
Variation in Yield Responses to Elevated CO2 and a Brief High Temperature Treatment in Quinoa

Abstract
:1. Introduction
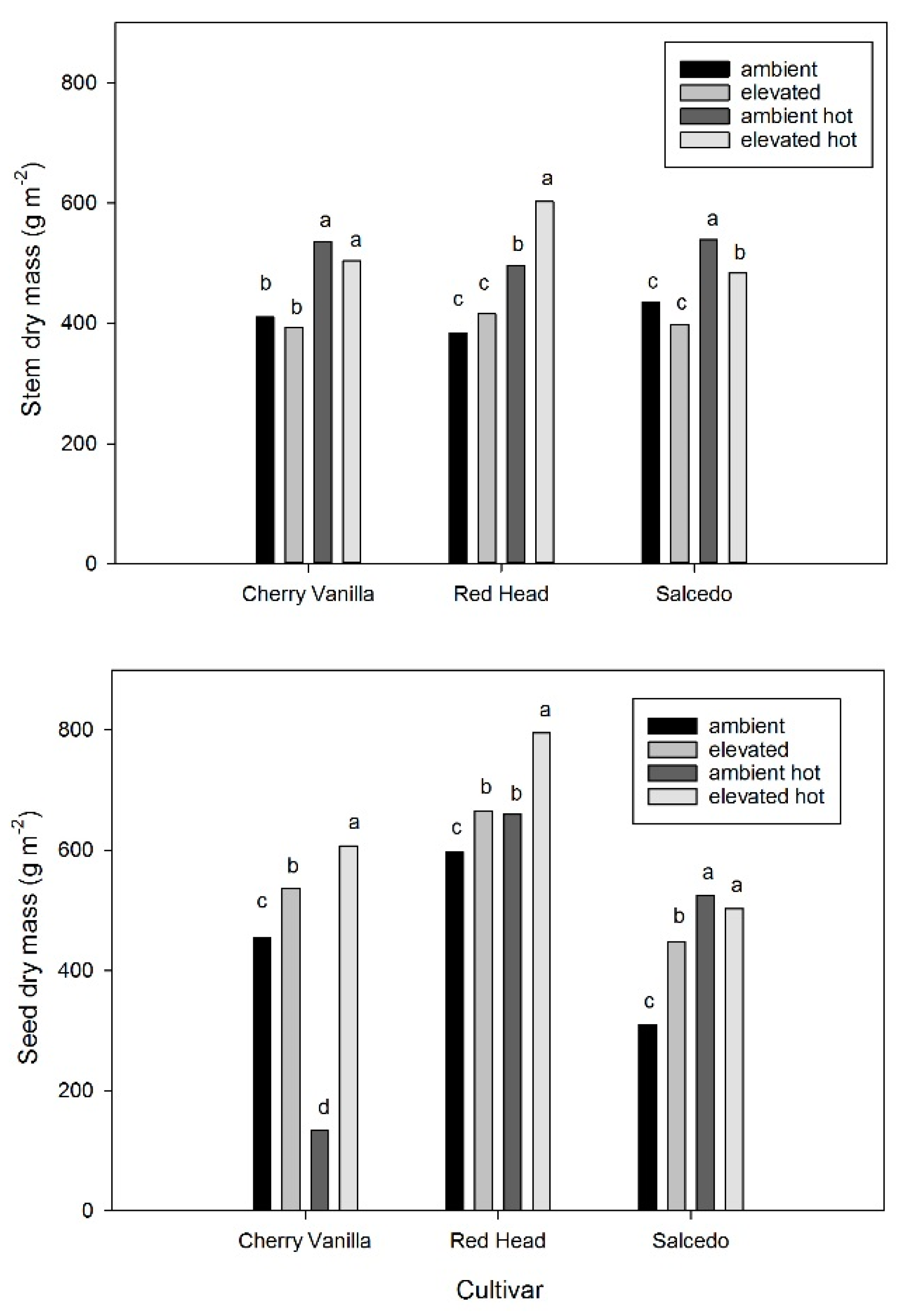
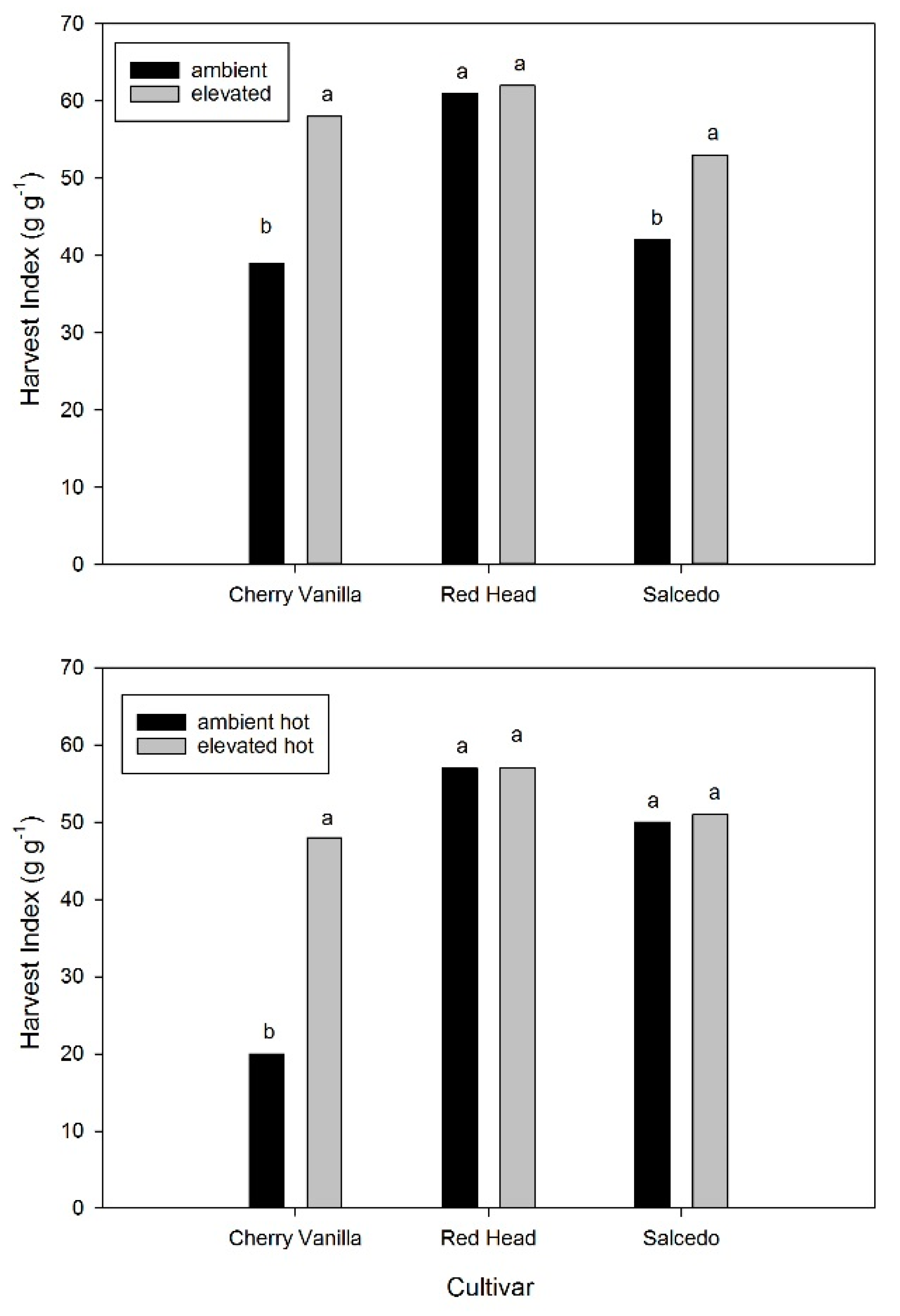
2. Results
3. Discussion
4. Materials and Methods
Acknowledgments
Conflicts of Interest
References
- Clausen, S.K.; Frenck, G.; Linden, L.G.; Mikkelsen, T.N.; Lunde, C.; Jorgensen, R.B. Effects of single and multifactor treatments with elevated temperature, CO2 and ozone on oilseed rape and barley. J. Agron. Crop Sci. 2011, 197, 442–453. [Google Scholar] [CrossRef]
- Bunce, J.A. Contrasting responses of seed yield to elevated carbon dioxide under field conditions within Ph. Vulg. Agric. Ecosyst. Environ. 2008, 128, 219–234. [Google Scholar] [CrossRef]
- Ahmed, F.E.; Hall, A.E.; Madore, M.A. Interactive effects of high temperature and elevated carbon dioxide concentration on cowpea [(Vigna unguiculata (L.) Walp.]. Plant Cell Environ. 1993, 16, 835–842. [Google Scholar] [CrossRef]
- Bishop, K.A.; Betzelberger, A.M.; Long, S.P.; Ainsworth, E.A. Is there potential to adapt soybean (Glycine max Merr.) to future CO2? An analysis of the yield response of 18 genotypes in free-air CO2 enrichment. Plant Cell Environ. 2015, 38, 1765–1774. [Google Scholar] [CrossRef] [PubMed]
- Bunce, J.A. Variable responses to CO2 of the duration of vegetative growth and yield within a maturity group in soybeans. Am. J. Plant Sci. 2016, 7, 1759–1764. [Google Scholar] [CrossRef]
- Batts, G.R.; Ellis, R.H.; Morison, J.I.L.; Nkemka, P.N.; Gregory, P.J.; Hadley, P. Yield and partitioning in crops of contrasting cultivars of winter wheat in response to CO2 and temperature in field studies using temperature gradient tunnels. J. Agric. Sci. 1998, 130, 17–27. [Google Scholar] [CrossRef]
- Bunce, J.A. Using FACE systems to screen wheat cultivars for yield increases at elevated CO2. Agronomy 2017, 7, 20. [Google Scholar] [CrossRef]
- Johannessen, M.M.; Mikkelsen, T.N.; Nersting, L.G.; Gullord, M.; von Bothmer, R.; Jorgenses, R.B. Effect of increased atmospheric CO2 on varieties of oat. Plant Breed. 2005, 124, 253–256. [Google Scholar] [CrossRef]
- Johannessen, M.M.; Mikkelsen, T.N.; Jorgensen, R.B. CO2 exploitation and genetic diversity in winter varieties of oilseed rape (Brassica napus); varieties of tomorrow. Euphytica 2002, 128, 75–86. [Google Scholar] [CrossRef]
- Hasegawa, T.; Tokida, T.; Nakamura, H.; Zhu, C.; Usui, Y.; Yoshimoto, M.; Fukuoka, M.; Fukuoka, M.; Wakatsuki, H.; Katayanagi, N.; et al. Rice cultivar responses to elevated CO2 at two free–air CO2 enrichment (FACE) site in Japan. Funct. Plant Biol. 2013, 40, 148–159. [Google Scholar] [CrossRef]
- Ziska, L.H.; Bunce, J.A.; Shimono, H.; Gealy, D.R.; Baker, J.T.; Newton, P.C.D.; Reynolds, M.P.; Jagadish, K.S.V.; Zhu, C.; Howden, M.; et al. Food Security and climate change: On the potential to adapt global crop production by active selection to rising atmospheric carbon dioxide. Proc. Biol. Sci. 2012, 279, 4097–4105. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, L.A.; Davey, P.A.; Bernacchi, C.J.; Dermody, O.C.; Heaton, E.A.; Moore, D.J.; Morgan, P.B.; Naidu, S.L.; Yoora, H.S.; Zhu, X.G.; et al. A meta-analysis of elevated [CO2] effect on soybean (Glycine max) physiology, growth and yield. Glob. Chang. Biol. 2002, 8, 695–709. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Xiong, Z.; Wolfe, D.; Zou, J. Response of rice production to elevated [CO2] and its interaction with rising temperature or nitrogen supply: A meta-analysis. Clim. Chang. 2015, 130, 529–543. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.Z.; Schjoerring, K.J. Effects of elevated atmospheric CO2 on physiology and yield of wheat (Triticum aestivum L.): A meta-analytic test of current hypotheses. Agric. Ecosyst. Environ. 2013, 178, 57–63. [Google Scholar] [CrossRef]
- Wheeler, T.; von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Schlenker, W.; Roberts, M.J. Nonlinear temperature effects indicate severe damages to U.S. crop yield under climate change. Proc. Natl. Acad. Sci. USA 2009, 106, 15594–15598. [Google Scholar] [CrossRef] [PubMed]
- Urban, D.; Robers, M.; Schlenker, W.; Lobell, D. Projected temperature changes indicate significant increase in interannual variability of U.S. maize yields. Clim. Chang. 2012, 112, 525–595. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Boote, K.J.; Kimball, B.A.; Ziska, L.H.; Izaurralde, R.C.; Ort, D.; Thomson, A.M.; Wolfe, D. Climate impacts on agriculture: Implication for crop production. Agron. J. 2011, 103, 351–370. [Google Scholar] [CrossRef]
- Lobell, D.B.; Field, C.B. Global scale climate-crop yield relationships and the impacts of recent warming. Environ. Res. Lett. 2007, 2, 014002. [Google Scholar] [CrossRef]
- Bishop, K.A.; Leakey, A.D.B.; Ainsworth, E.A. How seasonal temperature or water inputs affect the relative response of C3 crops to elevated [CO2]: A global analysis of open top chamber and free air CO2 enrichment studies. Food Energy Secur. 2014, 3, 33–45. [Google Scholar] [CrossRef]
- Broughton, K.J.; Smith, R.A.; Duursma, R.A.; Tan, D.K.Y.; Payton, P.; Bange, M.P.; Tissue, D.T. Warming alters the positive impact of elevated CO2 concentration on cotton growth and physiology during soil water deficit. Funct. Plant Biol. 2017, 44, 267–278. [Google Scholar] [CrossRef]
- Cai, C.; Yin, X.; He, S.; Jiang, W.; Si, C.; Struik, P.C.; Luo, W.; Li, G.; Xie, Y.; Xiong, Y.; et al. Responses of wheat and rice to factorial combinations of ambient and elevated CO2 and temperature in FACE experiments. Glob. Chang. Biol. 2015, 22, 856–874. [Google Scholar] [CrossRef] [PubMed]
- Dias de Oliveira, E.; Bramley, J.; Siddique, K.H.M.; Berger, H.S. Can elevated CO2 combined with high temperature ameliorate the effect of terminal drought in wheat? Funct. Plant Biol. 2013, 40, 160–171. [Google Scholar] [CrossRef]
- Kimball, B.A. Crop responses to elevated CO2 and interactions with H2O, N, and temperature. Curr. Opin. Plant Biol. 2016, 31, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Prasad, P.V.V.; Pisipati, S.R.; Mutava, R.N.; Tuinstra, M.R. Sensitivity of grain sorghum to high temperature stress during reproductive development. Crop Sci. 2008, 48, 1911–1917. [Google Scholar] [CrossRef]
- Ruiz-Vera, U.M.; Siebers, M.; Gray, S.B.; Drag, D.W.; Rosenthal, D.M.; Kimball, B.A.; Ort, D.R.; Bernacchi, C.J. Global warming can negate the expected CO2 stimulation in photosynthesis and productivity for soybean grown in the Midwestern United States. Plant Physiol. 2013, 162, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Vera, U.M.; Siebers, M.H.; Drag, D.W.; Ort, D.R.; Bernacchi, C.J. Canopy warming caused photosynthetic acclimation and reduced seed yield in maize grown at ambient and elevated [CO2]. Glob. Chang. Biol. 2015, 21, 4237–4249. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.R.; Bunce, J.A.; Tomecek, M.B.; Gealy, D.; McClung, A.; McCouch, S.; Ziska, L.H. Evidence for divergence of response in Indica, Japonica, and wild rice to high CO2 × temperature interaction. Glob. Chang. Biol. 2016, 22, 2620–2632. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Heckathorn, S.A.; Wang, X.; Philpott, S.M. A meta-analysis of plant physiological and growth responses to temperature and elevated CO2. Oecologia 2012, 169, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pulvento, C.; Riccardi, M.; Lavini, A.; Iafelice, G.; Marconi, E.; d’Andria, R. Yield and quality characteristics of quinoa grown in open field under different saline and non-saline irrigation regimes. J. Agron. Crop Sci. 2012, 198, 254–263. [Google Scholar] [CrossRef]
- Curti, R.N.; de la Vega, A.J.; Andrade, A.J.; Bramardi, S.J.; Bertero, H.D. Multi-environment evaluation for grain yield and its physiological determinants of quinoa genotypes across Northwest Argentina. Field Crops Res. 2014, 166, 46–57. [Google Scholar] [CrossRef]
- Curti, R.N.; de la Vega, A.J.; Andrade, A.J.; Bramardi, S.J.; Bertero, H.D. Adaptive responses of quinoa to diverse agro-ecological environments along an altitudinal gradient in North West Argentina. Field Crop. Res. 2016, 189, 10–18. [Google Scholar] [CrossRef]
- Gonzalez, J.A.; Bruno, M.; Valoy, M.; Prado, F.E. Genotypic variation of gas exchange parameters and leaf stable carbon and nitrogen isotopes in ten quinoa cultivars grown under drought. J. Agron. Crop Sci. 2011, 197, 81–93. [Google Scholar] [CrossRef]
- Lavini, A.; Pulvento, C.; d’Andria, R.; Riccardi, M.; Choukr-Allad, R.; Belhabib, O.; Yazar, A.; Incekaya, C.; Metin Sezen, S.; Qudir, M.; Jacobsen, S.E. Quinoa’s potential in the Mediterranean region. J. Agron. Crop Sci. 2014, 200, 344–360. [Google Scholar] [CrossRef]
- Lloyd, J.; Farquhar, G.D. Effects of rising temperature and CO2 on the physiology of tropical forest trees. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 1811–1817. [Google Scholar] [CrossRef] [PubMed]
- Patrick, J.W.; Colyvas, K. Crop yield components–photoassimilate supply- or utilization limted-organ development? Funct. Plant Biol. 2014, 41, 893–913. [Google Scholar] [CrossRef]
- Gomez, M.B.; Aguirre Castro, P.; Mignone, C.; Bertero, H.D. Can yield potential be increased by manipulation of reproductive partitioning in quinoa (Chenopodium quinoa)? Evidence from gibberellic acid synthesis inhibition using paclobrutrazol. Funct. Plant Biol. 2011, 38, 420–430. [Google Scholar] [CrossRef]
- Fitzgerald, G.J.; Tausz, M.; O’Leary, G.; Mollah, M.R.; Tausz-Pausch, S.; Seneweera, S.; Mock, I.; Low, M.; Partington, D.L.; McNeil, D.; et al. Elevated atmospheric [CO2] can dramatically increase wheat yields in semi-arid environments and buffer against heat waves. Glob. Chang. Biol. 2016, 22, 2269–2284. [Google Scholar] [CrossRef] [PubMed]
- Ferris, R.; Wheeler, T.R.; Ellis, R.H.; Hadley, P. Seed yield after environmental stress in soybean grown under elevated CO2. Crop Sci. 1999, 39, 710–718. [Google Scholar] [CrossRef]
- Shimono, H.; Bunce, J.A. Acclimation of nitrogen uptake capacity of rice to elevated atmospheric CO2 concentration. Ann. Bot. 2009, 103, 87–94. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Growth Stage | Cultivar | Ambient | Elevated | Ambient at Elevated |
|---|---|---|---|---|
| Before stress | Cherry Vanilla | 38.8a | 42.6b | 44.2b |
| Red Head | 39.5a | 42.5b | 45.0b | |
| Salcedo | 37.8a | 43.6b | 42.0b | |
| During stress | Cherry Vanilla | 32.7a | 39.8b | 40.8b |
| Stressed plants | Red Head | 34.1a | 41.6b | 41.7b |
| Salcedo | 32.2a | 37.8b | 38.9b | |
| After stress | Cherry Vanilla | 35.2a | 34.2a | 40.3b |
| Stressed plants | Red Head | 36.3a | 37.1a | 41.1b |
| Salcedo | 34.1a | 35.0a | 39.6b | |
| After stress | Cherry Vanilla | 34.7a | 35.4a | 40.8b |
| Control plants | Red Head | 35.8a | 36.2a | 41.0b |
| Salcedo | 35.1a | 35.8a | 39.3b |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bunce, J.A. Variation in Yield Responses to Elevated CO2 and a Brief High Temperature Treatment in Quinoa. Plants 2017, 6, 26. https://doi.org/10.3390/plants6030026
Bunce JA. Variation in Yield Responses to Elevated CO2 and a Brief High Temperature Treatment in Quinoa. Plants. 2017; 6(3):26. https://doi.org/10.3390/plants6030026
Chicago/Turabian StyleBunce, James A. 2017. "Variation in Yield Responses to Elevated CO2 and a Brief High Temperature Treatment in Quinoa" Plants 6, no. 3: 26. https://doi.org/10.3390/plants6030026
APA StyleBunce, J. A. (2017). Variation in Yield Responses to Elevated CO2 and a Brief High Temperature Treatment in Quinoa. Plants, 6(3), 26. https://doi.org/10.3390/plants6030026