Effect of Silicon on the Tolerance of Wheat (Triticum aestivum L.) to Salt Stress at Different Growth Stages: Case Study for the Management of Irrigation Water

Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Parameters, Chlorophyll and Carotenoid Contents
2.2. Determination of Ions and Protein Contents
2.3. Estimation of the H2O2 Content
2.4. Enzyme Extraction and Activity Assay
2.5. Statistical Analysis
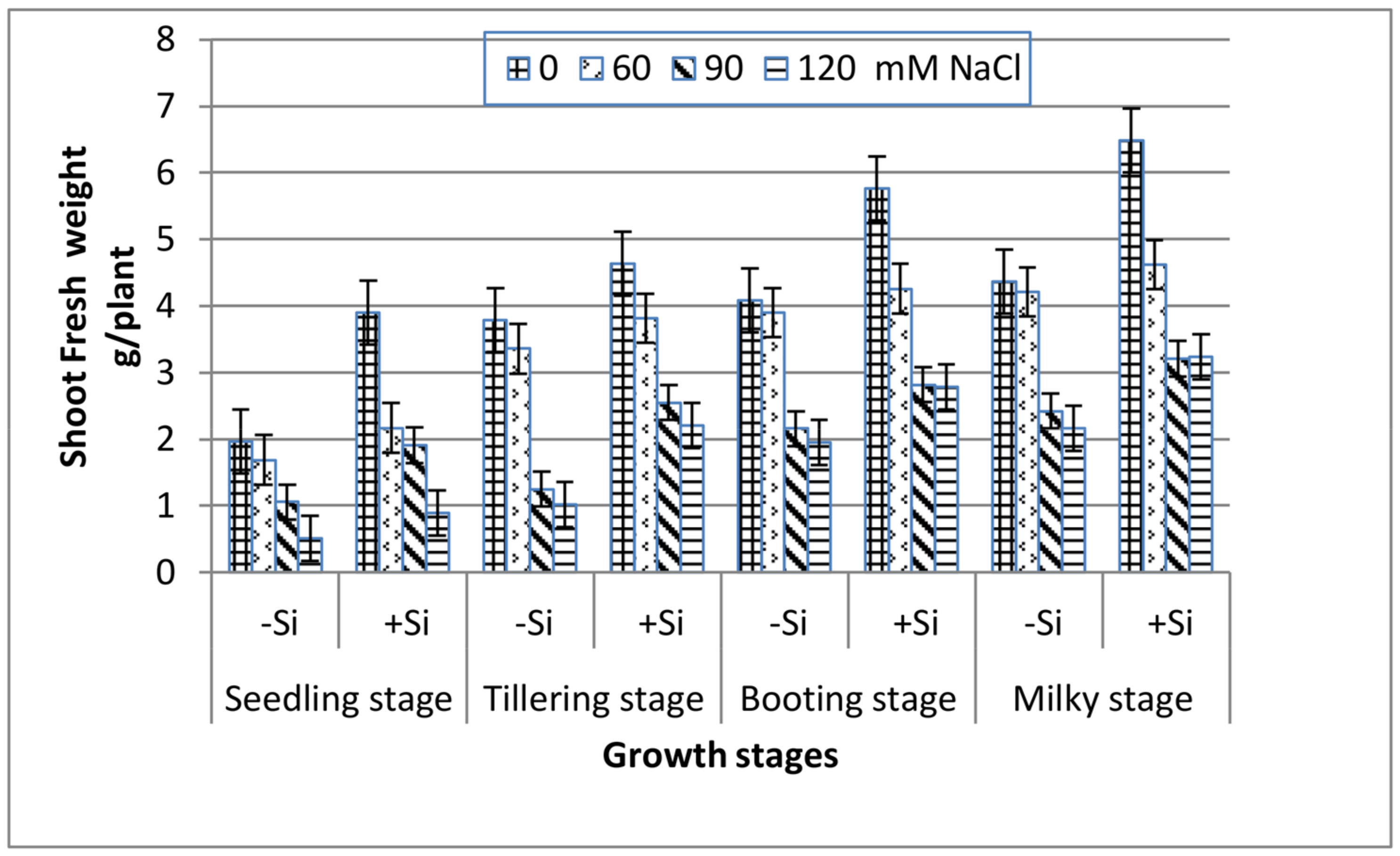
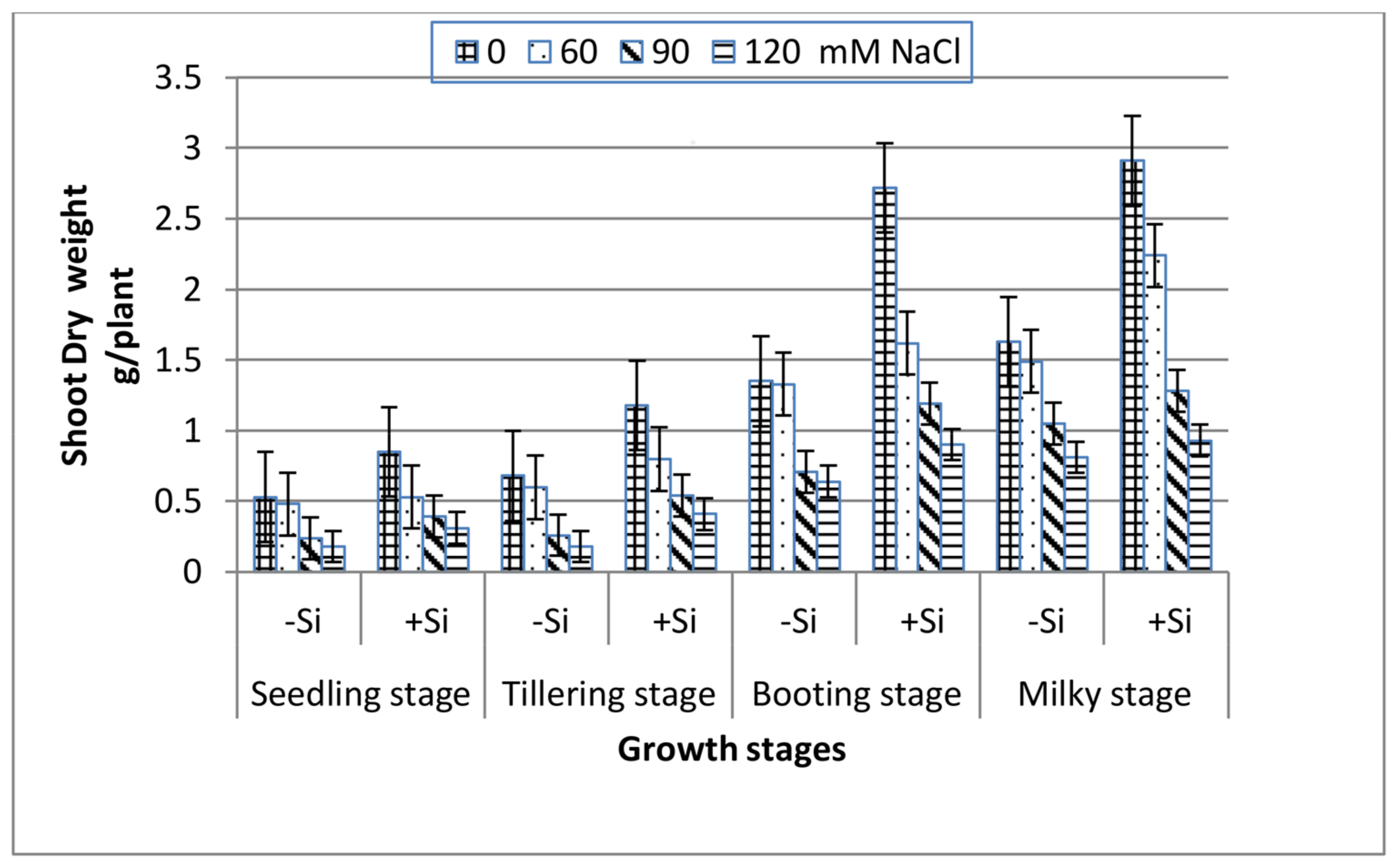
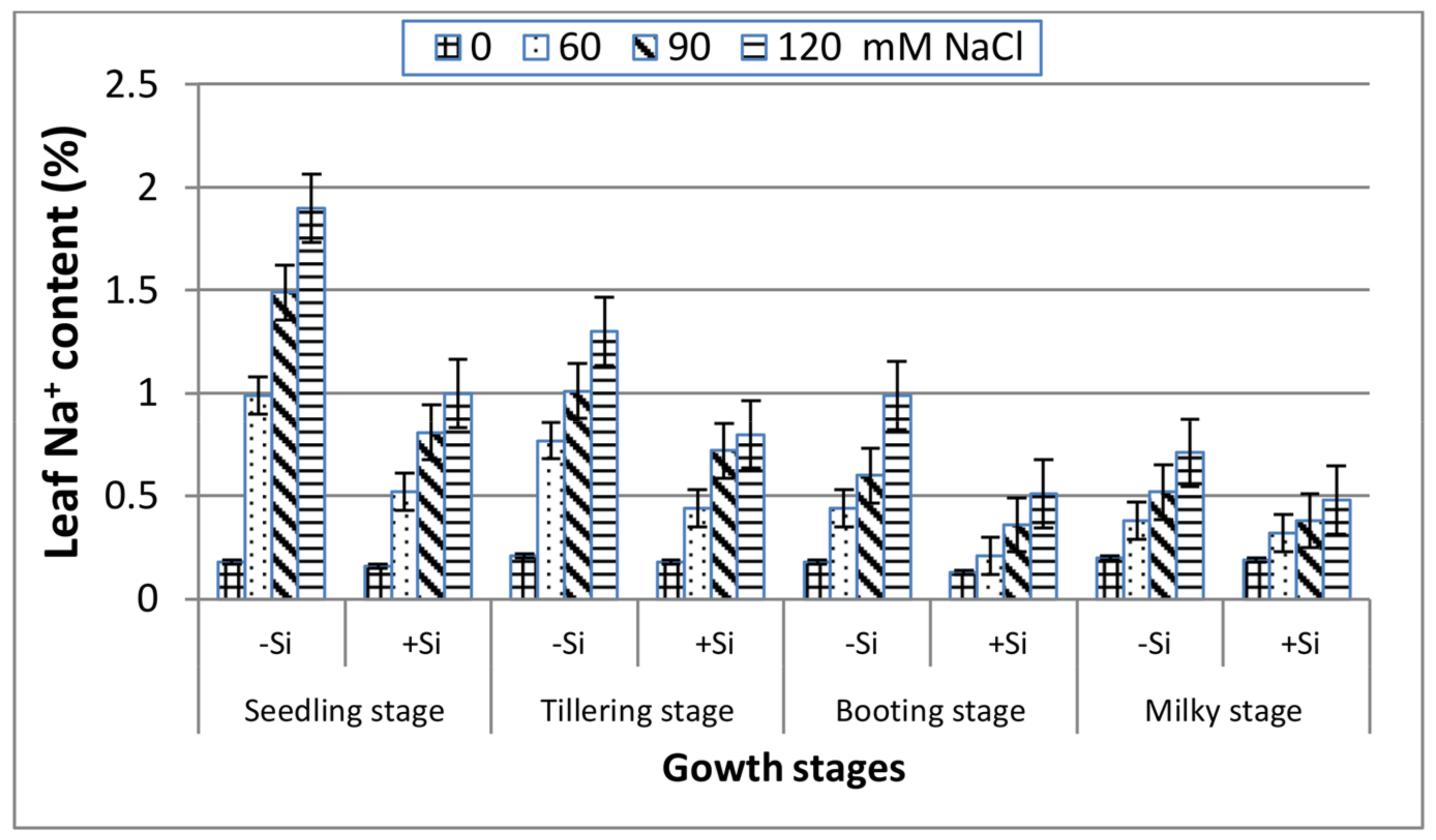
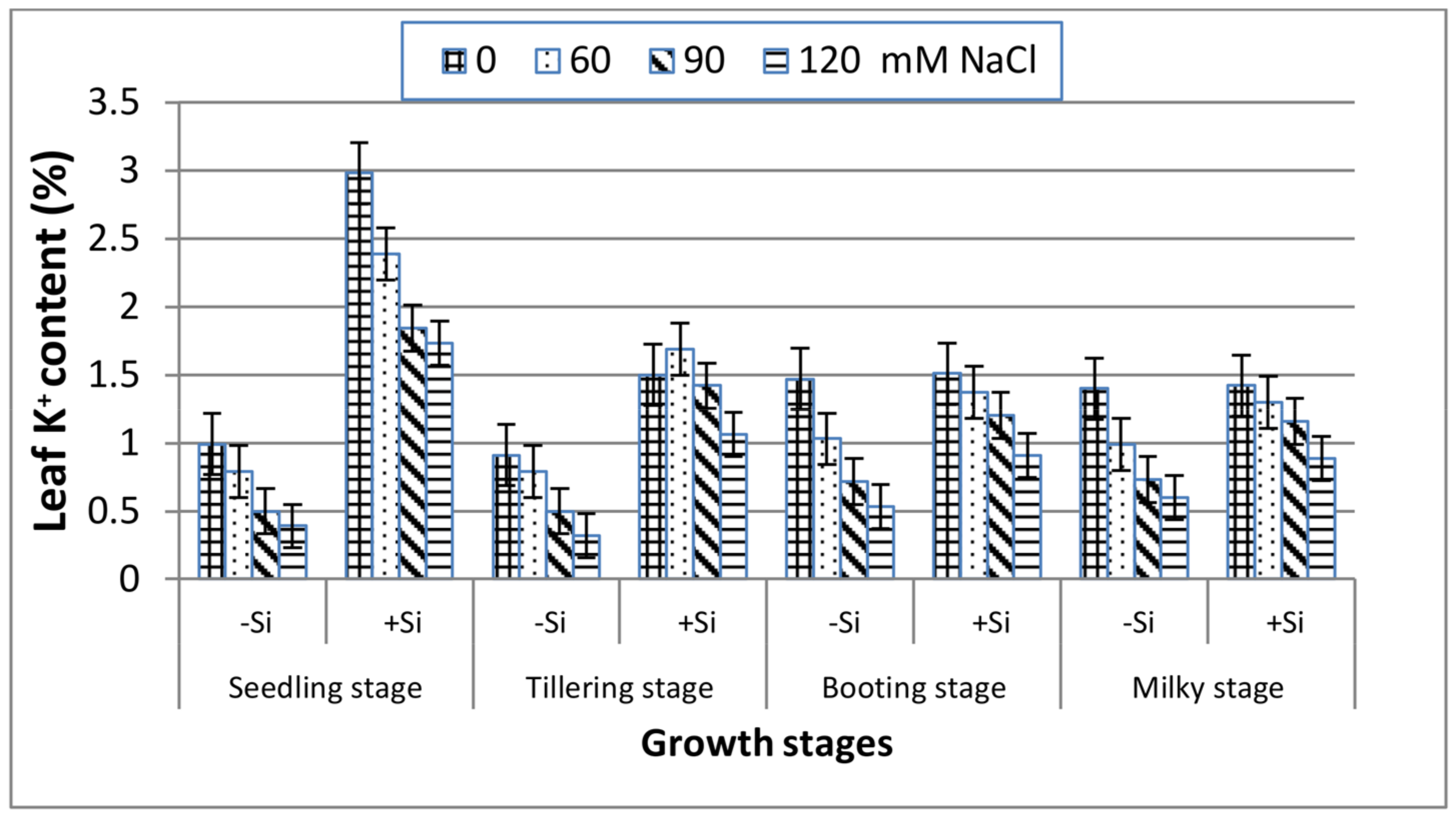
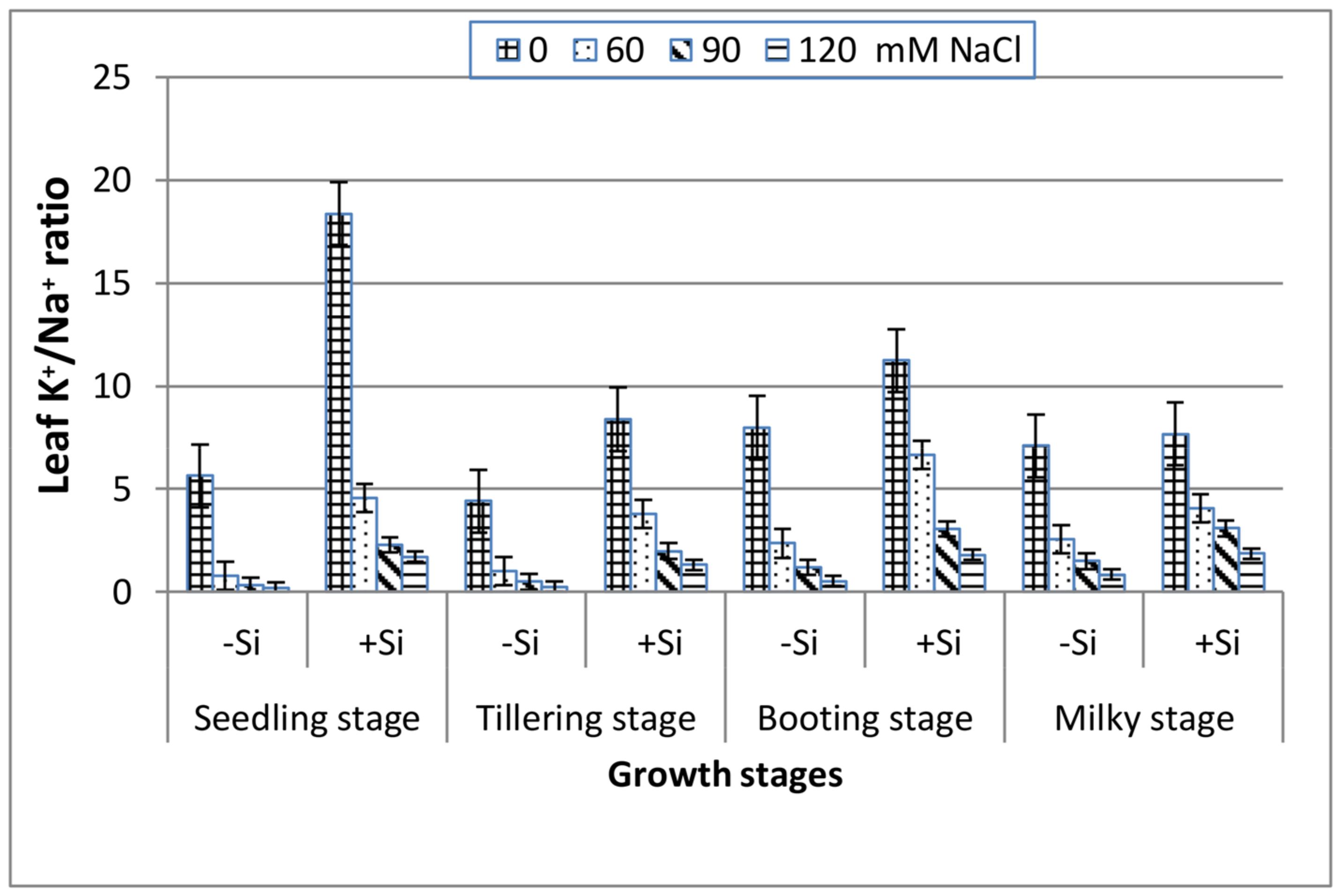
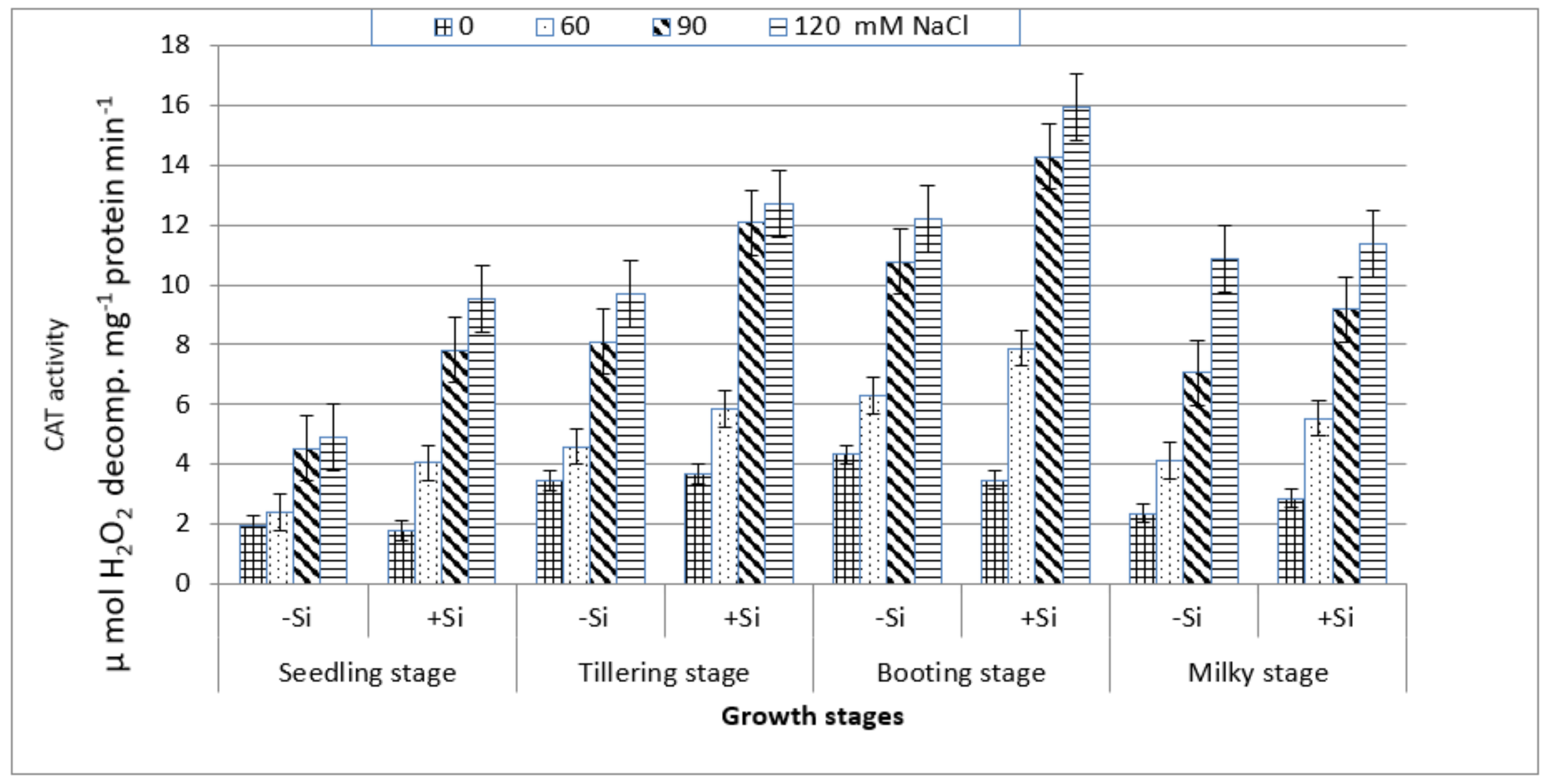
3. Results
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Ashraf, M.; Akram, N.A. Improving salinity tolerance of plants through conventional breading and genetic engineering: An analytical comparison. Biotechnol. Adv. 2009, 27, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Sairam, R.K.; Rao, K.V.; Srivastava, G.C. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical and molecular characterization. Int. J. Genom. 2014. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Song, R.; Shao, H.; Song, F.; Xu, H.; Lu, Y. Silicon improves maize photosynthesis in saline-alkaline soils. Sci. World J. 2015, 2015, 245072. [Google Scholar] [CrossRef] [PubMed]
- Younes, A.H.H.; Hasan, N.A.; Daoud, A.M. The effect of application time of saline water on wheat production. J. Agric. Sci. Mansoura Univ. 1995, 20, 1879–1884. [Google Scholar]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Antioxidant responses of wheat plants under stress. Genet. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Kao, C.H. Effect of NaCl stress on H2O2 metabolism in rice leaves. Plant Growth Regul. 2000, 30, 1151–1155. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Goery, M.; Shuleav, V.; Van Breusegen, F. ROS Signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Scandalios, J.G. Oxygen stress and superoxide dismutase. Plant Physiol. 1993, 101, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Akram, A.; Ashraf, M. Alleviation of adverse effects of salt stress on sunflower (Helionthus annus L.) by exogenous application of potassium nitrate. Appl. Bot. Food Qual. 2009, 83, 19–27. [Google Scholar]
- Pitman, M.G.; Läuchli, A. Global impact of salinity and agricultural ecosystem. In Salinity: Environment—Plant—Molecules; Lächli, A., Lüttge, U., Eds.; Kluwer: Dordrecht, The Netherlands, 2002; pp. 30–50. [Google Scholar]
- Ali, M.A.; Tahir, M.; Amin, S.; Basra, S.M.; Maqbool, M.; Lee, D.J. Si induced stress tolerance in wheat (Triticum aestivum L.) hydroponically grown under water deficit conditions. Bulg. J. Agric. Sci. 2013, 19, 951–957. [Google Scholar]
- Daoud, A.M. Yield response and mineral composition of wheat in relation to the applied silicon under saline environment. Alex. Sci. Exch. J. 2005, 26, 385–395. [Google Scholar]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Shim, J.K.; Kim, D.H.; Lee, K.Y.; Lee, I.J. Silicon application to rice root zone influenced the photo hormonal and antioxidant responses under salinity stress. J. Plant Growth Regul. 2014, 33, 137–149. [Google Scholar] [CrossRef]
- Daoud, A.M.; Fayed, R.I.; Mahmoud, A.M.; El-Zahaby, E.M. Impact of steal slag application on nutrients availability and corn yield grown on saline soil. J. Soil Sci. Agric. Eng. Mansoura Univ. 2013, 4, 777–791. [Google Scholar]
- Laing, Y.; Zhang, W.; Chen, Q.; Ding, R. Effect of silicon on H+-ATPase and H+-PPase activity, Fatty acid composition and fluidity of tonoplast vesicles from roots of salt-stressed barley (Hordeum vulgare L.). J. Environ. Exp. Bot. 2005, 53, 29–37. [Google Scholar] [CrossRef]
- Lee, S.K.; Sohn, E.Y.; Hamayun, M.; Moon, J.Y.; Lee, I.J. Effect of silicon on growth and salinity stress of soybean plant grown under hydroponic system. Agrofor. Syst. 2010, 80, 333–340. [Google Scholar] [CrossRef]
- Rizwan, M.; Meunier, J.D.; Miche, H.; Keller, C. Effect of silicon on reducing cadmium toxicity in durum wheat (Triticum turgidum L. cv. Claudio W.) grown in a soil with aged contamination. J. Hazard. Mater. 2012, 209, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Sommer, M.; Kaczorek, D.; Kuzyakov, Y.; Breuer, J. Silicon pools and fluxes in soils and landscapes. A review. J. Plant Nutr. Soil Sci. 2006, 169, 310–329. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Functions and transport of silicon in plants. Cell. Mol. Life Sci. 2008, 65, 3049–3059. [Google Scholar] [CrossRef] [PubMed]
- El-Ghonemy, A.A.; Wallace, A.; El-Gazzar, A.M.; Romney, E.M. Sodium relations in desert plants: 6. Variations in vegetation characteristics along a traces in the Qattara depression. Egypt Sci. 1982, 134, 57–64. [Google Scholar] [CrossRef]
- Ming, D.F.; Pei, Z.F.; Naeem, M.S.; Gong, H.J.; Zhou, W.J. Silicon alleviates PEG-induced water-deficit stress in upland rice seedling by enhancing osmotic adjustment. J. Agron. Crop Sci. 2012, 198, 14–26. [Google Scholar] [CrossRef]
- Ahmad, R.; Zaheer, S.H.; Ismail, S. Role of Silicon in salt tolerance of wheat (Triticum aestivum L.). Plant Sci. 1992, 85, 43–50. [Google Scholar] [CrossRef]
- Gurmani, A.R.; Bano, A.; Ullah, N.; Khan, H.; Jahangir, M.; Flowers, T.J. Exogenous abscisic acid (ABA) and silicon (Si) promote salinity tolerance by reducing sodium (Na+) transport and bypass flow in rice (Oryza sativa indica). Aust. J. Crop Sci. 2013, 7, 1219–1226. [Google Scholar]
- Yeo, A.R.; Flowers, S.A.; Rao, G.; Welfare, K.; Senanayake, N.; Flowers, T.J. Silicon reduces sodium uptake in rice (Oryza sativa L.) in saline conditions and this is accounted for by a reduction in transpirational bypass flow. Plant Cell Environ. 1999, 22, 559–565. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Baharwan, S.A.; Zia-ur-Rehman, M.; Qayyum, F.M.; Abbas, F. Mechanisms of silicon—Mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef] [PubMed]
- Helal, R.M. Influence of exogenous application of silicon on physiological response of salt-stressed maize (Zea mays L.). Int. J. Agric. Biol. 2006, 8, 293–297. [Google Scholar]
- Laing, Y.; Chen, Q.; Lui, Q.; Zhang, W.; Ding, R. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in root of salt-stressed barley (Hordeum vulgare L.). J. Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef]
- Farshidi, M.; Abdolzadeh, A.; Sadeghipour, H.R. Silicon nutrition alleviates physiological disorders imposed by salinity in hydroponically grown canola (Brassica napus L.) plants. Acta Physiol. Plant. 2012, 34, 1779–1788. [Google Scholar] [CrossRef]
- Rafi, M.M.; Epstein, E. Silicon absorption by wheat (Triticum aestivum L.). Plant Soil 1999, 211, 223–230. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Bloom, P.R. Extinction coefficients of chlorophyll a and b in N,N-Dimethylformamide and 80% acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H. Chlorophyll and Carotenoid: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Cottenie, A.; Verloo, M.; Kiekens, L.; Velghe, G.; Camerlynck, R. Chemical Analysis of Plants and Soils; Laboratory of Analytical and Agrochemistry, State University: Ghent, Belgium, 1982. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Ann. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Valeria, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant system in acid rain treated bean plants: Protective role of exogenous polyamine. Plant Sci. 2000, 151, 59–66. [Google Scholar]
- Esfandiari, E.; Shakiba, M.R.; Mahboob, S.A.; Alyari, H.; Toorshi, M. Water stress, antioxidant enzyme activity and lipid peroxidation in wheat seedling. J. Food Agric. Environ. 2007, 5, 149–153. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics, 2nd ed.; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Liu, P.; Yin, L.; Wang, S.; Zhang, M.; Deng, X.; Zhang, S.; Tanaka, K. Enhanced root hydraulic conductance by aquaporin regulation accounts for silicon alleviated salt-induced osmotic stress in Sorghum bicolor L. Environ. Exp. Bot. 2015, 111, 42–51. [Google Scholar] [CrossRef]
- Takemura, T.; Hanagata, N.; Sugihara, K.; Baba, S.; Karube, I.; Dubinski, Z. Physiological and biochemical responses to salt stress in the mangrove, Bruguiera gymnorrliza. Aquat. Bot. 2000, 68, 15–28. [Google Scholar] [CrossRef]
- Magnaye, M.A.; Santos, P.J.A.; Cruz, P.C.S. Responses of yellow corn (Zea mays L.) inbreds to salinity. Asia Life Sci. 2011, 20, 521–533. [Google Scholar]
- Grattan, S.R.; Grieve, C.M. Salinity-mineral nutrient relations in horticultural crops. Sci. Hortic. 1999, 78, 127–157. [Google Scholar] [CrossRef]
- Bybordi, A. Interactive effects of silicon and potassium nitrate in improving salt tolerance of wheat. Int. J. Agric. 2004, 13, 1889–1899. [Google Scholar]
- Haghighi, M.; Afifipour, Z.; Mozafarian, M. The alleviation effect of silicon on seed germination and seedling growth of tomato under salinity stress. Veg. Crops Res. Bull. 2012, 76, 119–126. [Google Scholar] [CrossRef]
- Yin, L.; Wang, S.; Li, J.; Tanaka, K.; Oka, M. Application of silicon improves salt tolerance through ameliorating osmotic and ionic stresses in the seedling of Sorghum bicolor. Acta Physiol. Plant. 2013, 35, 3099–3107. [Google Scholar] [CrossRef]
- Khoshgoftarmanesh, A.H.; Khodarahmi, S.; Haghighi, M. Effect of silicon nutrition on lipid peroxidation and antioxidant response of cucumber plants exposed to salinity stress. Arch. Agron. Soil Sci. 2014, 60, 639–653. [Google Scholar] [CrossRef]
- Sun, Y.; Luo, W.; Zhang, W.; Ziume, X. Effects of exogenous silicon on germination characteristics of cucumber seeds under NaHCO3 stress. In Proceedings of the 2010 International Conference on Challenges in Environmental Science and Computer Engineering (CESCE), Wuhan, China, 6–7 March 2010; pp. 471–474. [Google Scholar]
- Barakat, N.; Laudadio, V.; Cazzato, E.; Tufarelli, V. Antioxidant potential and oxidative stress markers in wheat (Triticum aestivum L.) treated with phyto hormones under salt-stress conditions. Int. J. Agric. Biol. 2013, 15, 843–849. [Google Scholar]
- Singh, A.C.; Dubey, R.S. Changes in chlorophyll a and b contents and activities of photosystem 1 and 2 in rice seedlings induced by NaCl. Photosynthetica 1995, 31, 489–499. [Google Scholar]
- Al-Aghabary, K.; Zhu, Z.; Shi, Q. Influence of silicon supply on chlorophyll content, chlorophyll fluorescence and anti-oxidative enzyme activities in tomato plants under salt stress. J. Plant Nutr. 2004, 27, 2101–2115. [Google Scholar] [CrossRef]
- Hashemi, A.; Abdolzadeh, A.; Sadeghipour, H. Beneficial effects of silicon nutrition in alleviating salinity stress in hydroponically grown canola, Brassica napas L. plants. Soil Sci. Plant Nutr. 2010, 56, 244–253. [Google Scholar] [CrossRef]
- Farida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotexicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar]
- Li, Q.; Niu, H.; Yin, J.; Wang, M.; Shao, H.; Deng, D.; Chen, X.; Rend, J.; Li, Y. Protective role of exogenous nitric oxide against oxidative-stress induced by salt stress in barley (Hordeum vulgare). Colloids Surf. B Biointerfaces 2008, 65, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Seong, E.S.; Cho, H.S.; Choi, D.; Joung, Y.H.; Lim, C.K.; Hur, J.H.; Wang, M.H. Tomato plants over expression CaKRI enhanced tolerance to salt stress and oxidative stress. Biochem. Biophys. Res. Commun. 2007, 363, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.J.; Chen, K.M.; Zhao, Z.G.; Chen, G.C.; Zhou, W.J. Effects of silicon on defense of wheat against oxidative stress under drought at different developmental stages. Biol. Plant. 2008, 52, 592–596. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration |
|---|---|
| KNO3 | 3 mM |
| Ca(NO3)2 | 2 mM |
| NH4H2PO4 | 0.5 mM |
| MgSO4 | 0.5 mM |
| KCl | 25 µM |
| H3BO3 | 12.5 µM |
| MnSO4·H2O | 1 µM |
| ZnSO4·7H2O | 1 µM |
| H2MO4 | 0.25 µM |
| CuSO4·5H2O | 0.25 µM |
| NiSO4·6H2O | 0.1 µM |
| Fe-EDTA | 100 µM |
| Treatments | Seedling Stage | Tillering Stage | Booting Stage | Milky Stage | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NaCl mM | Si mM | Chl.a | Chl.b | Cart. | Total | Chl.a | Chl.b | Cart. | Total | Chl.a | Chl.b | Cart. | Total | Chl.a | Chl.b | Cart. | Total |
| 0 | 0 | 9.77 | 6.72 | 4.09 | 20.58 | 14.55 | 9.76 | 6.63 | 30.9 | 16.31 | 8.73 | 8.73 | 33.77 | 10.01 | 6.07 | 12.66 | 28.74 |
| 60 | 0 | 8.91 | 6.52 | 3.91 | 19.34 | 12.61 | 9.17 | 7.96 | 29.7 | 14.52 | 10.65 | 8.21 | 33.38 | 8.93 | 4.31 | 14.31 | 27.55 |
| 90 | 0 | 6.62 | 5.33 | 3.16 | 15.11 | 10.92 | 8.33 | 8.23 | 27.5 | 9.09 | 6.95 | 9.95 | 25.99 | 6.71 | 4.06 | 11.92 | 22.69 |
| 120 | 0 | 4.08 | 4.91 | 3.7 | 12.69 | 7.08 | 6.04 | 8.91 | 22.0 | 6.13 | 4.91 | 8.91 | 19.95 | 4.09 | 1.71 | 16.97 | 20.77 |
| 0 | 0.78 | 9.82 | 6.96 | 3.21 | 19.99 | 15.22 | 10.2 | 6.69 | 32.1 | 16.95 | 12.24 | 7.91 | 37.10 | 9.52 | 6.71 | 11.91 | 28.14 |
| 60 | 0.78 | 8.76 | 6.71 | 3.62 | 19.09 | 15.18 | 9.76 | 7.21 | 32.2 | 15.67 | 11.52 | 7.67 | 34.86 | 10.69 | 6.03 | 11.36 | 28.08 |
| 90 | 0.78 | 8.31 | 6.01 | 3.55 | 17.87 | 13.23 | 8.99 | 7.76 | 30.0 | 13.05 | 8.36 | 7.51 | 28.92 | 7.71 | 5.88 | 12.71 | 26.30 |
| 120 | 0.78 | 6.21 | 4.99 | 4.09 | 15.19 | 10.51 | 8.12 | 7.09 | 25.7 | 9.17 | 6.11 | 8.64 | 23.92 | 5.12 | 4.61 | 12.74 | 22.47 |
| LSD 0.05 | 1.09 | 1.12 | 2.18 | 4.35 | 2.58 | 1.93 | 1.18 | 2.26 | 2.71 | 1.80 | 2.24 | 5.19 | 1.49 | 2.38 | 2.19 | 4.49 | |
| NaCl mM | Si mM | Seedling Stage | Tillering Stage | Booting Stage | Milky Stage |
|---|---|---|---|---|---|
| 0 | 0 | 10.1 | 13.5 | 19.9 | 19.8 |
| 60 | 0 | 18.7 | 22.8 | 16.7 | 18.9 |
| 90 | 0 | 31.1 | 50.6 | 66.4 | 88.1 |
| 120 | 0 | 16.9 | 73.5 | 89.7 | 119.7 |
| 0 | 0.78 | 9.9 | 16.8 | 17.1 | 17.3 |
| 60 | 0.78 | 12.5 | 18.4 | 8.0 | 9.8 |
| 90 | 0.78 | 22.1 | 34.9 | 53.9 | 48.9 |
| 120 | 0.78 | 38.5 | 54.2 | 60.3 | 79.4 |
| LSD 0.05 | 4.81 | 9.11 | 8.93 | 7.50 | |
| Treatment | Seedling Stage | Tillering Stage | Booting Stage | Milky Stage | |||||
|---|---|---|---|---|---|---|---|---|---|
| NaCl mM | Si mM | T.P. | SOD | T.P. | SOD | T.P. | SOD | T.P. | SOD |
| 0 | 0 | 87.1± 7.3 | 0.81± 0.06 | 102.9 ± 8.6 | 3.34± 0.24 | 127.9±10.7 | 6.09± 0.44 | 136.3 ± 11.4 | 2.50±0.18 |
| 60 | 0 | 58.2 ± 6.5 | 3.43 ± 0.38 | 83.2±9.2 | 5.62± 0.62 | 99.1 ± 11.0 | 8.11± 0.90 | 102.1 ± 11.3 | 4.43 ± 0.49 |
| 90 | 0 | 40.0 ± 4.0 | 5.05 ± 0.42 | 60.9 ± 6.1 | 9.25 ± 0.77 | 74.1 ± 7.4 | 13.0 ± 1.15 | 83.1 ± 8.3 | 11.65 ± 0.97 |
| 120 | 0 | 115.0 ± 8.2 | 7.21 ± 0.56 | 133.8 ±9.6 | 8.45 ± 0.65 | 148.5 ± 10.6 | 16.90 ± 1.30 | 154.4 ± 11.0 | 17.21 ± 1.32 |
| 0 | 0.78 | 97.0 ± 10.8 | 0.84 ± 0.08 | 128.1 ± 14.2 | 3.03 ± 0.09 | 136.9 ± 15.2 | 5.44 ± 0.22 | 148.1 ± 16.5 | 1.91 ± 0.08 |
| 60 | 0.78 | 69.6 ± 8.0 | 6.21 ± 0.52 | 94.1 ± 7.8 | 8.81 ± 0.73 | 110.1 ± 9.2 | 10.31 ± 0.86 | 113.3 ± 9.4 | 9.85 ± 0.82 |
| 90 | 0.78 | 51.1 ± 3.9 | 7.85 ± 0.87 | 75.2 ±5.8 | 13.65 ± 1.52 | 83.6 ± 6.4 | 18.09 ± 2.01 | 87.4± 6.7 | 20.41 ± 2.27 |
| 120 | 0.78 | 91.2 ± 8.3 | 9.47 ± 0.95 | 116.5 ± 10.6 | 19.77 ± 1.98 | 139.7 ± 12.7 | 22.24 ± 2.22 | 143.2 ± 13.0 | 27.97 ± 2.80 |
| LSD 0.05 | - | 2.02 | - | 2.62 | - | 4.91 | - | 6.88 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
A.M., D.; M.M., H.; N., S.; A.A., E.-A.; L., M. Effect of Silicon on the Tolerance of Wheat (Triticum aestivum L.) to Salt Stress at Different Growth Stages: Case Study for the Management of Irrigation Water. Plants 2018, 7, 29. https://doi.org/10.3390/plants7020029
A.M. D, M.M. H, N. S, A.A. E-A, L. M. Effect of Silicon on the Tolerance of Wheat (Triticum aestivum L.) to Salt Stress at Different Growth Stages: Case Study for the Management of Irrigation Water. Plants. 2018; 7(2):29. https://doi.org/10.3390/plants7020029
Chicago/Turabian StyleA.M., Daoud, Hemada M.M., Saber N., El-Araby A.A., and Moussa L. 2018. "Effect of Silicon on the Tolerance of Wheat (Triticum aestivum L.) to Salt Stress at Different Growth Stages: Case Study for the Management of Irrigation Water" Plants 7, no. 2: 29. https://doi.org/10.3390/plants7020029
APA StyleA.M., D., M.M., H., N., S., A.A., E. -A., & L., M. (2018). Effect of Silicon on the Tolerance of Wheat (Triticum aestivum L.) to Salt Stress at Different Growth Stages: Case Study for the Management of Irrigation Water. Plants, 7(2), 29. https://doi.org/10.3390/plants7020029