Nitrogen Nutrition of Fruit Trees to Reconcile Productivity and Environmental Concerns




Abstract
:1. Introduction
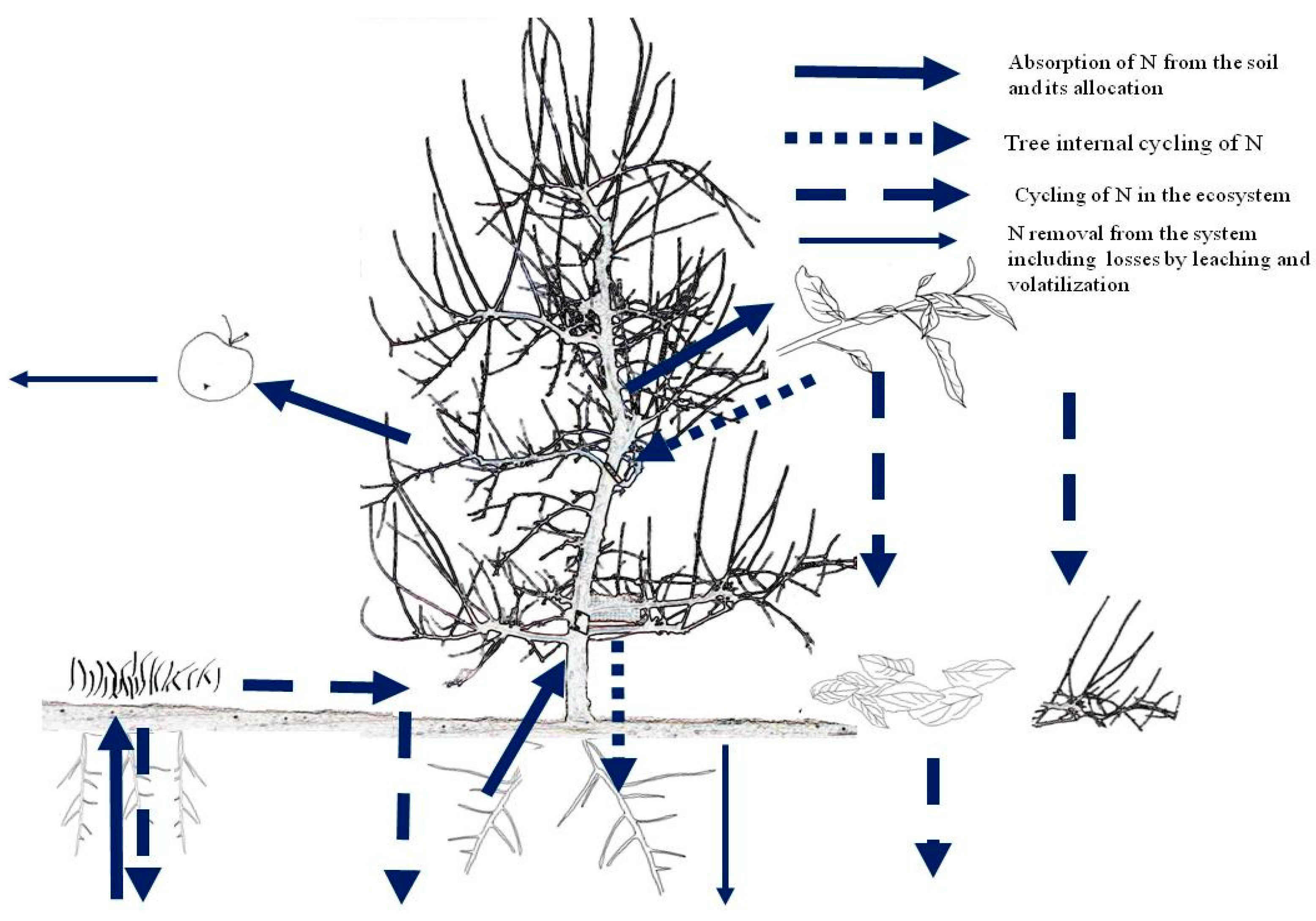
2. Tree N Uptake and Internal Cycling
3. Nitrogen Fertilization of Fruit Trees
3.1. Nitrogen Needs
3.2. Nitrogen Management in Young Orchards
3.3. Nitrogen Management of Mature Fruit Trees
4. Soil Management to Enhance Fertilizer Nitrogen Use Efficiency
5. Efficient Nitrogen Supply Methods
6. Future Directions
Author Contributions
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). How to Feed the World in 2050; FAO: Quebec, QC, Canada, 2009; p. 35. [Google Scholar]
- Pathak, H.; Nedwell, D.B. Nitrous oxide emission from soil with different fertilizer, water levels and nitrification inhibitors. Water Air Soil Pollut. 2011, 129, 217–228. [Google Scholar] [CrossRef]
- Srivastava, A.K. New paradigms in soil health management in fruit crops for improving farmers’ income. Shodh Chintan 2017, 9, 191–224. [Google Scholar]
- Khan, Z.I.; Ahmad, K.; Ashraf, M.; Parveen, R.; Bibi, Z.; Mustafa, I.; Noorka, I.R.; Tahir, H.M.; Akram, N.A.; Ullah, M.F.; et al. Risk assessment of heavy metal and metalloid toxicity through a contaminated vegetable (Cucurbita maxima) from wastewater irrigated area: A case study for a site-specific risk assessment in Jhang, Pakistan. Hum. Ecol. Risk Assess. 2016, 22, 86–98. [Google Scholar] [CrossRef]
- Verzeaux, J.; Hirel, B.; Dubois, F.; Lea, P.J.; Tétu, T. Agricultural practices to improve nitrogen use efficiency through the use of arbuscular mycorrhizae: Basic and agronomic aspects. Plant Sci. 2017, 264, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and interpretation of factors which contribute to efficiency to nitrogen utilization. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]
- Carranca, C. Nitrogen use efficiency by annual and perennial crops. In Farming for Food and Water Security; Series: Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Springer Science + Business Media: Dordrecht, The Netherlands, 2012; Volume 10, pp. 57–82. [Google Scholar]
- Food and Agriculture Organization (FAO). Statistical Year Book of Food and Agricultural Organization; FAO: Quebec, QC, Canada, 2011; pp. 108–113. [Google Scholar]
- Tagliavini, M.; Quartieri, M.; Millard, P. Remobilised nitrogen and root uptake of nitrate for leaf growth, flowers and developing fruits of pear (P. communis) trees. Plant Soil 1997, 195, 137–142. [Google Scholar] [CrossRef]
- Menino, M.R.; Carranca, C.; De Varennes, A. Distribution and remobilization of nitrogen in young non-bearing orange trees grown under Mediterranean conditions. J. Plant Nutr. 2007, 30, 1083–1096. [Google Scholar] [CrossRef]
- Neto, C.; Carranca, C.; Clemente, J.; De Varennes, A. Nitrogen distribution, remobilization and re-cycling in young orchard of non-bearing ‘Rocha’ pear trees. Sci. Hortic. 2008, 118, 299–307. [Google Scholar] [CrossRef]
- Zekri, M.; Obreza, T. Nitrogen (N) for Citrus Trees. In SL378—A Series of the Department of Soil and Water Sciences; UF/IFAS Extension, University of Florida: Gainesville, FL, USA, 2013; p. 3. [Google Scholar]
- Stellacio, A.M.; Cristiano, G.; Rubino, P.; De Lucia, B.; Cazzat, E. Nitrogen uptake, nitrogen partitioning and N-use efficiency of container-grown holm oak (Quercus ilex L.) under different nitrogen levels and fertilizer sources. J. Food Agric. Environ. 2013, 11, 990–994. [Google Scholar]
- Millard, M. Internal cycling of nitrogen in trees. Acta Hortic. 1995, 383, 3–14. [Google Scholar] [CrossRef]
- Brunetto, G.; De Melo, G.W.B.; Toseli, M.; Quatieri, M.; Tagliavini, M. The role of mineral nutrition on yields and fruit quality in grapevine, pear and apple. Rev. Bras. Frutic. 2015, 3, 4–1104. [Google Scholar] [CrossRef]
- Quartieri, M.; Millard, P.; Tagliavini, M. Storage and remobilization of nitrogen by pear (Pyrus communis L.) trees as affected by timing of N supply. Eur. J. Agron. 2002, 17, 105–110. [Google Scholar] [CrossRef]
- Tromp, J.; Ovaa, J.C. Uptake and distribution of nitrogen in young apple trees after application of nitrate or ammonium, with special reference to asparagine and arginine. Physiol. Plant. 1979, 45, 23–28. [Google Scholar] [CrossRef]
- Näsholm, T.; Kielland, K.; Ganeteg, U. Uptake of organic nitrogen by plants. New Phytol. 2009, 182, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, M.D.; Digman, M.A.; Gratton, E.; Treseder, K.K. Organic nitrogen uptake by arbuscular mycorrhizal fungi in a boreal forest. Soil Biol. Biochem. 2012, 55, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: Cambridge, MA, USA, 2012; p. 651. [Google Scholar]
- Grassi, G.; Millard, P.; Gioacchini, P.; Tagliavini, M. Recycling of nitrogen in the xylem of Prunus avium trees starts when spring remobilization of internal reserves declines. Tree Physiol. 2003, 23, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Loescher, W.H.; McCamant, T.; Keller, J.D. Carbohydrate reserves, translocation, and storage in woody plant roots. Hortscience 1990, 25, 3–281. [Google Scholar]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition, 5th ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Niederholzer, J.A.; Dejong, T.M.; Sanz, J.L.; Muraoka, T.T.; Weinbaum, S.A. Effectiveness of fall vs. spring soil fertilization of field-grown peach trees. J. Am. Soc. 2001, 126, 5–648. [Google Scholar]
- Tromp, J. Nutrient reserves in roots of fruit trees, in particular carbohydrates and nitrogen. Plant Soil 1983, 71, 401–413. [Google Scholar] [CrossRef]
- Menino, M.R.; Carranca, C.; De Varennes, A.; Valente D’Almeida, V.; Baeta, J. Tree size and flowering intensity as affected by nitrogen fertilization in non-bearing orange trees grown under Mediterranean conditions. J. Plant Physiol. 2003, 160, 1435–1440. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Alcantara, B.; Quiñones, A.O.; Primi-Millo, E.; Legaz, F. Nitrogen remobilization response to current supply in young citrus trees. Plant Soil 2011, 342, 433–443. [Google Scholar] [CrossRef]
- Roccuzzo, G.; Scandellari, F.; Allegra, M.; Torrisi, B.; Stagno, F.; Mimmo, T.; Zanotelli, D.; Gioacchini, P.; Millard, P.; Tagliavini, M. Seasonal dynamics of root uptake and spring remobilisation of nitrogen in field grown orange trees. Sci. Hortic. 2017, 226, 223–230. [Google Scholar] [CrossRef]
- Legaz, F.; Serna, M.D.; Primo-Millo, E. Mobilization of the reserve N in citrus. Plant Soil 1995, 173, 205–210. [Google Scholar] [CrossRef]
- Wendler, R.; Carvalho, P.O.; Pereira, J.S.; Millard, P. Role of nitrogen remobilization from old leaves for new leaf growth of Eucalyptus globulus seedlings. Tree Physiol. 1995, 15, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Tagliavini, M.; Scandellari, F. Methodologies and concepts in the study of nutrient uptake requirements and partitioning in fruit trees. In Proceedings of the Seventh International Symposium on Mineral Nutrition of Fruit Crops, Chanthaburi, Thailand, 19–25 May 2012; pp. 19–25. [Google Scholar]
- Tagliavini, M.; Scandellari, F.; Toselli, M. La fertilizzazione dei sistemi frutticoli. In Fertilizzazione Sostenibile; Grignani, C., Ed.; Edagricole-New Business Media: Bologna, Italy, 2016; pp. 391–416. [Google Scholar]
- Tagliavini, M.; Failla, O.; Xiloyannis, C. La fertilizzazione nell’arboreto. In Arboricoltura Generale; di Silviero, S., Costa, G., Gucci, R., Eds.; Patron Editore: Bologna, Italy, 2012; pp. 425–443. [Google Scholar]
- Feigenbaum, S.; Bielorai, H.; Erner, Y.; Dasberg, S. The fate of 15N labelled nitrogen applied to mature citrus trees. Plant Soil 1987, 97, 179–187. [Google Scholar] [CrossRef]
- Ledgard, S.F.; Smith, G.S. Fate of 15N-labeled nitrogen fertilizer applied to kiwifruit (Actinidia deliciosa) vines. II. Temporal changes in 15N within vines. Plant Soil 1992, 147, 59–68. [Google Scholar] [CrossRef]
- Weinbaum, S.A.; Niederholzer, F.J.A.; Ponchner, S.; Rosecrance, R.C.; Carlson, R.M.; Whittlesey, A.C.; Muraoka, T.T. Nutrient uptake by cropping and defruited field-grown ‘French’ prune trees. J. Am. Soc. Hortic. Sci. 1994, 119, 925–930. [Google Scholar]
- Weih, M. A calculation tool for analyzing nitrogen use efficiency in annual and perennial crops. Agronomy 2014, 4, 470–477. [Google Scholar] [CrossRef]
- Gonçalves, M.; Cavaco, M.; Sousa, A.J.T.; Ferreira, J.T.; Jorge, O.; Avelar, M.L.; Sousa, R.; Costa, A.S.V.; Calouro, F.; Dias, J.C.S.; et al. Produção Integrada de Pomóideas—Planos de Fertilização e Práticas Culturais; DGDR, MADRP: Oeiras, Portugal, 1997. [Google Scholar]
- Luz, R.; Azevedo, J.; Calouro, F. Fertilização. In Manual Técnico de ProduçãoIntegrada de Pêra ‘Rocha´; APAS, INIAP/ENFVN, LQARS, DGPC, Eds.; Associação nacional de Produtos Pêra Rocha: Cadaval, Portugal, 2005; pp. 29–41. [Google Scholar]
- Cavaco, M.; Jordão, P.; De Sousa, R. Produção Integrada da Cultura de Pomóideas; MADRP/DGPC: Oeiras, Portugal, 2006. [Google Scholar]
- Quartieri, M.; Tagliavini, M.; Scudellari, D.; Marangoni, B.; Millard, P. Assorbimento radicale di azoto e rimobilizzazione delle reserve azotate nel pêro durante la ripresa vegetativa. Riv. Fruttic. 1996, 10, 57–60. [Google Scholar]
- Raese, J.T. Cold tolerance, yield and fruit quality of ‘d’Anjou’ pears influenced by nitrogen fertilizer rates and time of application. J. Plant Nutr. 1997, 20, 1007–1102. [Google Scholar] [CrossRef]
- Fracchiola, M.; Terzi, M.; Frabboni, L.; Caramia, D.; Lasorella, C.; De Giorgio, D.; Montemurro, P. Cazzato, E. Influence of different practices on ground-flora vegetation in an almond orchard. Renew. Agric. Food Syst. 2016, 31, 4–308. [Google Scholar] [CrossRef]
- Comin, J.J.; Loss, A.; Da Veiga, M.; Guardini, R.; Schmitt, D.E.; Oliveira, P.A.V.; Filho, P.B.; Couto, R.R.; Benedet, L.; Múller Júnior, V.M.; et al. Physical properties and organic carbon content of a Typic Hapludult soil fertilized with pig slurry and pig litter in a no-tillage system. Soil Res. 2013, 51, 459–470. [Google Scholar] [CrossRef]
- Lourenzi, C.R.; Ceretta, C.A.; Silva, L.S.; Trentin, G.; Girotto, E.; Lorensini, F.; Tiecher, T.L.; Brunetto, G. Soil chemical properties related to acidity under successive pig slurry application. Rev. Bras. Ciênc. Solo 2011, 35, 1827–1836. [Google Scholar] [CrossRef]
- Brunetto, G.; Comin, J.J.; Schmitt, D.E.; Guardini, R.; Mezzari, C.P.; Oliveira, B.S.; Moraes, M.P.; Gatiboni, L.C.; Lovato, P.E.; Ceretta, C.A. Changes in soil acidity and organic carbon in a sandy typic hapludalf after medium-term pig-slurry and deep-litter application. Rev. Bras. Ciênc. Solo 2012, 36, 10–20. [Google Scholar] [CrossRef]
- Altieri, M.A. Biodiversity and Pest Management in Agroecosystems; CRC Press: Boca Raton, FL, USA, 1994; p. 185. [Google Scholar]
- Neto, C.; Carranca, C.; Clemente, J. Senescent leaf decomposition in a Mediterranean pear orchard. Eur. J. Agron. 2009, 30, 34–40. [Google Scholar] [CrossRef]
- Tagliavini, M.; Tonon, G.; Solimando, D.; Gioacchini, P.; Toselli, M.; Boldreghini, P.; Ciavatta, C. Nitrogen uptake by ryegrass (Lolium perenne) as affected by the decomposition of apple leaves and pruning wood in soil. In Proceedings of the 12th N Workshop, Controlling N Flows and Losses. Wageningen; Hatch, D.J., Ed.; Academic Publishers: Wageningen, The Netherlands, 2004; pp. 239–241. [Google Scholar]
- Tagliavini, M.; Scandellari, F. Nutrient fluxes in kiwifruit orchards. Acta Hortic. 2007, 753, 487–494. [Google Scholar] [CrossRef]
- Ventura, M.; Scandellari, F.; Bonora, E.; Tagliavini, M. Nutrient release during decomposition of leaf litter in a peach (Prunus persica L.) orchard. Nutr. Cycl. Agroecosyst. 2010, 87, 115–125. [Google Scholar] [CrossRef]
- Tonon, G.; Ciavatta, C.; Solimando, D.; Gioacchini, P.; Tagliavini, M. Fate of 15N derived from soil decomposition of abscised leaves and pruning wood from apple (Malus domestica) trees. Soil Sci. Plant Nutr. 2007, 53, 78–85. [Google Scholar] [CrossRef]
- Banyal, S.K.; Sharma, S.K. Effect of fertigation and rootstock on yield and quality of apple. Indian J. Hortic. 2011, 68, 419–424. [Google Scholar]
- Shirgure, P.S.; Srivastava, A.K.; Singh, S. Economics of drip and fertigation in acid lime orchards. J. Soil Water Conserv. 2002, 46, 56–60. [Google Scholar]
- Baldi, E.; Toselli, M.; Marangoni, B. Nutrient partitioning in potted peach (Prunus persica L.) trees supplied with mineral and organic fertilizers. J. Plant Nutr. 2010, 33, 14–2062. [Google Scholar] [CrossRef]
- Ahmed, F.M.; Abhijit, S.; Abida, J. Response of sweet cherry (Prunus avium) to fertigation of nitrogen, phosphorus and potassium under Kerawa land of Kashmir valley. Indian J. Agric. Sci. 2010, 80, 512–516. [Google Scholar]
- Tagliavini, M.; Millard, P.; Quartieri, M. Storage of foliar absorbed N and remobilisation for spring growth in young nectarine (P. persica var nectarina) trees. Tree Physiol. 1998, 18, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Toselli, M.; Thalheimer, M.; Tagliavini, M. Leaf uptake and partitioning of urea-N as affected by the concentration and volume of spray solution and by the shoot leaf position in apple (Malus domestica) trees. J. Hortic. Sci. Biotechnol. 2004, 79, 97–100. [Google Scholar] [CrossRef]
- Dong, S.; Neilsen, D.; Neilsen, G.H.; Fuchigami, L.H. Foliar N application reduces soil NO3−-N leaching loss in apple orchards. Plant Soil 2005, 268, 357–366. [Google Scholar] [CrossRef]
- Tagliavini, M.; Toselli, M. Foliar application of nutrients. In Encyclopedia of Soils in the Environment; Hillel, D., Hatfield, J.L., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands; ISBN 0-12-348530-4.


{kind=link}
| Tree Species | Net N Removal (g N/kg Fresh Fruit) |
|---|---|
| Apple | 0.9 |
| Peach | 2.7 |
| Pear | 1.7 |
| Orange | 3.7 |
| Walnut | 10 |
| Olive | 22 |
| Kiwi | 4.5 |
| Organ | N Content (g/Tree) | |||
|---|---|---|---|---|
| Total N in the Organ | Ndff of the Year | Ndff of Previous Year | N from Other Sources | |
| New leaves | 29.2 | 10.8 | 3.9 | 14.5 |
| Old leaves | 3.2 | 0.8 | 0.6 | 1.8 |
| New branches | 7.3 | 2.9 | 1.0 | 3.4 |
| Old branches | 7.2 | 2.8 | 1.2 | 3.2 |
| Trunk | 4.0 | 1.2 | 0.9 | 1.9 |
| Fine roots | 4.6 | 2.0 | 0.8 | 1.8 |
| Old roots | 12.3 | 3.5 | 2.4 | 6.4 |
| Strategy | Mechanisms to Increase Tree N Use Efficiency |
|---|---|
| N fertilization practices | Split mineral N fertilizer rate into several applications possibly by fertigation; targeted supply of N in the soil volume explored by roots; application of N in the period of active root growth; promote internal N storage and remobilization also by foliar N applications; use organic N fertilizers. |
| Soil management | Promote the presence of legume plants among the orchard floor vegetation; use cover crops and incorporate into the soil their biomass; enhance the soil organic matter and the organic soil N, e.g., manure and compost, leave plant residues and mowed grasses on the orchard floor; promote presence of mycorrhizae that expand absorption ability and may allow organic N uptake. |
| Plant genotype | Choice of efficient plant genotypes with limited growth and high yield potential; rootstocks with a high root density, able to take up N at high rates; genotypes able to maximize photosynthesis with low leaf N. |
| Foliar N Fertilizer Supplied to | Example |
|---|---|
| prevent or treat temporary N deficiency | After remobilisation has finished in spring and root N uptake is still low; latter, during fruit set/maturation, in N deficient plants |
| overcome limiting conditions of N availability or uptake | Poor root growth, low soil temperature, low soil moisture, poor soil aeration |
| increase reserve accumulation for remobilisation in the following year | In late summer-autumn |
| apply N to deep-rooted trees, when broadcasting is almost ineffective | When the soil supplied N does not reach the soil layers explored by roots due to the absence of rainfall or irrigation |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carranca, C.; Brunetto, G.; Tagliavini, M. Nitrogen Nutrition of Fruit Trees to Reconcile Productivity and Environmental Concerns. Plants 2018, 7, 4. https://doi.org/10.3390/plants7010004
Carranca C, Brunetto G, Tagliavini M. Nitrogen Nutrition of Fruit Trees to Reconcile Productivity and Environmental Concerns. Plants. 2018; 7(1):4. https://doi.org/10.3390/plants7010004
Chicago/Turabian StyleCarranca, Corina, Gustavo Brunetto, and Massimo Tagliavini. 2018. "Nitrogen Nutrition of Fruit Trees to Reconcile Productivity and Environmental Concerns" Plants 7, no. 1: 4. https://doi.org/10.3390/plants7010004
APA StyleCarranca, C., Brunetto, G., & Tagliavini, M. (2018). Nitrogen Nutrition of Fruit Trees to Reconcile Productivity and Environmental Concerns. Plants, 7(1), 4. https://doi.org/10.3390/plants7010004