, N.; et al. A detrimental mitochondrial-nuclear interaction causes cytoplasmic male sterility in rice. Nat. Genet. 2013, 45, 573–577. [Google Scholar] [CrossRef]
- Tang, H.; Luo, D.; Zhou, D.; Zhang, Q.; Tian, D.; Zheng, X.; Chen, L.; Liu, Y.-G. The Rice Restorer Rf4 for Wild-Abortive Cytoplasmic Male Sterility Encodes a Mitochondrial-Localized PPR Protein that Functions in Reduction of WA352 Transcripts. Mol. Plant 2014, 7, 1497–1500. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Zhang, D. Molecular Control of Male Fertility for Crop Hybrid Breeding. Trends Plant Sci. 2018, 23, 53–65. [Google Scholar] [CrossRef]
- Fujii, S.; Toriyama, K. Suppressed expression of RETROGRADE-REGULATED MALE STERILITY restores pollen fertility in cytoplasmic male sterile rice plants. Proc. Natl. Acad. Sci. USA 2009, 106, 9513–9518. [Google Scholar] [CrossRef]
- Malik, T.U.H.; Baba, A. Techniques hybrid seed production in rice. J. Pharmacogn. Phytochem. 2018, 7, 962–967. [Google Scholar]
- Li, J.; Luo, X.; Zhou, K. Research and development of hybrid rice in China. Plant Breed. 2023, 143, 96–104. [Google Scholar] [CrossRef]
- Ying, J.; Qin, Y.; Zhang, F.; Duan, L.; Cheng, P.; Yin, M.; Wang, Y.; Tong, X.; Huang, J.; Li, Z.; et al. A weak allele of TGW5 enables greater seed propagation and efficient size-based seed sorting for hybrid rice production. Plant Commun. 2024, 5, 100811. [Google Scholar] [CrossRef]
- Rubiyo; Satoto; Widyastuti, Y.; Indrawanto, C. Hybrid rice development in Indonesia: Constraints and opportunities. E3S Web Conf. 2021, 306, 01047. [Google Scholar] [CrossRef]
- Tazib, T.; Kobayashi, Y.; Koyama, H.; Matsui, T. QTL analyses for anther length and dehiscence at flowering as traits for the tolerance of extreme temperatures in rice (Oryza sativa L.). Euphytica 2014, 203, 629–642. [Google Scholar] [CrossRef]
- Khumto, S.; Pusadee, T.; Olsen, K.M.; Jamjod, S. Genetic relationships between anther and stigma traits revealed by QTL analysis in two rice advanced-generation backcross populations. Euphytica 2017, 214, 5. [Google Scholar] [CrossRef]
- Ogami, T.; Yasui, H.; Yoshimura, A.; Yamagata, Y. Identification of Anther Length QTL and Construction of Chromosome Segment Substitution Lines of Oryza longistaminata. Plants 2019, 8, 388. [Google Scholar] [CrossRef]
- Uga, Y.; Siangliw, M.; Nagamine, T.; Ohsawa, R.; Fujimura, T.; Fukuta, Y. Comparative mapping of QTLs determining glume, pistil and stamen sizes in cultivated rice (Oryza sativa L.). Plant Breed. 2010, 129, 657–669. [Google Scholar] [CrossRef]
- Xiong, L.Z.; Liu, K.D.; Dai, X.K.; Xu, C.G.; Zhang, Q. Identification of genetic factors controlling domestication-related traits of rice using an F2 population of a cross between Oryza sativa and O. rufipogon. Theor. Appl. Genet. 1999, 98, 243–251. [Google Scholar] [CrossRef]
- Shen, G.; Sun, W.; Chen, Z.; Shi, L.; Hong, J.; Shi, J. Plant GDSL Esterases/Lipases: Evolutionary, Physiological and Molecular Functions in Plant Development. Plants 2022, 11, 468. [Google Scholar] [CrossRef]
- Nardini, M.; Dijkstra, B.W. Alpha/beta hydrolase fold enzymes: The family keeps growing. Curr. Opin. Struct. Biol. 1999, 9, 732–737. [Google Scholar] [CrossRef]
- Eastmond, P.J. SUGAR-DEPENDENT1 Encodes a Patatin Domain Triacylglycerol Lipase That Initiates Storage Oil Breakdown in Germinating Arabidopsis Seeds. Plant Cell 2006, 18, 665–675. [Google Scholar] [CrossRef]
- Zhu, J.; Lou, Y.; Shi, Q.-S.; Zhang, S.; Zhou, W.-T.; Yang, J.; Zhang, C.; Yao, X.-Z.; Xu, T.; Liu, J.-L.; et al. Slowing development restores the fertility of thermo-sensitive male-sterile plant lines. Nat. Plants 2020, 6, 360–367. [Google Scholar] [CrossRef]
- Kim, H.U.; Hsieh, K.; Ratnayake, C.; Huang, A.H.C. A Novel Group of Oleosins Is Present Inside the Pollen of Arabidopsis. J. Biol. Chem. 2002, 277, 22677–22684. [Google Scholar] [CrossRef]
- Ha, C.M.; Escamilla-Trevino, L.; Yarce, J.C.S.; Kim, H.; Ralph, J.; Chen, F.; Dixon, R.A. An essential role of caffeoyl shikimate esterase in monolignol biosynthesis in Medicago truncatula. Plant J. 2016, 86, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Barros, J.; Escamilla-Trevino, L.; Song, L.; Rao, X.; Serrani-Yarce, J.C.; Palacios, M.D.; Engle, N.; Choudhury, F.K.; Tschaplinski, T.J.; Venables, B.J.; et al. 4-Coumarate 3-hydroxylase in the lignin biosynthesis pathway is a cytosolic ascorbate peroxidase. Nat. Commun. 2019, 10, 1994. [Google Scholar] [CrossRef]
- Aya, K.; Ueguchi-Tanaka, M.; Kondo, M.; Hamada, K.; Yano, K.; Nishimura, M.; Matsuoka, M. Gibberellin Modulates Anther Development in Rice via the Transcriptional Regulation of GAMYB. Plant Cell 2009, 21, 1453–1472. [Google Scholar] [CrossRef]
- Marchant, D.B.; Walbot, V. Anther development—The long road to making pollen. Plant Cell 2022, 34, 4677–4695. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef]
- Cheng, Y.C.; Li, G.; Yin, M.; Adegoke, T.V.; Wang, Y.F.; Tong, X.H.; Zhang, J.; Ying, J.Z. Verification and dissection of one quantitative trait locus for grain size and weight on chromosome 1 in rice. Sci. Rep. 2021, 11, 18252. [Google Scholar] [CrossRef]
- Zhou, Q.; Hu, X. Systemic Stress and Recovery Patterns of Rice Roots in Response to Graphene Oxide Nanosheets. Environ. Sci. Technol. 2017, 51, 2022–2030. [Google Scholar] [CrossRef]
- Li, Z.; Wei, X.; Tong, X.; Zhao, J.; Liu, X.; Wang, H.; Tang, L.; Shu, Y.; Li, G.; Wang, Y.; et al. The OsNAC23-Tre6P-SnRK1a feed-forward loop regulates sugar homeostasis and grain yield in rice. Mol. Plant 2022, 15, 706–722. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Heterozygous Interval | LOD | A | D | R2 |
|---|---|---|---|---|---|
| LY-1 | HK134–HK03 | ns | - | - | - |
| LY-2 | HK134–HK62 | 17.54 | 0.12 | 0.01 | 35.73 |
| LY-3 | HK03–HK62 | 10.19 | 0.08 | - | 25.10 |
| LY-4 | HK134–HK138 | ns | - | - | - |
| LY-5 | HK140–HK62 | ns | - | - | - |
| LY-6 | HK61–HK62 | ns | - | - | - |
| LY-7 | HK134–HK139 | ns | - | - | - |
| ORF | ID | Gene Product | The Highest Expression |
|---|---|---|---|
| ORF1 | LOC_Os05g29900 | expressed protein | Seed-S4 |
| ORF2 | LOC_Os05g29910 | retrotransposon protein, putative, expressed | No |
| ORF3 | LOC_Os05g29920 | expressed protein | Seed-S5 |
| ORF4 | LOC_Os05g29930 | late embryogenesis abundant protein, expressed | SAM |
| ORF5 | LOC_Os05g29940 | expressed protein | No |
| ORF6 | LOC_Os05g29950 | expressed protein | No |
| ORF7 | LOC_Os05g29960 | expressed protein | No |
| ORF8 | LOC_Os05g29974 | lipase, putative, expressed | Anther |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Yu, Z.; Tong, X.; Chang, L.; Huang, J.; Wang, Y.; Ying, J.; Li, X.; Ni, S.; Zhang, J. Fine Mapping of qAL5.2 Controlling Anther Length in Oryza sativa. Plants 2024, 13, 1130. https://doi.org/10.3390/plants13081130
Liu X, Yu Z, Tong X, Chang L, Huang J, Wang Y, Ying J, Li X, Ni S, Zhang J. Fine Mapping of qAL5.2 Controlling Anther Length in Oryza sativa. Plants. 2024; 13(8):1130. https://doi.org/10.3390/plants13081130
Chicago/Turabian StyleLiu, Xinyong, Zixuan Yu, Xiaohong Tong, Longxue Chang, Jie Huang, Yifeng Wang, Jiezheng Ying, Xingwang Li, Shen Ni, and Jian Zhang. 2024. "Fine Mapping of qAL5.2 Controlling Anther Length in Oryza sativa" Plants 13, no. 8: 1130. https://doi.org/10.3390/plants13081130
APA StyleLiu, X., Yu, Z., Tong, X., Chang, L., Huang, J., Wang, Y., Ying, J., Li, X., Ni, S., & Zhang, J. (2024). Fine Mapping of qAL5.2 Controlling Anther Length in Oryza sativa. Plants, 13(8), 1130. https://doi.org/10.3390/plants13081130