Integrating Soil Silicon Amendment into Management Programs for Insect Pests of Drill-Seeded Rice



Abstract
:1. Introduction
2. Results
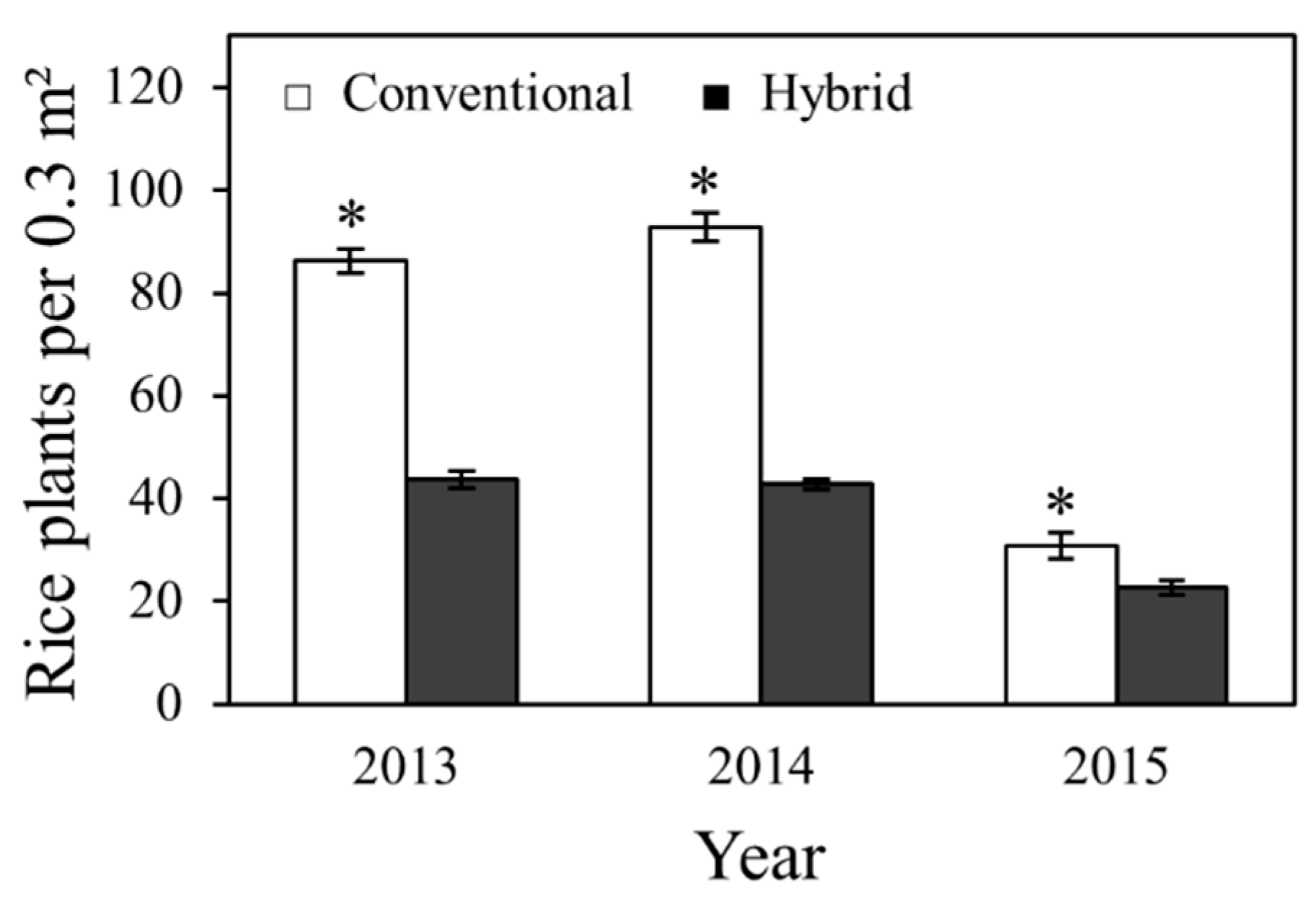
2.1. Effects on Plant Stand
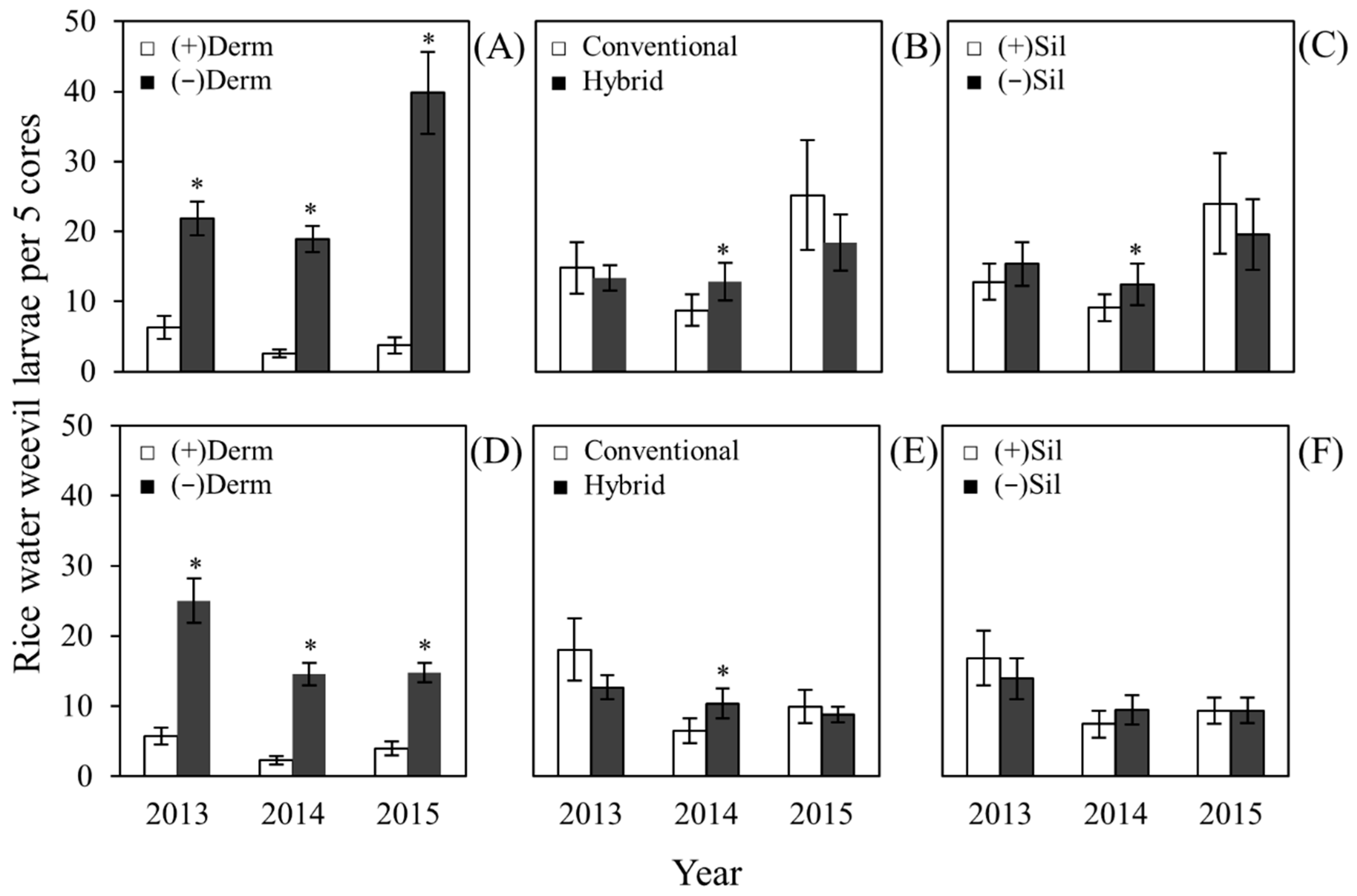
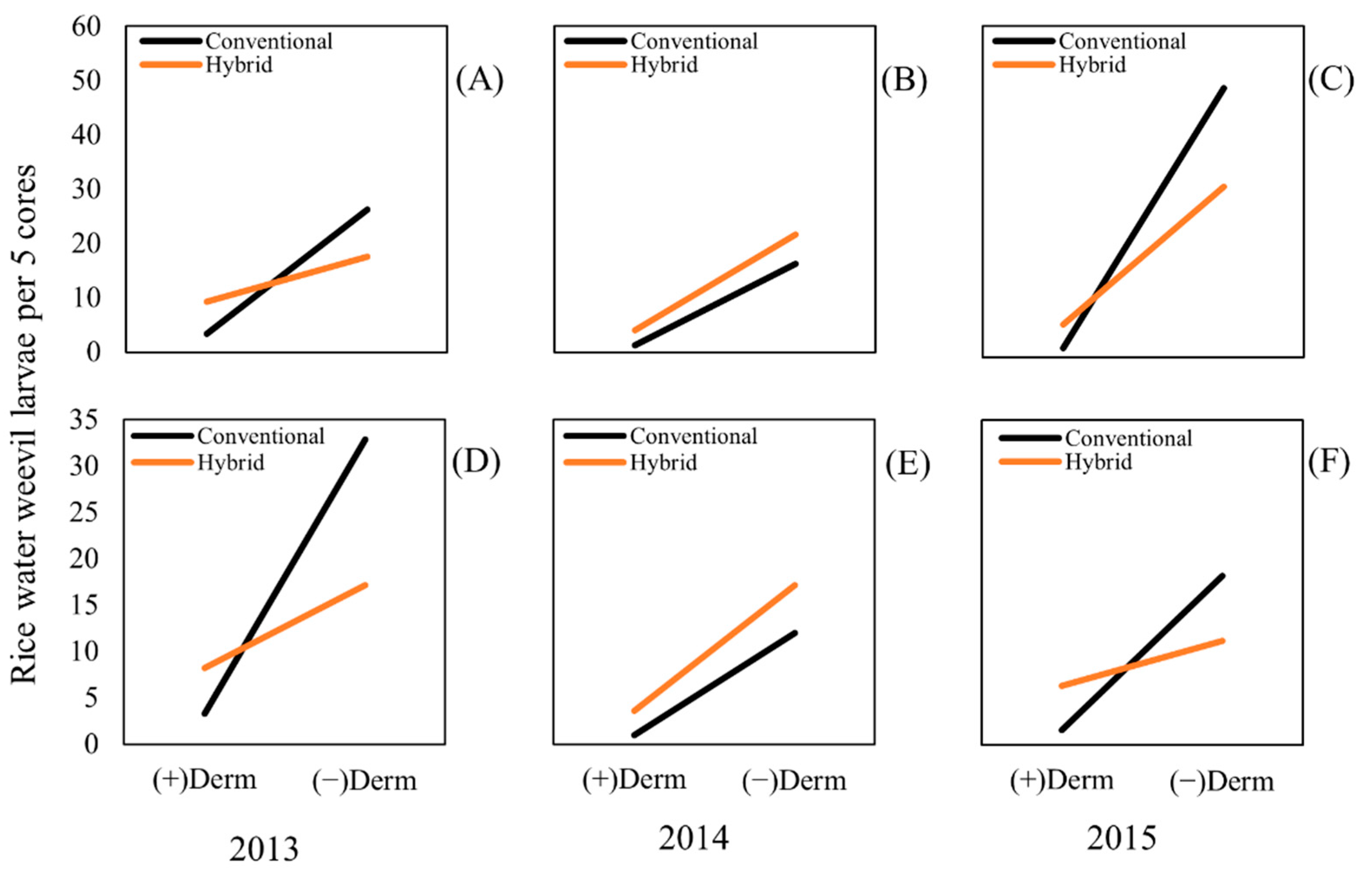
2.2. Effects on Rice Water Weevil Larval Density
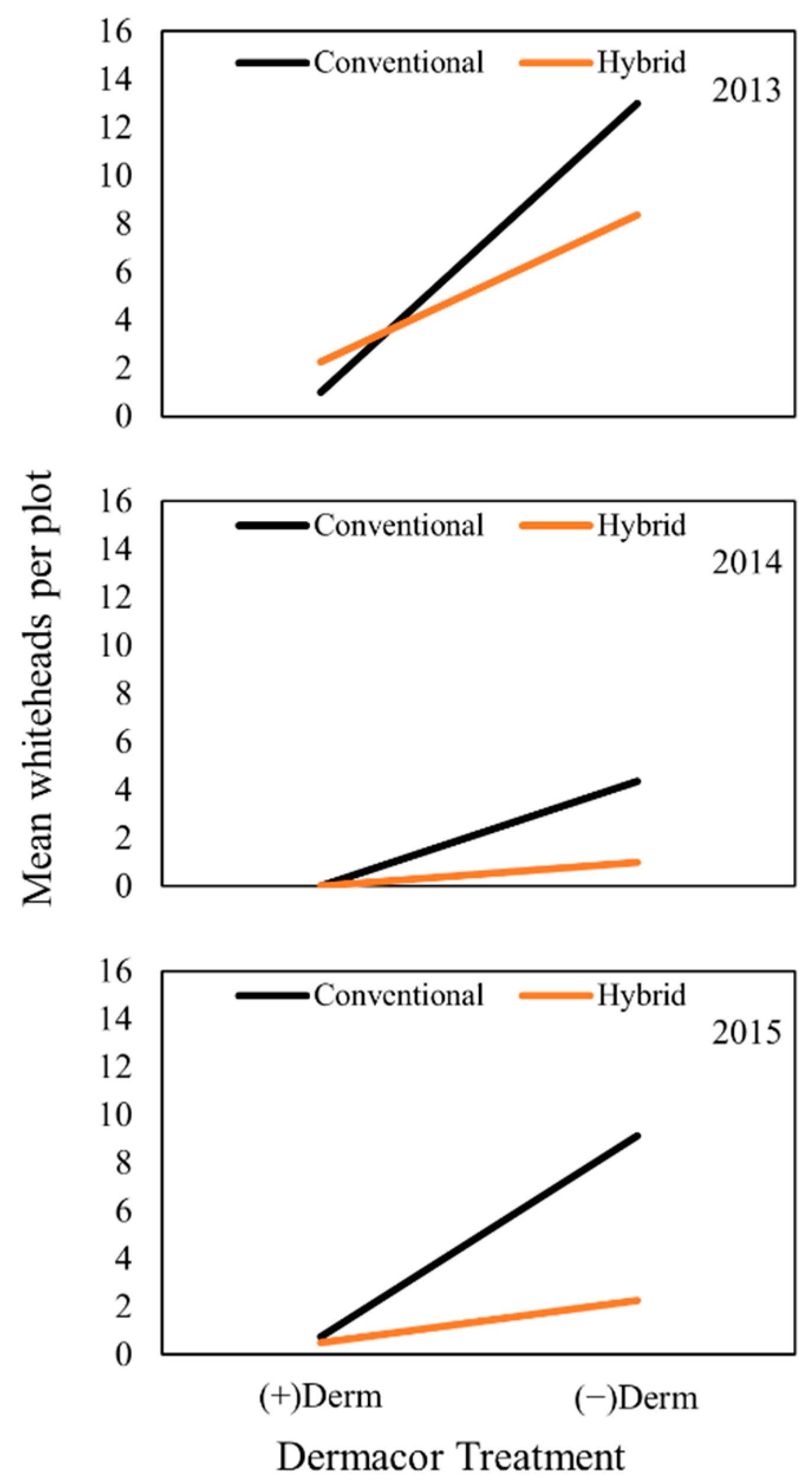
2.3. Effects on Whitehead Incidence
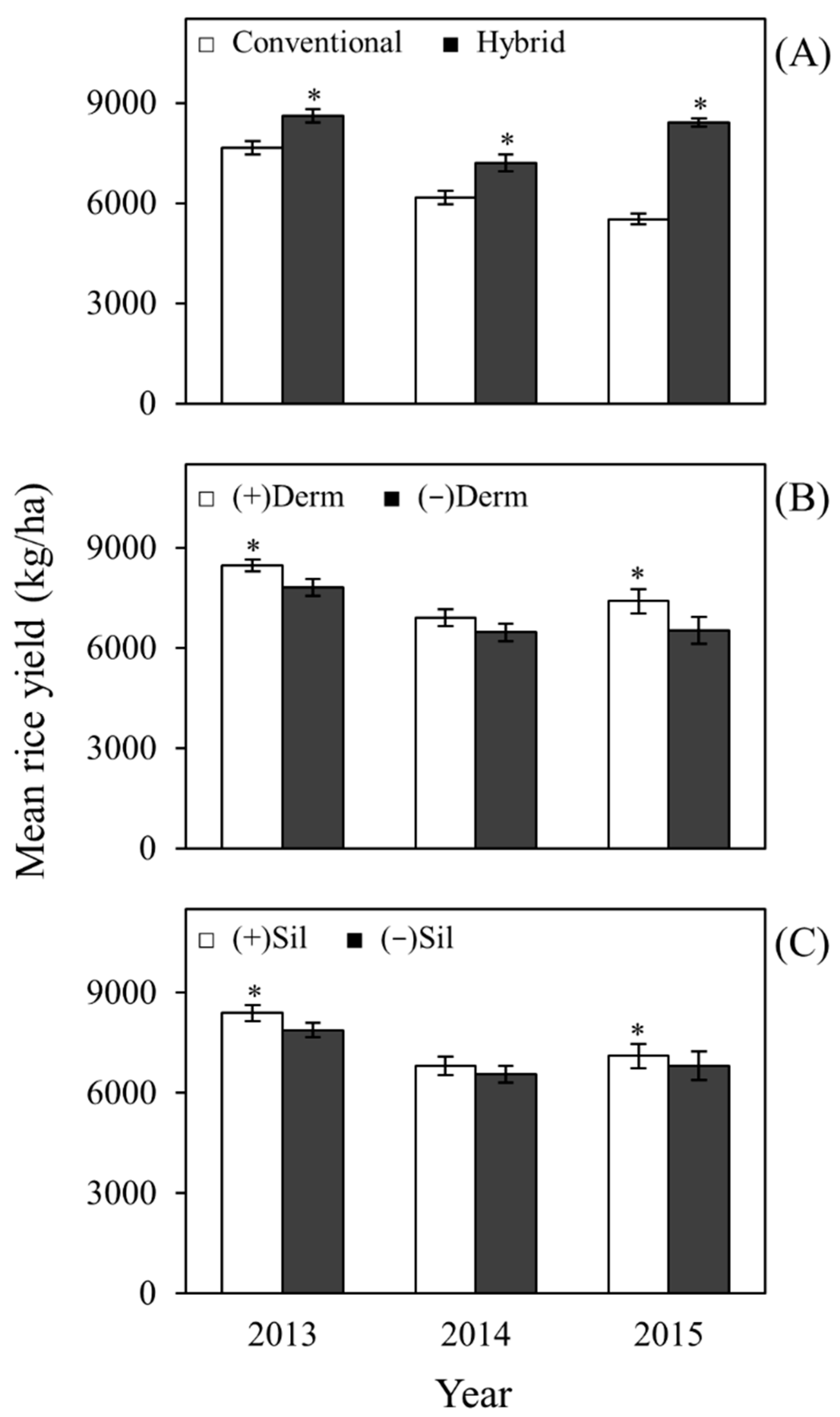
2.4. Effects on Yield
3. Discussion
4. Materials and Methods
Data Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A


References
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. 1999, 50, 641–664. [Google Scholar] [CrossRef] [PubMed]
- Sommer, M.; Kaczorek, D.; Kuzyahov, Y.; Breuer, J. Silicon Pools and Fluxes in Soils and Landscapes—A Review. J. Plant Nutr. Soil Sci. 2006, 169, 310–329. [Google Scholar] [CrossRef]
- Cornelius, J.T.; Delvauz, B.; Georg, R.B.; Lucas, Y.; Ranger, J.; Opfergelt, S. Tracing the Origin of Dissolved Silicon Transferred from Various Soil-Plant System towards River: A Review. Biogeosciences 2011, 8, 89–112. [Google Scholar] [CrossRef]
- Jones, L.H.P.; Handreck, K.A. Silica In Soils, Plants, and Animals. In Advances in Agronomy; Academic Press: Cambridge, MS, USA, 1967; pp. 107–149. [Google Scholar]
- Yoshida, S.; Ohnishi, Y.; Kitagishi, K. Chemical Forms, Mobility, and Deposition of Silicon in the Rice Plant. Soil Sci. Plant Nutr. 1962, 8, 107–111. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.; Christie, P. Mechanisms of Silicon Mediated Alleviation of Abiotic Stresses in Higher Plants: A Review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Takahashi, E. Soil, Fertilizer, and Plant Silicon Research in Japan; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Ma, J.F.; Goto, S.; Tamai, K.; Ichii, M. Role of Root Hairs and Lateral Roots in Silicon Uptake by Rice. Plant Physiol. 2001, 7, 1773–1780. [Google Scholar] [CrossRef]
- Katz, O. Beyond Grasses: The Potential Benefits of Studying Silicon Accumulation in Non-Grass Species. Front. Plant Sci. 2014. [Google Scholar] [CrossRef] [PubMed]
- International Plant Nutrition Center. Nutri Facts: Silicon. Available online: http://www.ipni.net/publication/nutrifacts-na.nsf/0/A7B4AB4D35C153BF85257ECE006E0E34/$FILE/NutriFacts-NA-14.pdf (accessed on 21 June 2017).
- Keeping, M.G.; Kvedaras, O.L.; Bruton, A.G. Epidermal Silicon in Sugarcane: Cultivar Differences and Role of Resistance to Sugarcane Stalk Borer Eldana Saccharina. Exp. Bot. 2009, 66, 54–60. [Google Scholar] [CrossRef]
- Reynolds, O.L.; Padula, M.P.; Zeng, R.; Gurr, G.M. Silicon: Potential to Promote Direct and Indirect Effects on Plant Defense Against Arthropod Pests in Agriculture. Front. Plant Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, O.L.; Keeping, M.G.; Meyer, J.H. Silicon-Augmented Resistance of Plants to Herbivorous Insects: A Review. Ann. Appl. Biol. 2009, 155, 171–186. [Google Scholar] [CrossRef]
- Ma, J.F.; Nishimura, K.; Takahashi, E. Effect of Silicon on the Growth of Rice Plants at Different Growth Stages. Soil Sci. Plant Nutr. 1989, 35, 347–356. [Google Scholar] [CrossRef]
- Mohanty, S. Trends in Global Rice Consumption. In Rice Today; IRRI: Los Baños, Philippines, 2013; pp. 44–45. [Google Scholar]
- USDA-ERA. U.S. Acreage, Production, Yield and Farm Price. Available online: https://www.ers.usda.gov/data-products/rice-yearbook/rice-yearbook/#U.S.%20Acreage,%20Production,%20Yield,%20and%20Farm%20Price (accessed on 21 June 2014).
- USDA-ERA. Trade. Available online: https://www.ers.usda.gov/data-products/rice-yearbook/rice-yearbook/#U.S.%20Rice%20Trade (accessed on 21 June 2017).
- Way, M.O. Insect Pest Management in Rice in the United States. In Pest Management in Rice; Grayson, B.T., Green, M.B., Copping, L.G., Eds.; Elsevier Applied Science Publishers: Barking, UK, 1990; pp. 181–189. [Google Scholar]
- Aghaee, M.A.; Godfrey, L.D. A Century of Rice Water Weevil (Coleoptera:Curculionidae): A History of Research and Management with an Emphasis on the United States. J. Int. Pest Manag. 2014, 5, D1–D14. [Google Scholar] [CrossRef]
- Saito, T.; Hirari, K.; Way, M.O. The Rice Water Weevil Lissorhoptrus Oryzophilus Kuschel (Coleoptera: Curculionidae). Appl. Entomol. Zool. 2005, 40, 31–39. [Google Scholar] [CrossRef]
- Stout, M.J.; Rice, W.C.; Riggio, R.M.; Ring, D.R. The Effects of Four Insecticides on the Population Dynamics of the Rice Water Weevil Lissorhoptrus Oryzophilus Kuschel. J. Entomol. Sci. 2000, 35, 48–61. [Google Scholar]
- Shang, H.; Stout, M.J.; Zhang, Z.; Cheng, J. Rice Water Weevil (Coleoptera: Curculionidae) Population Dynamics in Louisiana. J. Entomol. Sci. 2004, 39, 623–642. [Google Scholar]
- Stout, M.J.; Rice, W.C.; Ring, D.R. The Influence of of Plant Age on Tolerance of Rice to Injury by the Rice Water Weevil, Lissorhoptrus Oryzophilus (Coleoptera Curculionidae). Bull. Entomol. Res. 2002, 92, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Stout, M.J.; Riggio, R.M.; Zou, L.; Roberts, R. Flooding Influences Ovipositional and Feeding Behaviour of the Rice Water Weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 2002, 95, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Stout, M.J.; Ring, D.R. Degree-Day Models for Emergence and Development of the Rice Water Weevil (Coleoptera: Curculionidae) in Southwestern Louisiana. Environ. Entomol. 2004, 33, 1541–1548. [Google Scholar] [CrossRef]
- Zou, L.; Stout, M.J.; Ring, D.R. Density-Yield Relationships for Rice Water Weevil on Rice for Different Varieties and under Different Water Management Regimes. Crop Prot. 2004, 23, 543–550. [Google Scholar] [CrossRef]
- Akinsola, E.A. Effects of Rice Stem Borer Infestation on Grain Yiled and Yield Components. Insect Sci. Appl. 1984, 5, 91–94. [Google Scholar]
- Bowling, C.C. Insect Pests of Rice in the United States. In The Major Insect Pests of the Rice Plant, Proceedings of the Symposium at the International Rice Research Institute, Los Banos, Laguna, Philippines, September 1964; John Hopkins Press: Baltimore, MD, USA, 1967; pp. 551–570. [Google Scholar]
- Reay-Jones, F.P.F.; Reagan, T.E.; Way, M.O.; Legendre, B.L. Concepts of Area Wide Management of the Mexican Rice Borer (Lepidoptera: Crambidae). Sugarcane Int. 2005, 23, 20–24. [Google Scholar]
- Pathak, M.D.; Khan, Z.R. Insect Pest of Rice; International Rice Research Institute: Los Banos, Philippines, 1994. [Google Scholar]
- Way, M.O.; Reay-Jones, F.P.F.; Reagan, T.E. Resistance to Stem Borers (Lepidoptera: Crambidae) among Texas Cultivars. J. Econ. Entomol. 2006, 99, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Reay-Jones, F.P.F.; Way, M.O.; Reagan, T.E. Economic Assessment of Controlling Stem Borers (Lepidoptera: Crambidae). Crop Prot. 2007, 26, 963–970. [Google Scholar] [CrossRef]
- Hummel, N.A.; Stout, M.J. Rice Water Weevil Control with Dermacor X-100 Seed Treatment. Arthropod Manag. Test 2010, 35, F36. [Google Scholar]
- Stout, M.J.; Hamm, J.C.; Frey, M.J.; Hummel, N.A. Comparison of the Efficacies of CruisserMaxx and Dermacor X-100 against Rice Water Weevil at Three Planting Dates; LSU AgCenter: Baton Rouge, LA, USA, 2011. [Google Scholar]
- Way, M.O. Reduced Rates of Dermacor on Non-Hybrid Rice; Texas AgriLife Research and Extension Center: Beaumont, TX, USA, 2010. [Google Scholar]
- Sidhu, J.K.; Hardke, J.T.; Stout, M.J. Efficacy of Dermacor-X-100 Seed Treatment Diatraea Saccharalis (Lepidoptera: Crambidae) on Rice. Fla. Entomol. 2014, 97, 224–232. [Google Scholar] [CrossRef]
- Hesler, L.S.; Grigarick, A.A.; Oraze, M.J.; Palrang, A.T. Effects of Temporary Drainage on Selected Life History Stages of the Rice Water Weevil (Coleoptera: Curculionidae) in California. J. Econ. Entomol. 1992, 85, 950–956. [Google Scholar] [CrossRef]
- Stout, M.J.; Harrell, D.; Tindall, K.V.; Bond, J. Impacts of Seeding Rate on Interactions between Rice and Rice Water Weevils. J. Econ. Entomol. 2009, 102, 1837–1845. [Google Scholar] [CrossRef] [PubMed]
- Stout, M.J.; Rice, W.C.; Linscombe, S.D.; Bollich, P.K. Identification of Rice Cultivars Resistant to Lissorhoptrus Oryzophilus (Coleoptera: Curculionidae), and Their Use in an Integrated Management Program. J. Econ. Entomol. 2001, 94, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.M.; Robinson, J.F. Evaluation of Rice Cultivars Grown in North America for Resistance to the Rice Water Weevil (Coleoptera: Curculiondae). Environ. Entomol. 1982, 11, 334–336. [Google Scholar] [CrossRef]
- McClung, A.M. Techniques for Development of New Cultivars. In Rice: Origin, History, Technology, and Production; Smith, C.W., Ed.; Wiley: Hoboken, NJ, USA, 2003; pp. 177–202. [Google Scholar]
- Chaudhary, R.C.; Khush, G.S.; Heinrichs, E.A. Varietal Resistance to Rice Stem Borers in Asia. Asia Insect Sci. Appl. 1984, 5, 447–463. [Google Scholar] [CrossRef]
- Way, M.O.; McCauley, G.M.; Zhou, X.G.; Wilson, L.T.; Morace, B. 2014 Texas Rice Production Guidelines; Texas AgriLife Research and Extension Center: Beaumont, TX, USA, 2014. [Google Scholar]
- Frew, A.; Powell, J.R.; Sallam, N.; Allsopp, P.G.; Johnson, S.N. Trade-Offs between Silicon and Phenolic Defenses May Explain Enhanced Performance of Root Herbivores on Phenolic-Rich Plants. J. Chem. Ecol. 2016, 42, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F. Role of Silicon in Enhancing the Resistance of Plants to Biotic and Abiotic Stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Goussain, M.M.; Prado, E.; Moraes, J.C. Effect of Silicon Applied to Wheat Plants on the Biology and Probing Behaviour of the Greenbug Schizaphis Graminum (Rond.) (Hemiptera: Aphididae). Neotrop. Entomol. 2005, 34, 807–813. [Google Scholar] [CrossRef]
- Massey, F.P.; Hartley, S.E. Physical Defenses Wear You down: Progressive Irreversible Impacts of Silica on Insect Herbivores. J. Anim. Ecol. 2009, 78, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.B.; Moraes, J.C.D.; Santos, C.D.D.; Goussain, M.M. Resistance Induction in Wheat Plants by Silicon and Aphids. Sci. Agric. 2005, 62, 547–551. [Google Scholar] [CrossRef]
- Hou, M.L.; Han, Y.Q. Silicon-Mediated Rice Plant Resistance to Asiatic Rice Borer (Lepidoptera: Crambidae): Effects of Silicon Amendment and Rice Varietal Resistance. J. Econ. Entomol. 2010, 103, 1412–1419. [Google Scholar] [CrossRef] [PubMed]
- Chandramani, R.; Rajendran, R.; Muthiah, C.; Chinniah, C. Organic Source Induced Silica on Leaf Folder, Stem Borer and Gall Midge Population and Rice Yield. J. Biopest. 2010, 3, 423–427. [Google Scholar]
- Ishizuka, K. Nutrient Uptake at Different Stages of Growth. In The Mineral Nutrition in Rice; Santos, O.G., Ed.; John Hopkins Press: Baltimore, MD, USA, 1964; pp. 199–217. [Google Scholar]
- Deren, C.W. Plant Genotype, Silicon Concentration, and Silicon-Related Responses. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndorfer, G.H., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 2001; pp. 149–158. [Google Scholar]
- Babu, T.; Tubana, B.; Datnoff, L.E.; Wang, J.; Yzenas, J. Release Pattern of Monosilicic Acid from Different Sources of Silicon in Soils of Louisiana. In Proceedings of the ASA-CSSA-SSSA International Annual Meetings, Long Beach, CA, USA, 31 October–6 November 2014. [Google Scholar]
- Sasamoto, K. Studies on the Relation between Insect Pests and Silica Content in Rice Plant (II) on the Injury of the Second Generation Larvae of Rice Stem Borer. Oyo Kontyu 1953, 9, 108–110. [Google Scholar]
- Panda, N.; Pradhan, B.; Samalo, A.P.; Rao, P.S.P. Note on the Relationship of Some Biochemical Factors with the Resistance in Rice Varieties to Yellow Stem Borer. Indian J. Agric. Sci. 1975, 45, 499–501. [Google Scholar]
- Nakata, Y.; Ueno, M.; Kihara, J.; Ichii, M.; Taketa, S.; Arase, S. Rice Blast Disease and Susceptibility to Pests in a Silicon Uptake-Deficient Mutant lsi1 of Rice. Crop Prot. 2008, 27, 865–868. [Google Scholar] [CrossRef]
- Sidhu, J.K.; Stout, M.J.; Blouin, D.C.; Datnoff, L.E. Effect of Silicon Soil Amendment on Performance of Sugarcane Borer, Diatraea Saccharalis (Lepidoptera: Crambidae) on Rice. Bull. Entomol. Res. 2013, 103, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.C.; Ma, T.S.; Li, F.J.; Feng, Y.J. Silicon Availability and Response of Rice and Wheat to Silicon in Calcareous Soils. Commun. Soil Sci. Plan 1994, 25, 2285–2297. [Google Scholar] [CrossRef]
- Lahm, G.P.; Stevenson, T.M.; Selby, T.P.; Freudenberger, J.H.; Cordova, D.; Flexner, L.; Bellin, C.A.; Dubas, C.M.; Smith, B.K.; Hughes, K.A.; et al. Rynaxpyr: A New Insecticidal Anthranilic Diamide That Acts as a Potent Amd Selective Ryanodine Receptor Activator. Bioorg. Med. Chem. Lett. 2007, 17, 6274–6279. [Google Scholar] [CrossRef] [PubMed]
- Lahm, G.P.; Cordova, D.; Barry, J.D. New and Selective Ryanodine Receptor Activators for Insect Control. Bioorg. Med. Chem. 2009, 17, 4127–4133. [Google Scholar] [CrossRef] [PubMed]
- Lanka, S.K.; Stout, M.J.; Beuzelin, J.M.; Ottea, J.A. Activity of Chlorantraniliprole and Thiametoxam Seed Treatments on Life Stages of the Rice Water Weevilas Affected by the Disruption of Insecticides in Rice Plants. Pest Manag. Sci. 2014, 70, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Hummel, N.A.; Meszaros, A.; Ring, D.R.; Beuzelin, J.M.; Stout, M.J. Evaluation of Seed Treatment Insecticides of the Rice Water Weevil, Lissorhoptrus Oryzophilus Kuschel (Coleoptera: Curculionidae), in Commercial Rice Fields in Louisiana. Crop Prot. 2014, 65, 37–42. [Google Scholar] [CrossRef]
- Lanka, S.K.; Ottea, J.A.; Beuzelin, J.M.; Stout, M.J. Effects of Chlorantraniliprole and Thiametoxam Rice Seed Treatment on Egg Numbers and First Instar Survival of Lissorhoptrus Oryzophilus (Coleoptera: Curculionidae). J. Econ. Entomol. 2013, 106, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Lanka, S.K.; Ottea, J.A.; Davis, J.A.; Hernandez, A.B.; Stout, M.J. Systemic Effects of Thiametoxam and Chlorantraniliprole Seed Treatments on Adult Lissorhoptrus Oryzophilus (Coleoptera: Curculionidae) in Rice. Pest Manag. Sci. 2013, 69, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Mohammad Saad, M.; Rahaman, M.M.; Stout, M.J. Varietal Resistance against Rice Water Weevil, Lissorhoptrus Oryzophilus (Coleoptera: Curculionidae) in Field and Greenhouse Studies; Louisiana State University: Baton Rouge, LA, USA, 2017. [Google Scholar]
- Sidhu, J.K.; Stout, M.J.; Blouin, D.C. Performance and Preference of of Sugarcane Borer, Diatraea Saccharalis, on Different Rice Cultivars. Entomol. Exp. Appl. 2013, 103, 656–664. [Google Scholar]
- Hamm, J.C.; Sidhu, J.K.; Stout, M.J.; Hummel, N.A.; Reagan, T.E. Oviposition Behaviour of Diatraea Saccharalis (Lepidoptera: Crambidae) on Different Rice Cultivars in Louisiana. Environ. Entomol. 2011, 41, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Way, M.O.; Reay-Jones, F.P.F.; Stout, M.J.; Tarpley, L. Effects of Nitrogen Fertilizer Applied before Permanent Flood on the Interaction between Rice and Rice Water Weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 2006, 99, 2030–2037. [Google Scholar] [CrossRef] [PubMed]
- VanWeelden, M.T.; Wilson, B.E.; Beuzelin, J.M.; Reagan, T.E.; Way, M.O. Impact of Nitrogen Fertilization on Mexican Rice Borer (Lepidoptera: Crambidae) Injury and Yield in Bioenergy Sorghum. Crop Prot. 2016, 84, 37–43. [Google Scholar] [CrossRef]
- Lopez, E.; Fernandez, C.; Lopez, O. Effects of Nitrogen Fertilization on Diatraea Saccharalis (Fabr.) Incidence on Sugar Cane. In International Society of Sugarcane Technologists; Wiley: Hoboken, NJ, USA, 1983; pp. 910–913. [Google Scholar]
- Wilson, C.E.; Wamishe, Y.; Lorenz, G.; Hardke, J.T. Rice Stand Establishment. In Arkansas Rice Production Handbook; University of Arkansas, Division of Agriculture, Research and Extension: Lawrence, KS, USA, 2013; pp. 31–40. [Google Scholar]
- SAS Institute Inc. SAS/STAT User’s Guide, version 9.4; SAS Institute: Cary, NC, USA, 2013. [Google Scholar]
- Tukey, J.W. The Problem of Multiple Comparisons; Department of Statistics, Princeton University: Princeton, NJ, USA, 1953. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Larval Density | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2013 | 2014 | 2015 | ||||||||||
| Core 1 | Core 2 | Core 1 | Core 2 | Core 1 | Core 2 | |||||||
| F1, 23 | P | F1, 23 | P | F1, 23 | P | F1, 24 | P | F1, 23 | P | F1, 24 | P | |
| Var | 1.33 | 0.2607 | 0.14 | 0.7095 | 6.93 | 0.0149 | 5.47 | 0.0281 | 0.01 | 0.9117 | 0.61 | 0.4439 |
| Derm | 48.37 | <0.0001 | 68.50 | <0.0001 | 111.66 | <0.0001 | 54.62 | <0.0001 | 95.88 | <0.0001 | 55.33 | <0.0001 |
| Sil | 1.68 | 0.2077 | 2.25 | 0.1473 | 4.60 | 0.0427 | 1.46 | 0.2393 | 0.03 | 0.8591 | 0.00 | 1.000 |
| Var × Der | 10.65 | 0.0034 | 18.54 | 0.0003 | 0.72 | 0.4040 | 0.57 | 0.4581 | 8.57 | 0.0076 | 16.53 | 0.0004 |
| Var × Sil | 0.76 | 0.3908 | 0.39 | 0.5387 | 1.79 | 0.1946 | 0.00 | 1.000 | 0.01 | 0.9400 | 0.37 | 0.5506 |
| Derm × Sil | 0.39 | 0.5365 | 0.56 | 0.4631 | 6.93 | 0.0149 | 0.96 | 0.3367 | 2.90 | 0.1019 | 0.27 | 0.6085 |
| Var × Derm × Sil | 1.03 | 0.3214 | 0.08 | 0.7839 | 3.32 | 0.0815 | 0.46 | 0.5038 | 0.58 | 0.4523 | 0.19 | 0.6693 |
| Activity | Year | ||
|---|---|---|---|
| 2013 | 2014 | 2015 | |
| Seed Treatment | 20 May | 5 May | 8 June |
| Planting | 21 May | 9 May | 10 June |
| Silicon Slag Application | 21 May | 9 May | 10 June |
| Stand Count | 6 June | 2 June | 6 July |
| Permanent Flood | 19 June | 13 June | 10 July |
| 1st Core Sampling | 10 July | 8 July | 31 July |
| 2nd Core Sampling | 17 July | 15 July | 7 August |
| Whitehead counts | 16 August | 12 August | 14 September |
| Harvest | 13 September | 9 September | 6 October |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villegas, J.M.; Way, M.O.; Pearson, R.A.; Stout, M.J. Integrating Soil Silicon Amendment into Management Programs for Insect Pests of Drill-Seeded Rice. Plants 2017, 6, 33. https://doi.org/10.3390/plants6030033
Villegas JM, Way MO, Pearson RA, Stout MJ. Integrating Soil Silicon Amendment into Management Programs for Insect Pests of Drill-Seeded Rice. Plants. 2017; 6(3):33. https://doi.org/10.3390/plants6030033
Chicago/Turabian StyleVillegas, James M., Michael O. Way, Rebecca A. Pearson, and Michael J. Stout. 2017. "Integrating Soil Silicon Amendment into Management Programs for Insect Pests of Drill-Seeded Rice" Plants 6, no. 3: 33. https://doi.org/10.3390/plants6030033
APA StyleVillegas, J. M., Way, M. O., Pearson, R. A., & Stout, M. J. (2017). Integrating Soil Silicon Amendment into Management Programs for Insect Pests of Drill-Seeded Rice. Plants, 6(3), 33. https://doi.org/10.3390/plants6030033