Genus-Specific Real-Time PCR and HRM Assays to Distinguish Liriope from Ophiopogon Samples

 and
and
Abstract
:1. Introduction
2. Results
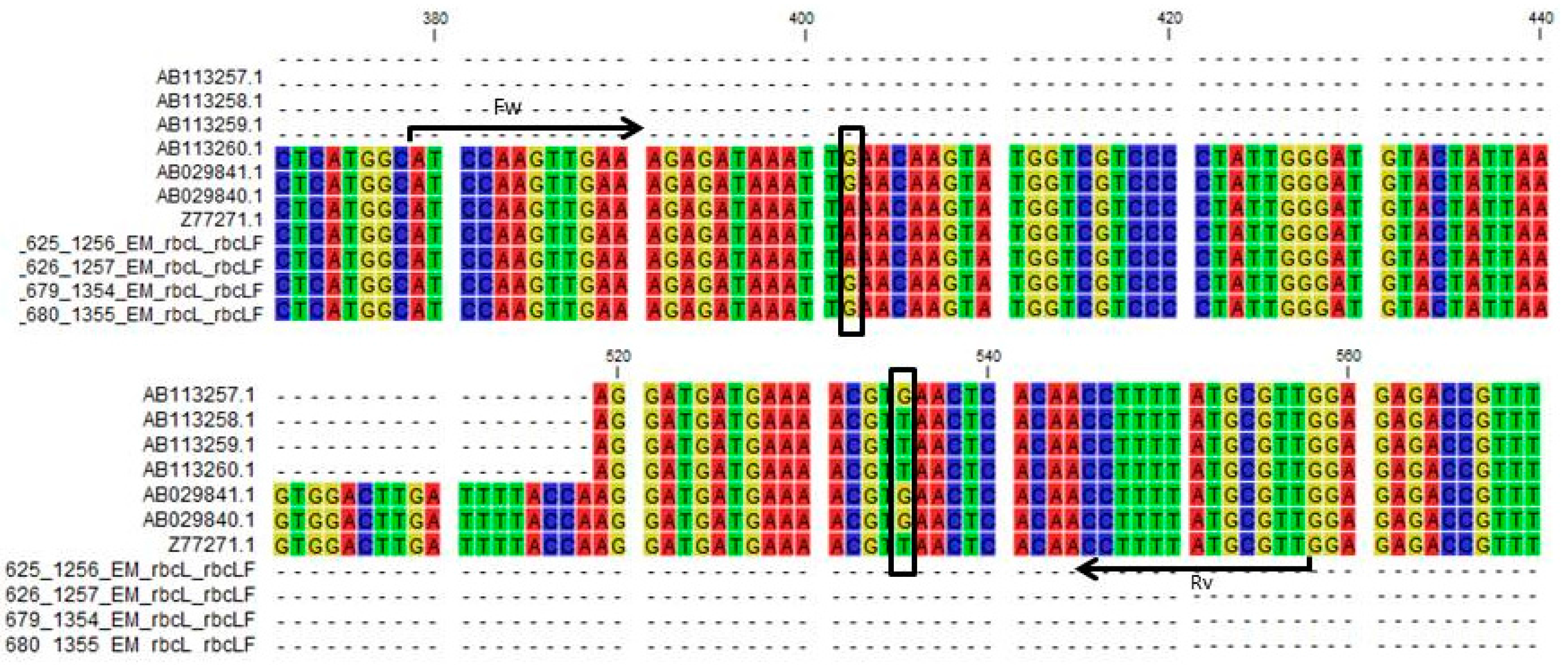
2.1. DNA Barcoding of the rbcLa Region of Liriope and Ophiopogon Accessions
2.2. The rbcL Feature Provides a Target for Simple Genus Discriminatory Tests
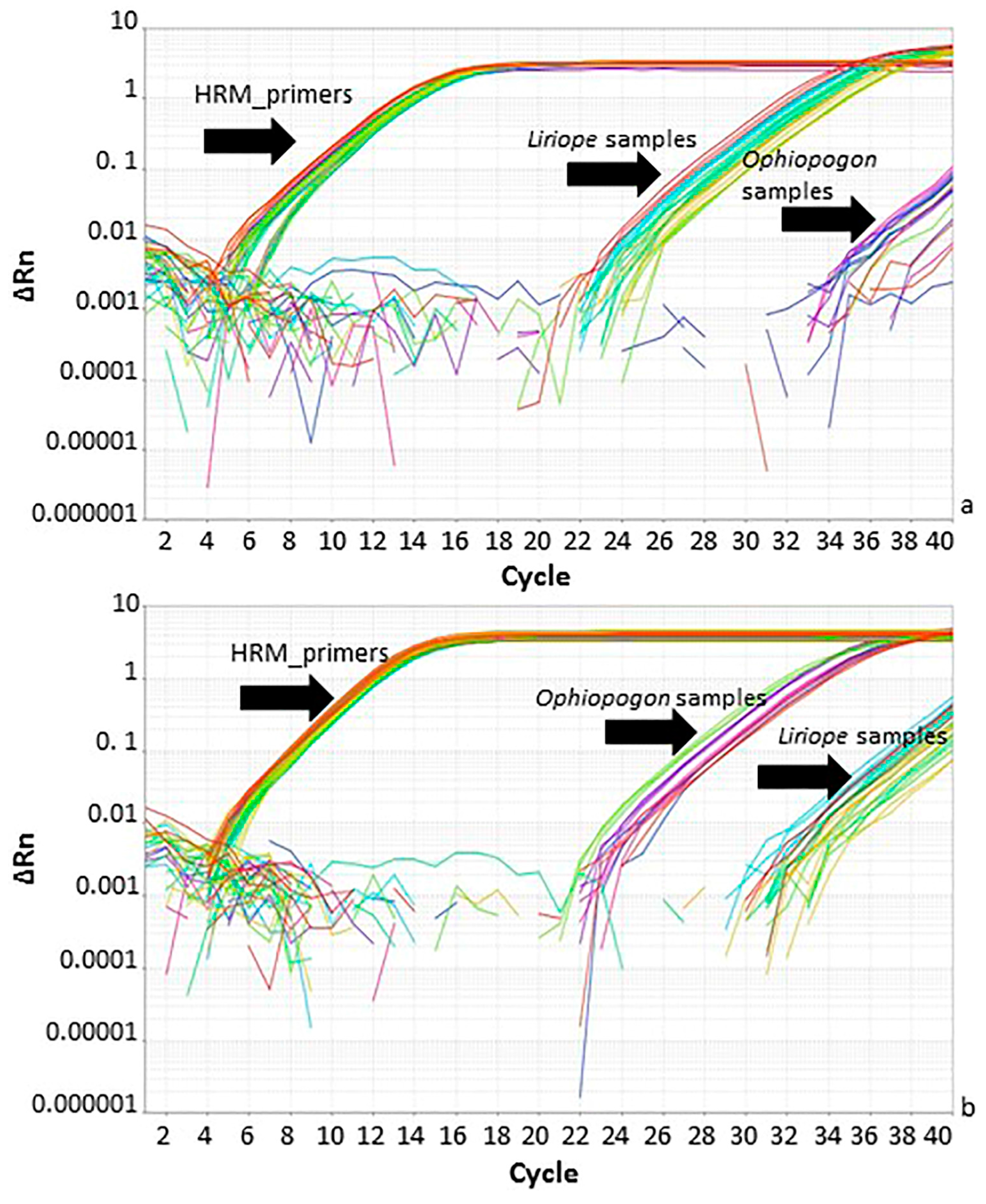
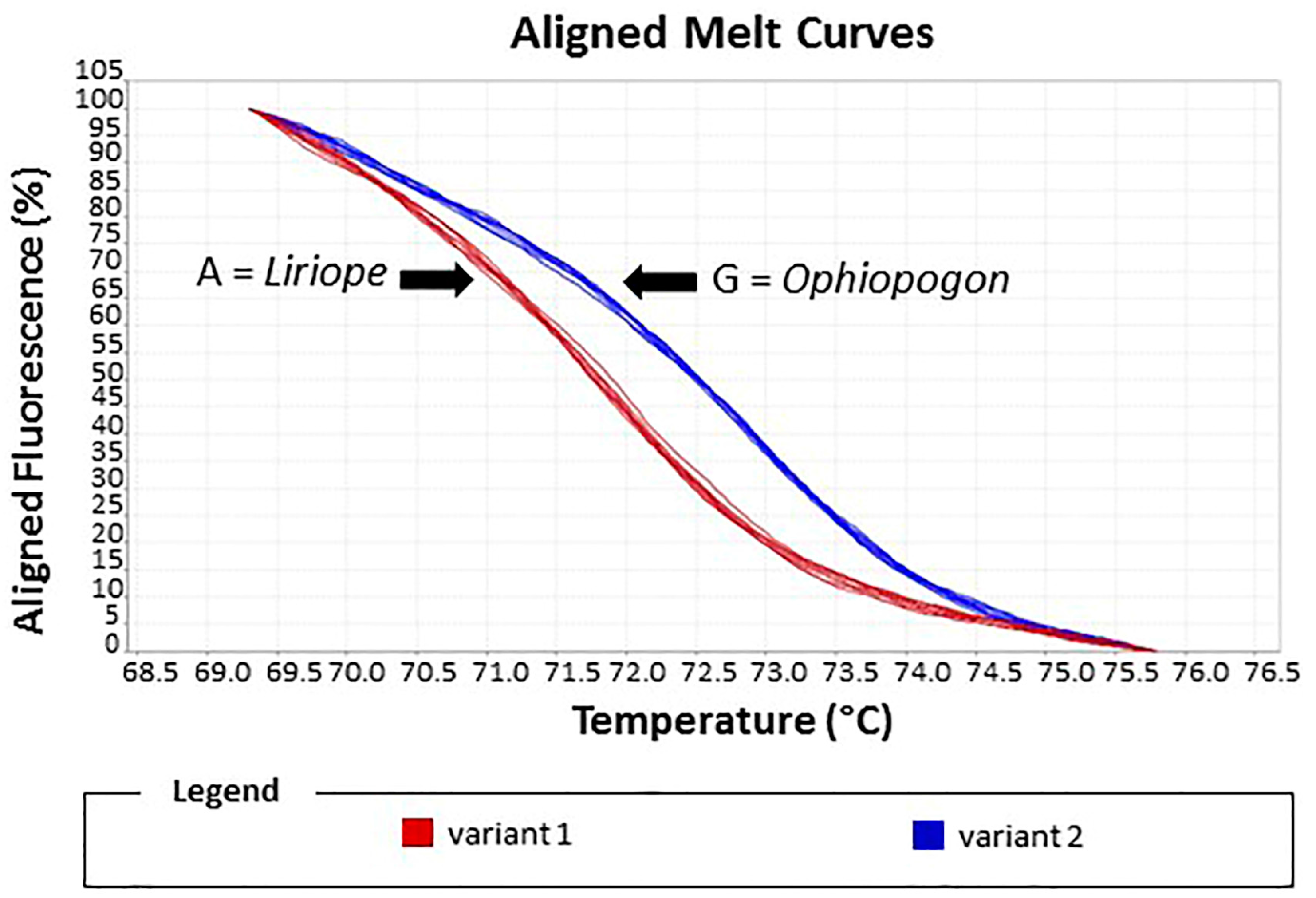
2.3. Identification of Liriope and Ophiopogon Samples by Using the HRM System
3. Discussion
4. Materials and Methods
4.1. Plant Material and Total DNA Extraction
4.2. PCR Protocols
- rbcLa PCR: initial denaturation step of 5 min at 95 °C followed by 35 cycles consisting of 30 s at 95 °C, 20 s at 52 °C, and 50 s at 72 °C, with a final extension period of 5 min at 72 °C.
- Ophiopogon-specific rbcL PCR: initial denaturation step of 5 min at 95 °C followed by 40 cycles consisting of 30 s at 95 °C, 30 s at 62.5 °C, and 45 s at 72 °C, with a final extension period of 5 min at 72 °C.
- Liriope-specific rbcL PCR: initial denaturation step of 5 min at 95 °C followed by 35 cycles consisting of 30 s at 95 °C, 30 s at 61 °C, and 30 s at 72 °C, with a final extension period of 5 min at 72 °C.
- HRM (generic) rbcL PCR: initial denaturation step of 5 min at 95 °C followed by 35 cycles consisting of 30 s at 95 °C, 30 s at 60 °C, and 30 s at 72 °C, with a final extension period of 5 min at 72 °C.
4.3. DNA Sequence Analysis
4.4. Real-Time PCR Analysis
4.5. High-Resolution Melt Curve Analysis (HRM) Methods
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| nrITS | nuclear ribosomal internal transcribed spacer |
| rbcL | large subunit of ribulose-1,5-bisphosphate carboxylase/oxygenase gene |
| HRM | high-resolution melt curve assay |
References
- Fantz, P.R. Species of Liriope Cultivated in the Southeastern United States. HortTechnology 2008, 18, 343–348. [Google Scholar]
- Fantz, P.R. Names and Species of Ophiopogon Cultivated in the Southeastern United States. HortTechnology 2009, 19, 385–394. [Google Scholar]
- Nesom, G.L. Overview of Liriope and Ophiopogon (Ruscaceae) naturalized and commonly cultivated in the USA. Phytoneuron 2010, 56, 1–31. [Google Scholar]
- Lattier, J.D.; Ranney, T.G. Identification, Nomenclature, Genome Sizes, and Ploidy Levels of Liriope and Ophiopogon Taxa. HortScience 2014, 49, 145–151. [Google Scholar]
- Mabberley, D.J. A Portable Dictionary of Plants, Their Classification and Uses; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Wu, Y.; Dong, Z.; Wu, H.; Ding, W.; Zhao, M.; Shi, Q.; Wang, Q. Comparative studies on Ophiopogonis and Liriopes based on the determination of 11 bioactive components using LC–MS/MS and hierarchical clustering analysis. Food Res. Int. 2014, 57, 15–25. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, H.; Shen, L.; Wang, W. Novel steroidal saponins from Liriope graminifolia (Linn.) Baker with anti-tumor activities. Carbohydr. Res. 2011, 346, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Che, Y.; Zhang, L.; Zhang, J.; Zhou, Y.; Jiang, Y.; Tu, P. Fingerprint analysis of Ophiopogonis Radix by HPLC-UV-ELSD coupled with chemometrics methods. J. Chin. Pharm. Sci. 2013, 22, 55–63. [Google Scholar] [CrossRef]
- Tang, X.; Cheng, Z.; Yu, B. Qualitative and quantitative studies on Chinese drug Maidong. China J. Chin. Mater. Med. 1999, 24, 390–445. [Google Scholar]
- Zeng, P.; Zhou, H.; Zheng, Y.; Xu, X.; Fu, S. Simultaneous determination of three homoisoflavonoids in Ophiopogon japonicus by HPLC. China J. Chin. Mater. Med. 2012, 37, 71–74. [Google Scholar]
- Mishra, P.; Kumar, A.; Nagireddy, A.; Mani, D.N.; Shukla, A.K.; Tiwari, R.; Sundaresan, V. DNA barcoding: An efficient tool to overcome authentication challenges in the herbal market. Plant Biotechnol. J. 2016, 14, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Coutinho Moraes, D.F.; Still, D.W.; Lum, M.R.; Hirsch, A.M. DNA-Based Authentication of Botanicals and Plant-Derived Dietary Supplements: Where Have We Been and Where Are We Going? Planta Med. 2015, 81, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Sgamma, T.; Lockie-Williams, C.; Kreuzer, M.; Williams, S.; Scheyhing, U.; Koch, E.; Slater, A.; Howard, C. DNA Barcoding for Industrial Quality Assurance. Planta Med. 2017, 83, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Kazi, T.; Hussain, N.; Bremner, P.; Slater, A.; Howard, C. The application of a DNA-based identification technique to over-the-counter herbal medicines. Fitoterapia 2013, 87, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Howard, C.; Bremner, P.D.; Fowler, M.R.; Isodo, B.; Scott, N.W.; Slater, A. Molecular identification of Hypericum perforatum by PCR amplification of the ITS and 5.8S rDNA region. Planta Med. 2009, 75, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.K.; Parveen, I.; Raghuvanshi, S.; Babbar, S.B. The loci recommended as universal barcodes for plants on the basis of floristic studies may not work with congeneric species as exemplified by DNA barcoding of Dendrobium species. BMC Res. Notes 2012, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Cameron, K.M.; Chase, M.W.; Whitten, W.M.; Kores, P.J.; Jarrell, D.C.; Albert, V.A.; Yukawa, T.; Hills, H.G.; Goldman, D.H. A phylogenetic analysis of the Orchidaceae: Evidence from rbcL nucleotide. Am. J. Bot. 1999, 86, 208–224. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Park, J.Y. SCAR markers for discriminating species of two genera of medicinal plants, Liriope and Ophiopogon. Genet. Mol. Res. 2012, 29, 2987–2996. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Ra, W.; Park, J.-W.; Kwon, S.-W.; Lee, J.; Park, C.; Park, Y. Developing EST-SSR markers to study molecular diversity in Liriope and Ophiopogon. Biochem. Syst. Ecol. 2011, 39, 241–252. [Google Scholar] [CrossRef]
- Ito, M.; Sato-Masumoto, N.; Kobayashi, F.; Matsumura, K. Distinguishing Ophiopogon and Liriope tubers based on DNA sequences. J. Nat. Med. 2015, 69, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Abson, J. Botany, medicine and mistaken identity. Plant Heritage J. 2017, 25, 41–43. [Google Scholar]
- Shiba, M.; Yamaji, H.; Kondo, K.; Ichiki, H.; Sakakibara, I.; terabayashi, S.; Amagaya, S.; Aburada, M.; Miyamoto, K. Discrimination of Maidong Derived from Ophiopogon and Liriope Species by rbcL Sequences, and Their Chemical Components and Tuber Anatomy. Nat. Med. 2004, 58, 15–21. [Google Scholar]
- Dong, W.; Cheng, T.; Li, C.; Xu, C.; Long, P.; Chen, C.; Zhou, S. Discriminating plants using the DNA barcode rbcLb: An appraisal based on a large data set. Mol. Ecol. Res. 2013, 14, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Applied Biosystems StepOne™ and StepOnePlus™ Real-Time PCR Systems Reagent Guide. Available online: https://assets.thermofisher.com/TFS-Assets/LSG/manuals/cms_046739.pdf (accessed on 25 October 2017).
- Applied Biosystems High Resolution Melting Getting Started Guide. Available online: https://tools.thermofisher.com/content/sfs/manuals/cms_050347.pdf (accessed on 25 October 2017).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Number | ΔCt Liriope-Specific | ΔCt Ophiopogon-Specific | ΔΔCt Liriope-Specific | ΔΔCt Ophiopogon-Specific |
|---|---|---|---|---|
| 624 | 12.41 | 23.53 | 0 | 8.61 |
| 626 | 13.28 | 24.21 | 0.87 | 9.29 |
| 628 | 25.51 | 13.00 | 13.10 | −1.92 |
| 633 | 12.28 | 23.64 | −0.13 | 8.72 |
| 634 | 11.67 | 22.91 | −0.74 | 7.99 |
| 667 | 24.74 | 15.17 | 12.33 | 0.25 |
| 677 | 24.46 | 13.87 | 12.05 | −1.05 |
| 678 | 24.34 | 14.92 | 11.92 | 0 |
| 679 | 25.89 | 15.45 | 13.48 | 0.53 |
| 693 | 13.76 | 25.68 | 1.35 | 10.76 |
| 695 | 14.43 | 25.62 | 2.02 | 10.70 |
| 709 | 13.03 | 24.33 | 0.62 | 9.41 |
| 711 | 12.56 | 22.55 | 0.15 | 7.63 |
| Primers | Sequences | Annealing Temperature (Conventional PCR) | Annealing Temperature (Real-Time PCR) | Amplicon Size (bp) |
|---|---|---|---|---|
| rbcLFw | ATGTCACCACAAACAGAGACTAAAGC | 52 °C | N/A | 700 |
| rbcLRv | GTAAAATCAAGTCCACCRCG | |||
| rbcL_Liriope_Fw | ATCCAAGTTGAAAGAGATAAATTA | 61 °C | 61 °C | 180 |
| rbcL_Liriope_Rv | AACGCATAAAAGGTTGTGAGTTA | |||
| rbcL_Ophiopogon_Fw | ATCCAAGTTGAAAGAGATAAATTG | 62.5 °C | 64.5 °C | 180 |
| rbcL_Ophiopogon_Rv | AACGCATAAAAGGTTGTGAGTTC | |||
| HRM_rbcL_Fw | CGCCTCATGGCATCCAAGT | 61 °C | 61 °C | 80 |
| HRM_rbcL_Rv | AATAGGGGACGACCATACTTG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masiero, E.; Banik, D.; Abson, J.; Greene, P.; Slater, A.; Sgamma, T. Genus-Specific Real-Time PCR and HRM Assays to Distinguish Liriope from Ophiopogon Samples. Plants 2017, 6, 53. https://doi.org/10.3390/plants6040053
Masiero E, Banik D, Abson J, Greene P, Slater A, Sgamma T. Genus-Specific Real-Time PCR and HRM Assays to Distinguish Liriope from Ophiopogon Samples. Plants. 2017; 6(4):53. https://doi.org/10.3390/plants6040053
Chicago/Turabian StyleMasiero, Eva, Dipanwita Banik, John Abson, Paul Greene, Adrian Slater, and Tiziana Sgamma. 2017. "Genus-Specific Real-Time PCR and HRM Assays to Distinguish Liriope from Ophiopogon Samples" Plants 6, no. 4: 53. https://doi.org/10.3390/plants6040053
APA StyleMasiero, E., Banik, D., Abson, J., Greene, P., Slater, A., & Sgamma, T. (2017). Genus-Specific Real-Time PCR and HRM Assays to Distinguish Liriope from Ophiopogon Samples. Plants, 6(4), 53. https://doi.org/10.3390/plants6040053