
The Unique Role of the ECERIFERUM2-LIKE Clade of the BAHD Acyltransferase Superfamily in Cuticular Wax Metabolism

{kind=link}
{kind=link}
{kind=link}
{kind=link}
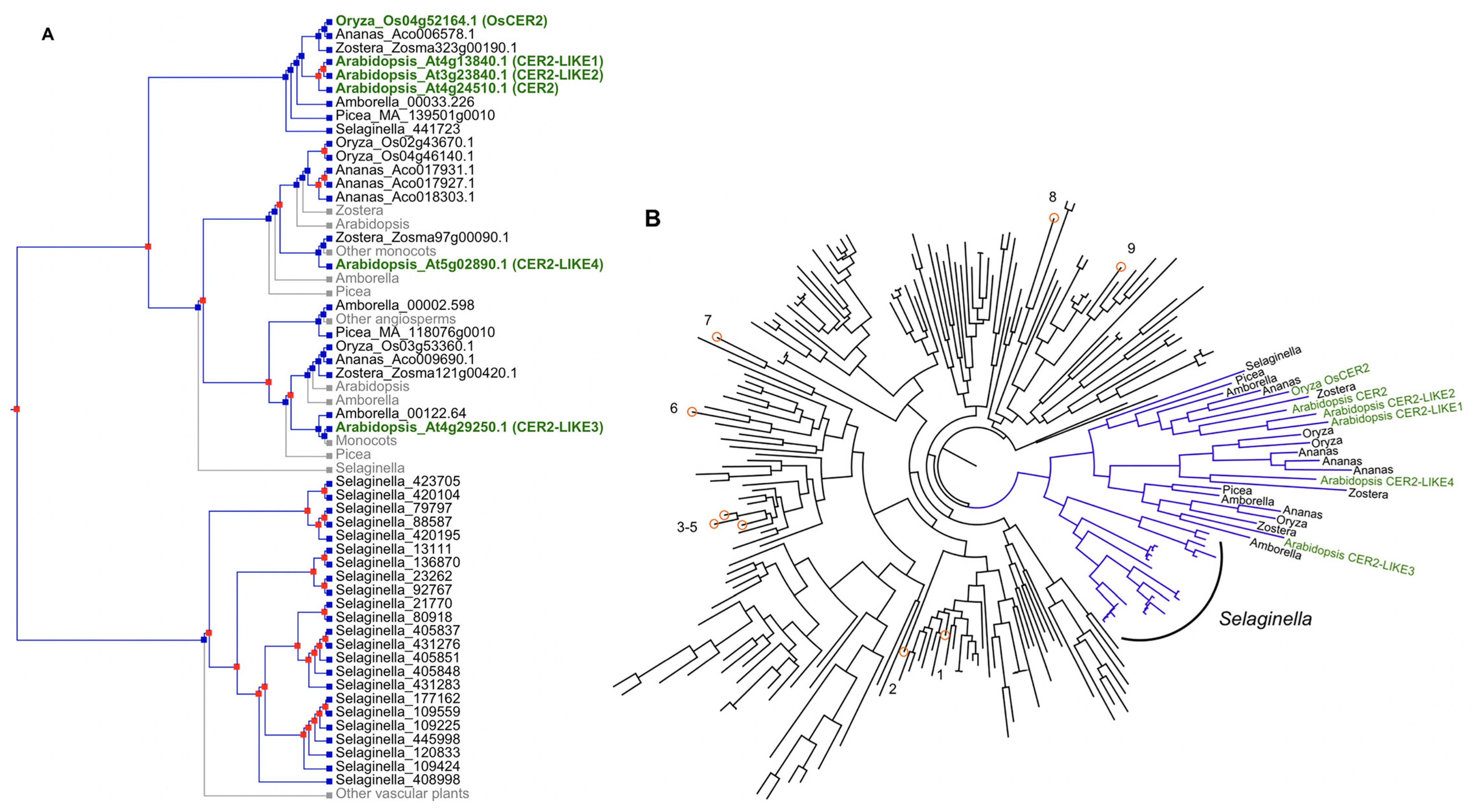{kind=link}
Abstract
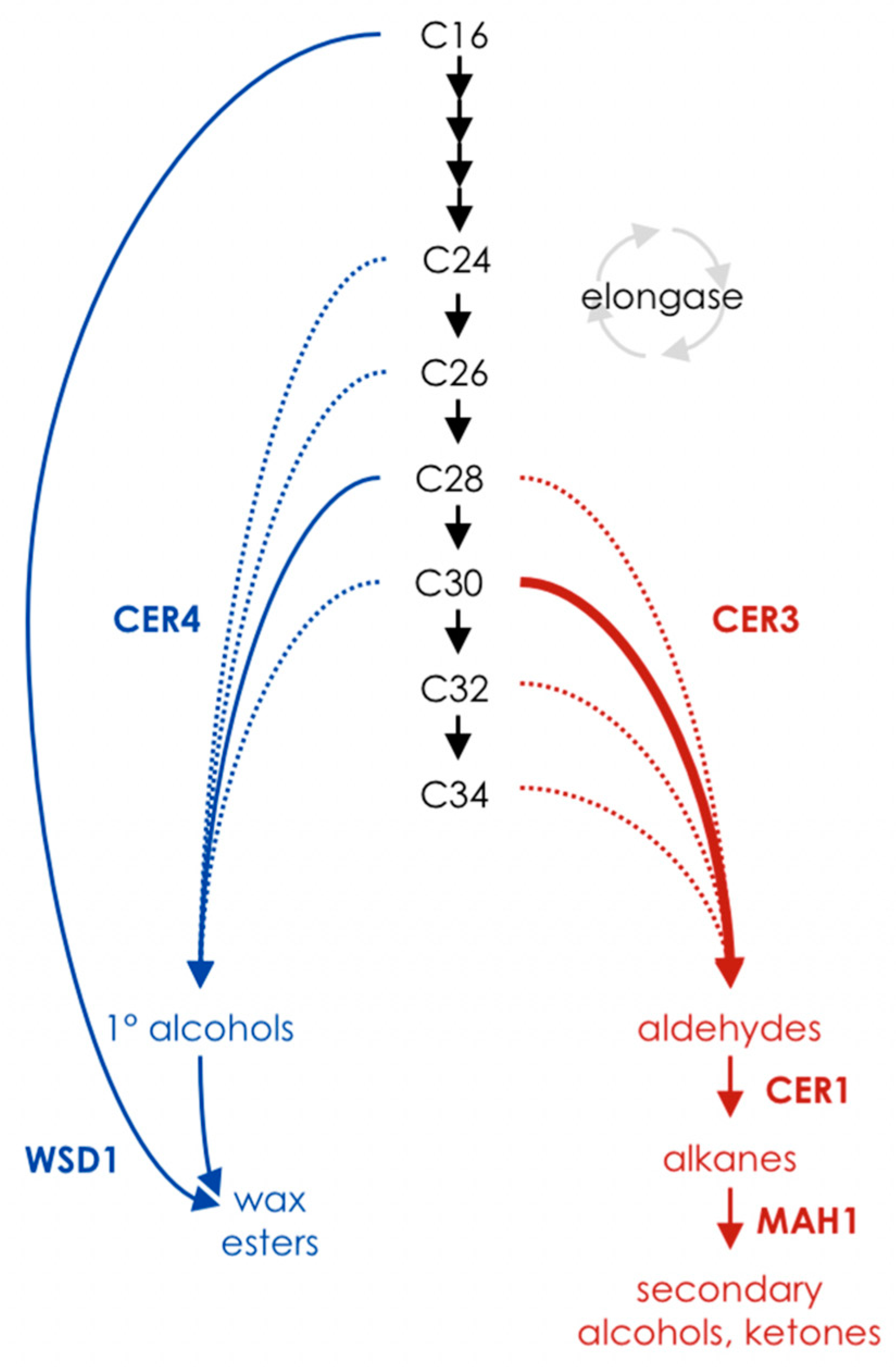
:1. Very-Long-Chain Fatty Acids and Cuticular Wax Metabolism
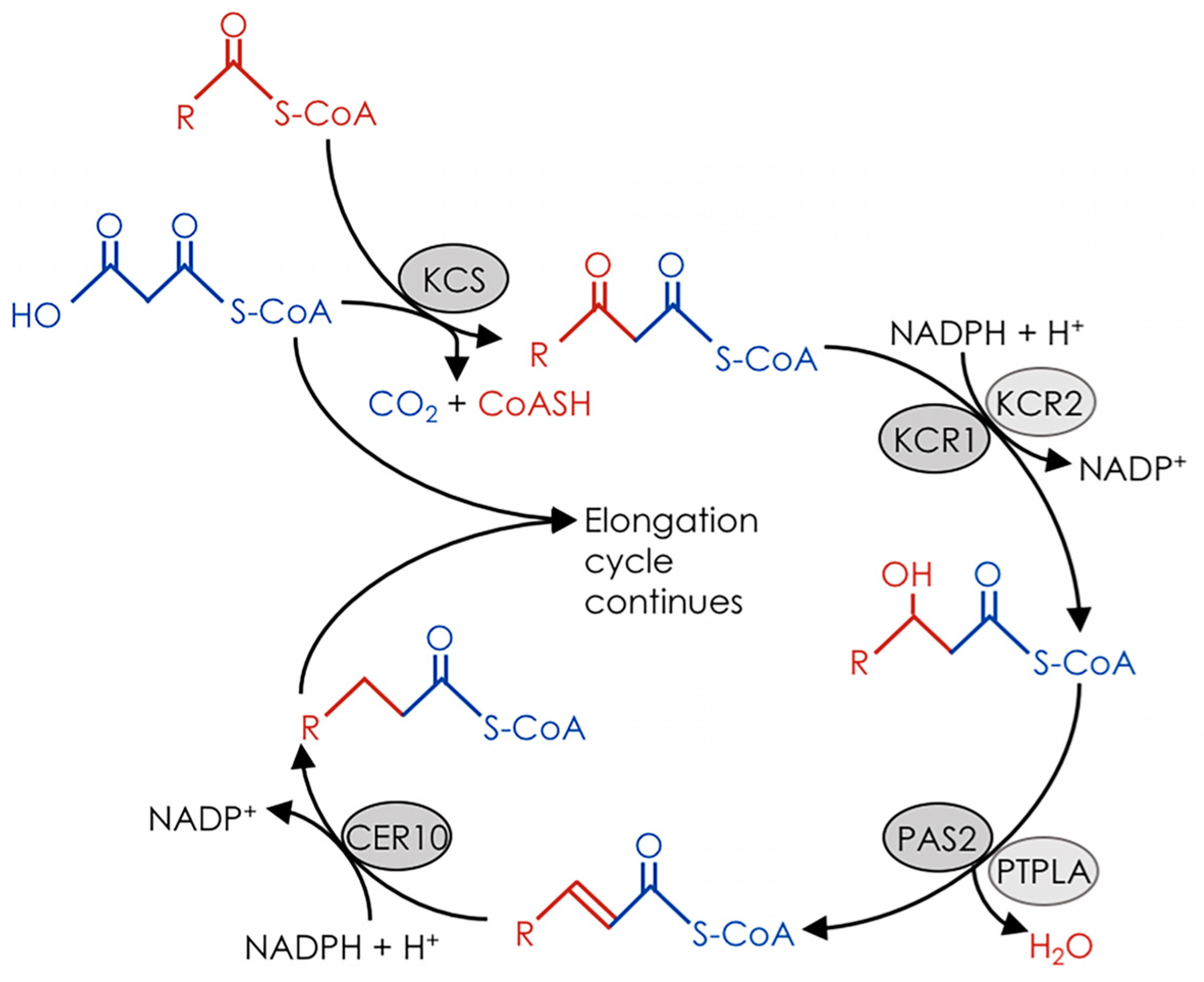
2. The Fatty Acid Elongase
3. Identification and Characterization of ECERIFERUM2-LIKE (CER2-LIKE) Genes
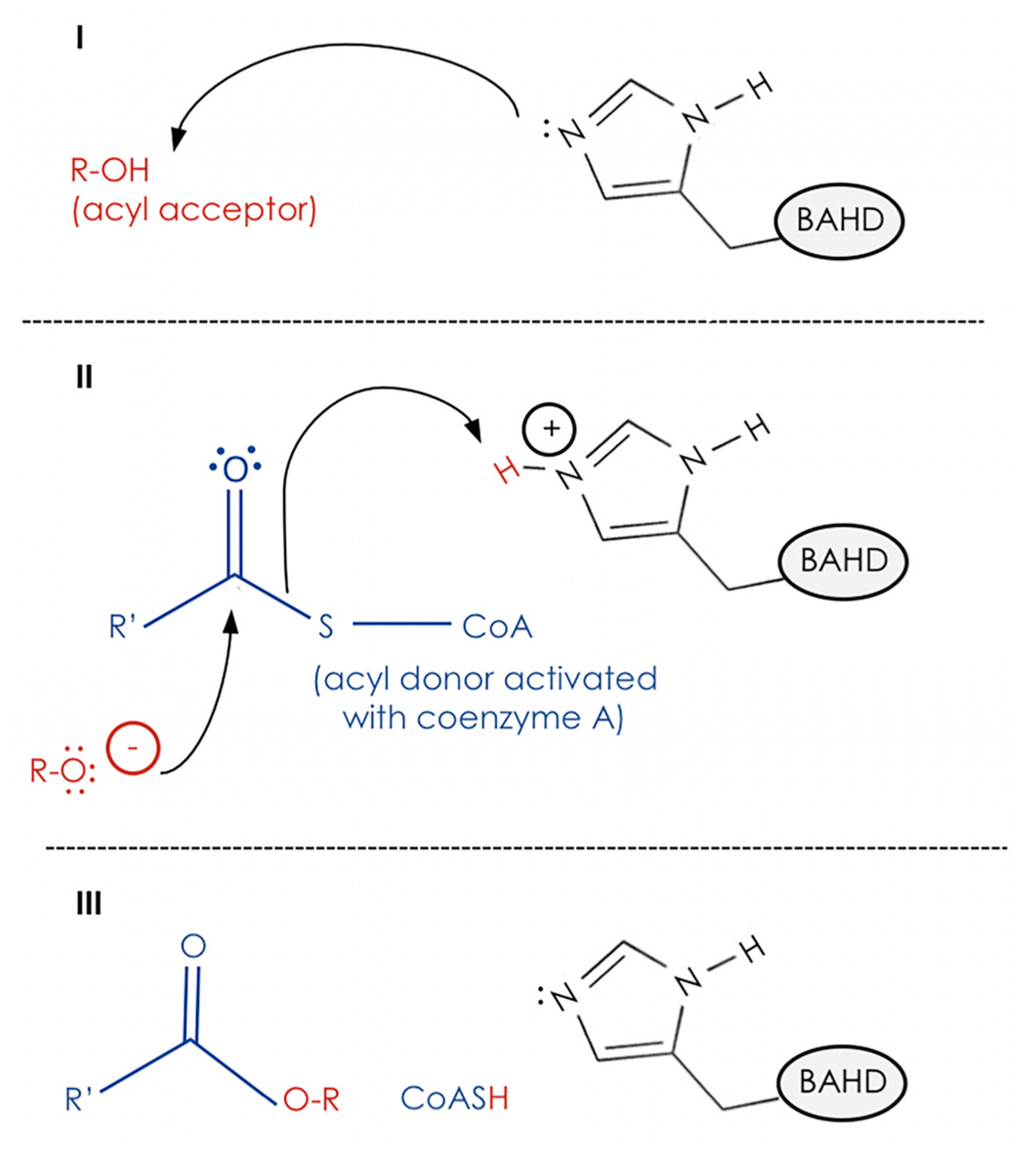
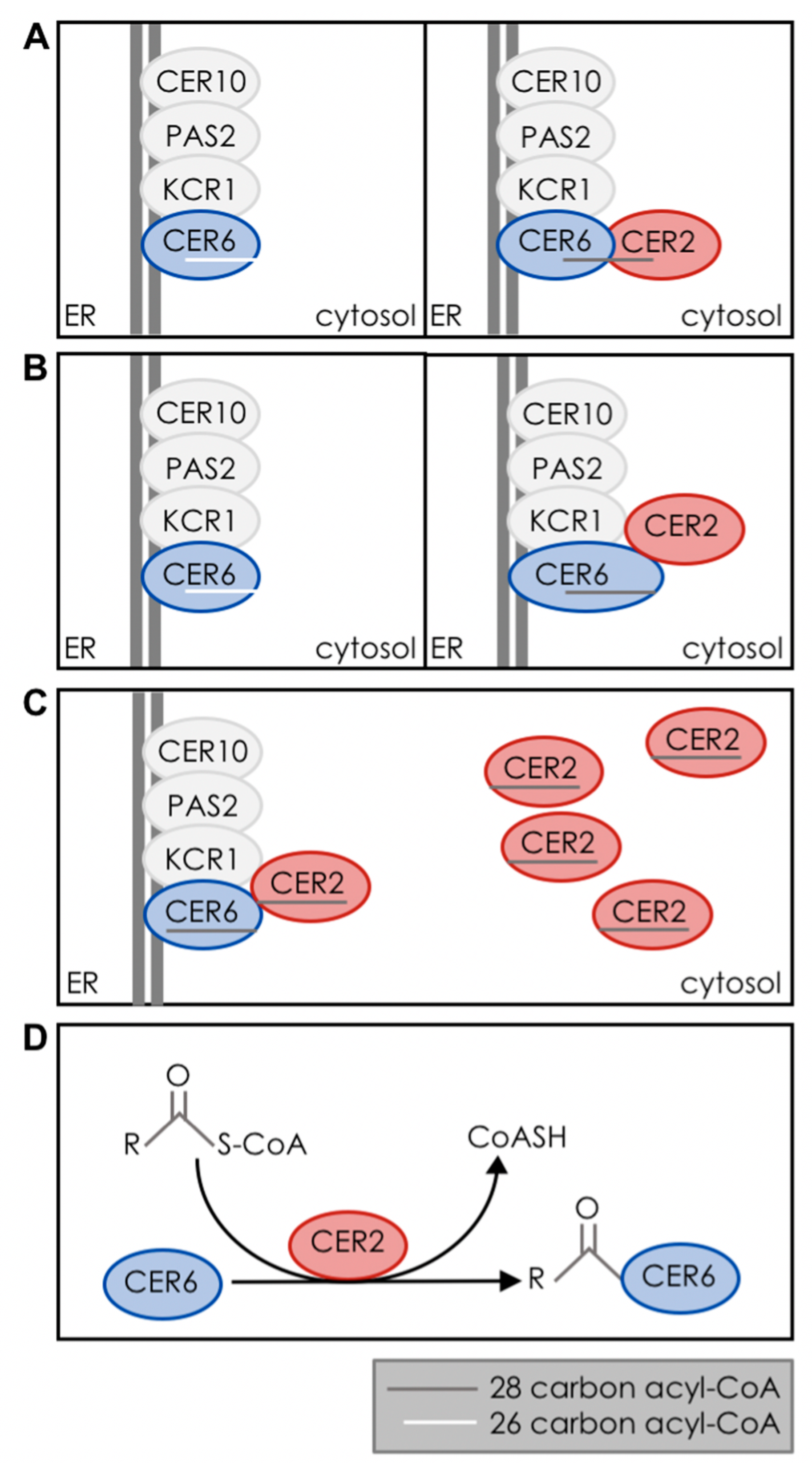
4. Biochemical Function of CER2
5. The CER2-LIKE Gene Family
6. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interests
References
- Fich, E.A.; Segerson, N.A.; Rose, J.K.C. The plant polyester cutin: Biosynthesis, structure, and biological roles. Annu. Rev. Plant Biol. 2016, 67, 207–233. [Google Scholar] [CrossRef] [PubMed]
- Yeats, T.H.; Rose, J.K.C. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Haslam, T.M.; Kunst, L. Extending the story of very-long-chain fatty acid elongation. Plant Sci. 2013, 210, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.A.; Kunst, L. Very-long-chain fatty acid biosynthesis is controlled through the expression and specificity of the condensing enzyme. Plant J. 1997, 12, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Bach, L.; Michaelson, L.V.; Haslam, R.; Bellec, Y.; Gissot, L.; Marion, J.; Da Costa, M.; Boutin, J.P.; Miquel, M.; Tellier, F.; et al. The very-long-chain hydroxy fatty acyl-CoA dehydratase PASTICCINO2 is essential and limiting for plant development. Proc. Natl. Acad. Sci. USA 2008, 105, 14727–14731. [Google Scholar] [CrossRef] [PubMed]
- Morineau, C.; Gissot, L.; Bellec, Y.; Hematy, K.; Tellier, F.; Renne, C.; Haslam, R.; Beaudoin, F.; Napier, J.; Faure, J.D. Dual fatty acid elongase complex interactions in Arabidopsis. PLoS ONE 2016, 11, e0160631. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin, F.; Wu, X.; Li, F.; Haslam, R.P.; Markham, J.E.; Zheng, H.; Napier, J.A.; Kunst, L. Functional characterization of the Arabidopsis β-ketoacyl-coenzyme A reductase candidates of the fatty acid elongase. Plant Physiol. 2009, 150, 1174–1191. [Google Scholar] [CrossRef] [PubMed]
- Joubès, J.; Raffaele, S.; Bourdenx, B.; Garcia, C.; Laroche-Traineau, J.; Moreau, P.; Domergue, F.; Lessire, R. The VLCFA elongase gene family in Arabidopsis thaliana: Phylogenetic analysis, 3D modelling and expression profiling. Plant Mol. Biol. 2008, 67, 547–566. [Google Scholar] [CrossRef] [PubMed]
- Zank, T.K.; Zähringer, U.; Beckmann, C.; Pohnert, G.; Boland, W.; Holtorf, H.; Reski, R.; Lerchl, J.; Heinz, E. Cloning and functional characterisation of an enzyme involved in the elongation of ∆6-polyunsaturated fatty acids from the moss Physcomitrella patens. Plant J. 2002, 31, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Eiamsa-Ard, P.; Kanjana-Opas, A.; Cahoon, E.B.; Chodok, P.; Kaewsuwan, S. Two novel Physcomitrella patens fatty acid elongases (ELOs): Identification and functional characterization. Appl. Microbiol. Biotech. 2013, 97, 3485–3497. [Google Scholar] [CrossRef] [PubMed]
- Trenkamp, S.; Martin, W.; Tietjen, K. Specific and differential inhibition of very-long-chain fatty acid elongases from Arabidopsis thaliana by different herbicides. Proc. Natl. Acad. Sci. USA 2004, 101, 11903–11908. [Google Scholar] [CrossRef] [PubMed]
- Blacklock, B.J.; Jaworski, J.G. Substrate specificity of Arabidopsis 3-ketoacyl-CoA synthases. Biochem. Biophys. Res. Commun. 2006, 346, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Gable, K.; Beaudoin, F.; Cahoon, E.; Jaworski, J.; Napier, J.A.; Dunn, T.M. Members of the Arabidopsis FAE1-like 3-ketoacyl-CoA synthase gene family substitute for the Elop proteins of Saccharomyces cerevisiae. J. Biol. Chem. 2006, 281, 9018–9029. [Google Scholar] [CrossRef] [PubMed]
- Tresch, S.; Heilmann, M.; Christiansen, N.; Looser, R.; Grossmann, K. Inhibition of saturated very-long-chain fatty acid biosynthesis by mefluidide and perfluidone, selective inhibitors of 3-ketoacyl-CoA synthases. Phytochemistry 2012, 76, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, M.; Hanhart, C.J.; Thiel, F. A genetic and phenotypic description of eceriferum (cer) mutants in Arabidopsis thaliana. J. Hered. 1989, 80, 118–122. [Google Scholar] [CrossRef]
- McNevin, J.P.; Woodward, W.; Hannoufa, A.; Feldmann, K.A.; Lemieux, B. Isolation and characterization of eceriferum (cer) mutants induced by T-DNA insertions in Arabidopsis thaliana. Genome 1993, 36, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Negruk, V.; Yang, P.; Subramanian, M.; McNevin, J.P.; Lemieux, B. Molecular cloning and characterization of the CER2 gene of Arabidopsis thaliana. Plant J. 1996, 9, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Nikolau, B.J.; Schnable, P.S. Cloning and characterization of CER2, an Arabidopsis gene that affects cuticular wax accumulation. Plant Cell 1996, 8, 1291–1304. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.A.; Clemens, S.; Zachgo, S.; Giblin, E.M.; Taylor, D.C.; Kunst, L. CUT1, an Arabidopsis gene required for cuticular wax biosynthesis and pollen fertility, encodes a very-long-chain fatty acid condensing enzyme. Plant Cell 1999, 11, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Haslam, T.M.; Manas-Fernandez, A.; Zhao, L.; Kunst, L. Arabidopsis ECERIFERUM2 is a component of the fatty acid elongation machinery required for fatty acid extension to exceptional lengths. Plant Physiol. 2012, 160, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, G.; Avato, P.; Salamini, F. Glossy mutants of maize. VI. Chemical constituents of glossy-2 epicuticular waxes. Maydica 1975, 20, 165–173. [Google Scholar]
- Tacke, E.; Korfhage, C.; Michel, D.; Maddaloni, M.; Motto, M.; Lanzini, S.; Salamini, F.; Doring, H.P. Transposon tagging of the maize Glossy2 locus with the transposable element En/Spm. Plant J. 1995, 8, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Pascal, S.; Bernard, A.; Sorel, M.; Pervent, M.; Vile, D.; Haslam, R.P.; Napier, J.A.; Lessire, R.; Domergue, F.; Joubès, J. The Arabidopsis cer26 mutant, like the cer2 mutant, is specifically affected in the very long chain fatty acid elongation process. Plant J. 2013, 73, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Haslam, T.M.; Haslam, R.; Thoraval, D.; Pascal, S.; Delude, C.; Domergue, F.; Fernández, A.M.; Beaudoin, F.; Napier, J.A.; Kunst, L.; et al. ECERIFERUM2-LIKE proteins have unique biochemical and physiological functions in very-long-chain fatty acid elongation. Plant Physiol. 2015, 167, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Haslam, T.M.; Kunst, L. Arabidopsis ECERIFERUM2-LIKEs are mediators of condensing enzyme function. Plant Physiol 2017. submitted for publication. [Google Scholar]
- Preuss, D.; Lemieux, B.; Yen, G.; Davis, R.W. A conditional sterile mutation eliminates surface components from Arabidopsis pollen and disrupts cell signaling during fertilization. Genes Dev. 1993, 7, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Hegebarth, D.; Buschhaus, C.; Joubès, J.; Thoraval, D.; Bird, D.; Jetter, R. Arabidopsis ketoacyl-CoA synthase 16 forms C36/C38 acyl precursors for leaf trichome and pavement surface wax. Plant. Cell Environ. 2007, in press. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Guan, Y.; Zhang, D.; Dong, X.; Tian, L.; Qu, L.Q. A β-ketoacyl-CoA synthase is involved in rice leaf cuticular wax synthesis and requires a CER2-LIKE protein as a cofactor. Plant Physiol. 2017, 173, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zeisler, V.; Schreiber, L.; Gao, J.; Hu, K.; Wen, J.; Yi, B. Overexpression of the novel Arabidopsis gene At5g02890 alters inflorescence stem wax composition and affects phytohormone homeostasis. Front. Plant Sci. 2017, 8, 68. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, J.C. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant Biol. 2006, 9, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Panikashvili, D.; Shi, J.X.; Schreiber, L.; Aharoni, A. The Arabidopsis DCR encoding a soluble BAHD acyltransferase is required for cutin polyester formation and seed hydration properties. Plant Physiol. 2009, 151, 1773–1789. [Google Scholar] [CrossRef] [PubMed]
- Rautengarten, C.; Ebert, B.; Ouellet, M.; Nafisi, M.; Baidoo, E.E.K.; Benke, P.; Stranne, M.; Mukhopadhyay, A.; Keasling, J.D.; Sakuragi, Y.; et al. Arabidopsis deficient in cutin ferulate encodes a transferase required for feruloylation of ω-hydroxy fatty acids in cutin polyester. Plant Physiol. 2012, 158, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Nakayama, T.; Nishino, T. Proposed mechanism and functional amino acid residues of malonyl-CoA: anthocyanin 5-O-glucoside-6’’’-O-malonyltransferase from flowers of Salvia splendens, a member of the versatile plant acyltransferase family. Biochemistry 2003, 42, 1764–1771. [Google Scholar] [CrossRef] [PubMed]
- Bayer, A.; Ma, X.; Stöckigt, J. Acetyltransfer in natural product biosynthesis—Functional cloning and molecular analysis of vinorine synthase. Bioorg. Med. Chem. 2004, 12, 2787–2795. [Google Scholar] [CrossRef] [PubMed]
- Lallemand, L.A.; Zubieta, C.; Lee, S.G.; Wang, Y.; Acajjaoui, S.; Timmins, J.; McSweeney, S.; Jez, J.M.; McCarthy, J.G.; McCarthy, A.A. A structural basis for the biosynthesis of the major chlorogenic acids found in coffee. Plant Physiol. 2012, 160, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.M.; Hayes, R.P.; Youn, B.; Vermerris, W.; Sattler, S.E.; Kang, C. Elucidation of the structure and reaction mechanism of sorghum hydroxycinnamoyltransferase and its structural relationship to other coenzyme A-dependent transferases and synthases. Plant Physiol. 2013, 162, 640–651. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Koepke, J.; Panjikar, S.; Fritzsch, G.; Stöckigt, J. Crystal structure of vinorine synthase, the first representative of the BAHD superfamily. J. Biol. Chem. 2005, 280, 13576–13583. [Google Scholar] [CrossRef] [PubMed]
- Unno, H.; Ichimaida, F.; Suzuki, H.; Takahashi, S.; Tanaka, Y.; Saito, A.; Nishino, T.; Kusunoki, M.; Nakayama, T. Structural and mutational studies of anthocyanin malonyltransferases establish the features of BAHD enzyme catalysis. J. Biol. Chem. 2007, 282, 15812–15822. [Google Scholar] [CrossRef] [PubMed]
- Morales-Quintana, L.; Nuñez-Tobar, M.X.; Moya-León, M.A.; Herrera, R. Molecular dynamics simulation and site-directed mutagenesis of alcohol acyltransferase: A proposed mechanism of catalysis. J. Chem. Inf. Model. 2013, 53, 2689–2700. [Google Scholar] [CrossRef] [PubMed]
- Blacklock, B.J.; Jaworski, J.G. Studies into factors contributing to substrate specificity of membrane-bound 3-ketoacyl-CoA synthases. Eur. J. Biochem. 2002, 269, 4789–4798. [Google Scholar] [CrossRef] [PubMed]
- Jasinski, S.; Lecureuil, A.; Miquel, M.; Loudet, O.; Raffaele, S.; Froissard, M.; Guerche, P. Natural variation in seed very long chain fatty acid content is controlled by a new isoform of KCS18 in Arabidopsis thaliana. PLoS ONE 2012, 7, e49261. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C.A.; Hubbell, T.; Behnke, W.D.; Schroeder, F. Fatty acid binding protein: Stimulation of microsomal phosphatidic acid formation. Arch. Biochem. Biophys. 1997, 341, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Gossett, R.E.; Frolov, A.A.; Roths, J.B.; Behnke, W.D.; Kier, A.B.; Schroeder, F. Acyl-CoA binding proteins: Multiplicity and function. Lipids 1996, 31, 895–918. [Google Scholar] [CrossRef] [PubMed]
- Boylan, J.G.; Hamilton, J.A. Interactions of acyl-coenzyme A with phosphatidylcholine bilayers and serum albumin. Biochemistry 1992, 31, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Suh, M.C.; Samuels, A.L.; Jetter, R.; Kunst, L.; Pollard, M.; Ohlrogge, J.; Beisson, F. Cuticular lipid composition, surface structure, and gene expression in Arabidopsis stem epidermis. Plant Physiol. 2005, 139, 1649–1665. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Chye, M.L. New roles for acyl-CoA-binding proteins (ACBPs) in plant development, stress responses and lipid metabolism. Prog. Lipid Res. 2011, 50, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Xiao, S.; Kim, J.; Lung, S.; Chen, L.; Tanner, J.A.; Suh, M.C.; Chye, M.L. Arabidopsis membrane-associated acyl-CoA-binding protein ACBP1 is involved in stem cuticle formation. J. Exp. Bot. 2014, 65, 5473–5483. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, B.; Luca, V.D. Evolution of acyltransferase genes: Origin and diversification of the BAHD superfamily of acyltransferase involved in secondary metabolism. Evol. Metab. Pathw. 2000, 285–315. [Google Scholar] [CrossRef]
- Nancolas, B.; Bull, I.; Stenner, R.; Dufour, V.; Curnow, P. Saccharomyces cerevisiae Atf1p is an alcohol acetyltransferase and a thioesterase in vitro. Yeast 2017, 34, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Bartley, L.E.; Peck, M.L.; Kim, S.R.; Ebert, B.; Maniseri, C.; Chiniquy, D.; Sykes, R.; Gao, L.; Rautengarten, C.; Vega-Sanchez, M.E.; et al. Overexpression of a BAHD acyltransferase, OsAt10, alters rice cell wall hydroxycinnamic acid content and saccharification. Plant Physiol. 2013, 161, 1615–1633. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.H.; Gou, J.Y.; Liu, C.J. BAHD superfamily of acyl-CoA dependent acyltransferases in Populus and Arabidopsis: Bioinformatics and gene expression. Plant Mol. Biol. 2009, 70, 421–442. [Google Scholar] [CrossRef] [PubMed]
- Tuominen, L.K.; Johnson, V.E.; Tsai, C.J. Differential phylogenetic expansions in BAHD acyltransferases across five angiosperm taxa and evidence of divergent expression among Populus paralogues. BMC Genom. 2011, 12, 236. [Google Scholar] [CrossRef] [PubMed]
- Busta, L.; Budke, J.M.; Jetter, R. The moss Funaria hygrometrica has cuticular wax similar to vascular plants, with distinct composition on leafy gametophyte, calyptra and sporophyte capsule surfaces. Ann. Bot. 2016, 118, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Kondo, S.; Hori, K.; Sasaki-Sekimoto, Y.; Kobayashi, A.; Kato, T.; Yuno-Ohta, N.; Nobusawa, T.; Ohtaka, K.; Shimojima, M.; Ohta, H. Primitive extracellular lipid components on the surface of the charophytic alga Klebsormidium flaccidum and their possible biosynthetic pathways as deduced from the genome sequence. Front. Plant Sci. 2016, 7, 952. [Google Scholar] [CrossRef] [PubMed]
- Jetter, R.; Riederer, M. Composition of cuticular waxes on Osmunda regalis fronds. J. Chem. Ecol. 2000, 26, 399–412. [Google Scholar] [CrossRef]
- Baker, E.A.; Gaskin, R.E. Composition of leaf epicuticular waxes of Pteridium sub-species. Phytochemistry 1987, 26, 2847–2848. [Google Scholar] [CrossRef]
- Buda, G.J.; Barnes, W.J.; Fich, E.A.; Park, S.; Yeats, T.H.; Zhao, L.; Domozych, D.S.; Rose, J.K.C. An ATP binding cassette transporter is required for cuticular wax deposition and desiccation tolerance in the moss Physcomitrella patens. Plant Cell 2013, 25, 4000–4013. [Google Scholar] [CrossRef] [PubMed]
- Baas, M.; Pancost, R.; van Geel, B.; Sinninghe Damsté, J.S. A comparative study of lipids in Sphagnum species. Org. Geochem. 2000, 31, 535–541. [Google Scholar] [CrossRef]
- Haas, K. Surface wax of Andreaea and Pogonatum species. Phytochemistry 1982, 21, 657–659. [Google Scholar] [CrossRef]
- Neinhuis, C.; Jetter, R. Ultrastructure and chemistry of epicuticular wax crystals in Polytrichales sporophytes. J. Bryol. 1995, 18, 399–406. [Google Scholar] [CrossRef]
- Richardson, A.; Wojciechowski, T.; Franke, R.; Schreiber, L.; Kerstiens, G.; Jarvis, M.; Fricke, W. Cuticular permeance in relation to wax and cutin development along the growing barley (Hordeum vulgare) leaf. Planta 2007, 225, 1471–1481. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, L.; Bessau, S.; Geoffroy, P.; Ritzenthaler, C.; Meyer, D.; Lapierre, C.; Pollet, B.; Legrand, M. Silencing of hydroxycinnamoyl-coenzyme A shikimate/quinate hydroxycinnamoyltransferase affects phenylpropanoid biosynthesis. Plant Cell 2004, 16, 1446–1465. [Google Scholar] [CrossRef] [PubMed]
- Grienenberger, E.; Besseau, S.; Geoffroy, P.; Debayle, D.; Heintz, D.; Lapierre, C.; Pollet, B.; Heitz, T.; Legrand, M. A BAHD acyltransferase is expressed in the tapetum of Arabidopsis anthers and is involved in the synthesis of hydroxycinnamoyl spermidines. Plant J. 2009, 58, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Molina, I.; Li-Beisson, Y.; Beisson, F.; Ohlrogge, J.B.; Pollard, M. Identification of an Arabidopsis feruloyl-coenzyme A transferase required for suberin synthesis. Plant Physiol. 2009, 151, 1317–1328. [Google Scholar] [CrossRef] [PubMed]
- Kosma, D.K.; Molina, I.; Ohlrogge, J.B.; Pollard, M. Identification of an Arabidopsis fatty alcohol: caffeoyl-coenzyme A acyltransferase required for the synthesis of alkyl hydroxycinnamates in root waxes. Plant Physiol. 2012, 160, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Leshem, Y.; Johnson, C.; Wuest, S.E.; Song, X.; Ngo, Q.A.; Grossniklaus, U.; Sundaresan, V. Molecular characterization of the glauce mutant: A central cell-specific function is required for double fertilization in Arabidopsis. Plant Cell 2012, 24, 3264–3277. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Wang, H.; Fujioka, S.; Zhou, T.; Tian, H.; Tian, W.; Wang, X. Homeostasis of brassinosteroids regulated by DRL1, a putative acyltransferase in Arabidopsis. Mol. Plant 2013, 6, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Cho, Y.H.; Kim, K.; Matsui, M.; Son, S.H.; Kim, S.K.; Fujioka, S.; Hwang, I. BAT1, a putative acyltransferase, modulates brassinosteroid levels in Arabidopsis. Plant J. 2013, 73, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.; Breuer, C.; Kawamura, A.; Jikumaru, Y.; Hanada, A.; Fujioka, S.; Ichikawa, T.; Kondou, Y.; Matsui, M.; Kamiya, Y.; et al. Arabidopsis PIZZA has the capacity to acylate brassinosteroids. PLoS ONE 2012, 7, e46805. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, G.; Ubukata, T.; Nozue, H.; Kobayashi, Y.; Takahi, M.; Yamamoto, H.; Hayashida, N. Malonylation is a key reaction in the metabolism of xenobiotic phenolic glucosides in Arabidopsis and tobacco. Plant J. 2010, 63, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Qualley, A.; Fan, B.; Dudareva, N.; Chen, Z. An important role of a BAHD acyl transferase-like protein in plant innate immunity. Plant J. 2009, 57, 1040–1053. [Google Scholar] [CrossRef] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haslam, T.M.; Gerelle, W.K.; Graham, S.W.; Kunst, L. The Unique Role of the ECERIFERUM2-LIKE Clade of the BAHD Acyltransferase Superfamily in Cuticular Wax Metabolism. Plants 2017, 6, 23. https://doi.org/10.3390/plants6020023
Haslam TM, Gerelle WK, Graham SW, Kunst L. The Unique Role of the ECERIFERUM2-LIKE Clade of the BAHD Acyltransferase Superfamily in Cuticular Wax Metabolism. Plants. 2017; 6(2):23. https://doi.org/10.3390/plants6020023
Chicago/Turabian StyleHaslam, Tegan M., Wesley K. Gerelle, Sean W. Graham, and Ljerka Kunst. 2017. "The Unique Role of the ECERIFERUM2-LIKE Clade of the BAHD Acyltransferase Superfamily in Cuticular Wax Metabolism" Plants 6, no. 2: 23. https://doi.org/10.3390/plants6020023
APA StyleHaslam, T. M., Gerelle, W. K., Graham, S. W., & Kunst, L. (2017). The Unique Role of the ECERIFERUM2-LIKE Clade of the BAHD Acyltransferase Superfamily in Cuticular Wax Metabolism. Plants, 6(2), 23. https://doi.org/10.3390/plants6020023