Transcriptome and Metabolome Analyses Reveal That Nitrate Strongly Promotes Nitrogen and Carbon Metabolism in Soybean Roots, but Tends to Repress It in Nodules

Abstract
:1. Introduction
2. Results
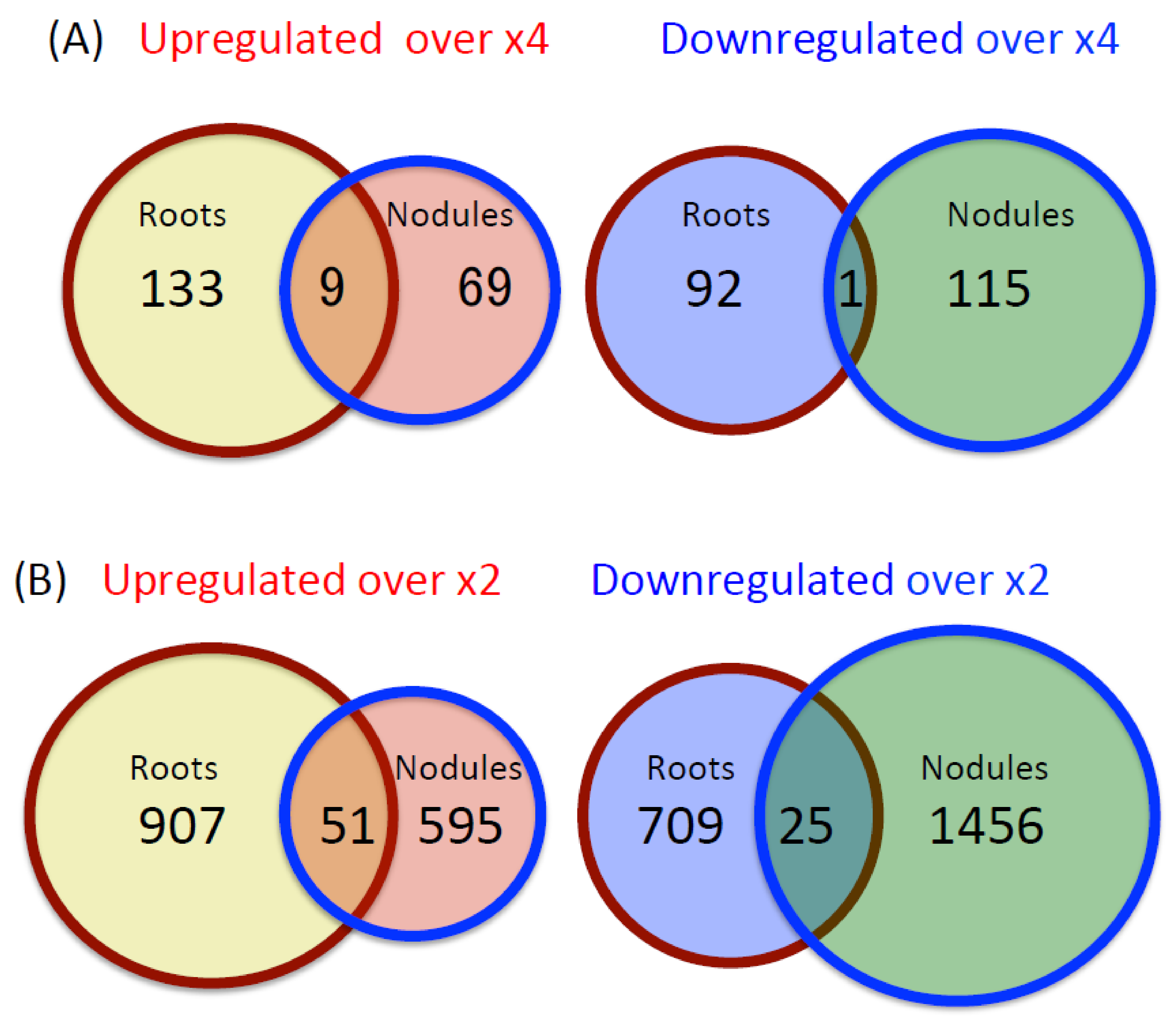
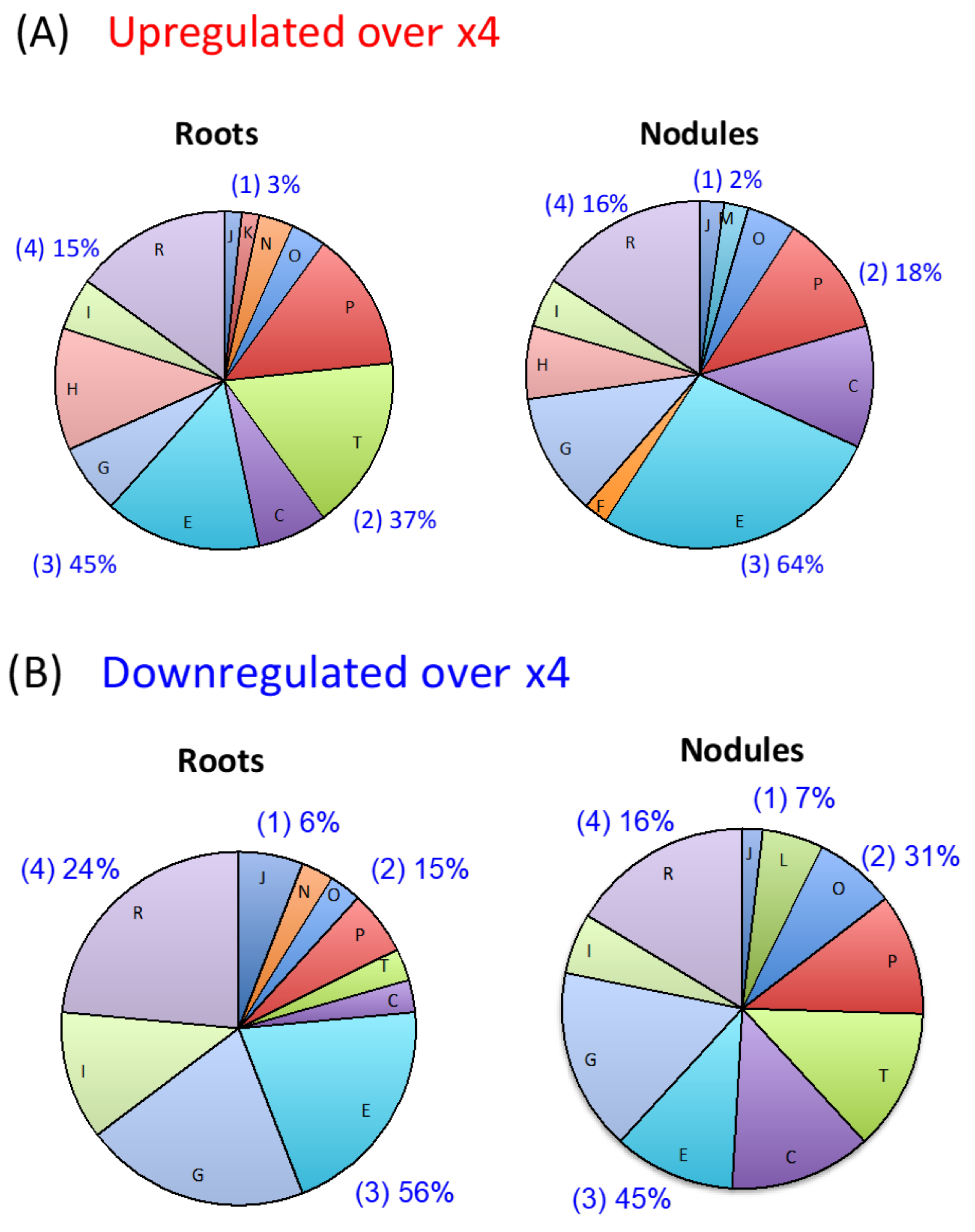
2.1. Number and Function of Upregulated and Downregulated Genes in Roots and Nodules
2.2. Upregulation and Downregulation of Root-Specific and Nodule-Specific Genes by the Nitrate Treatment
2.3. Upregulation and Downregulation of Leghemoglobin and Nodulin Genes by the Nitrate Treatment
2.4. Upregulation and Downergulation of Phytohormone-Related Genes by the Nitrate Treatment
2.5. Metabolome Analysis of Roots and Nodules Comparing the Nitrate Treatment and Control
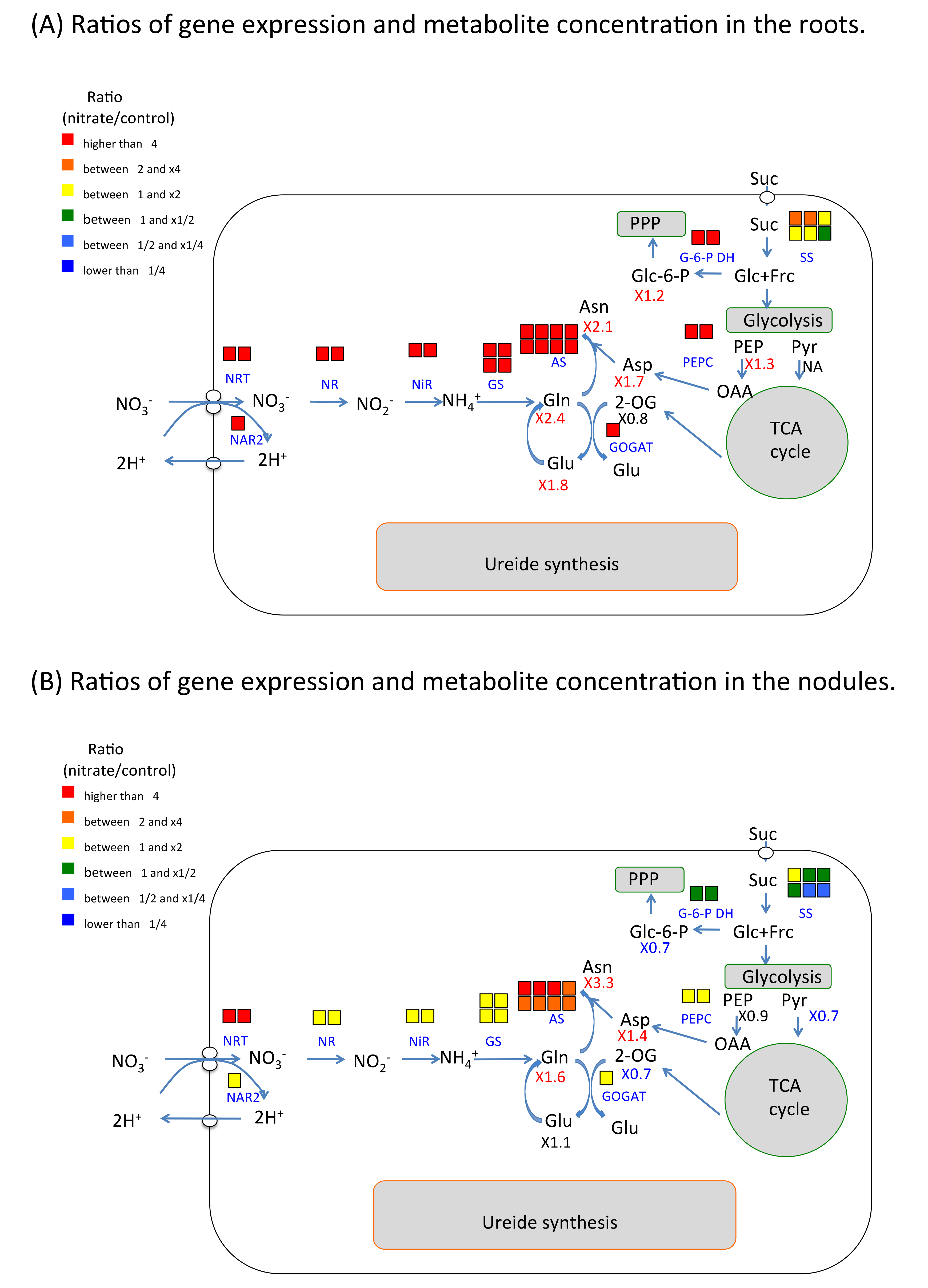
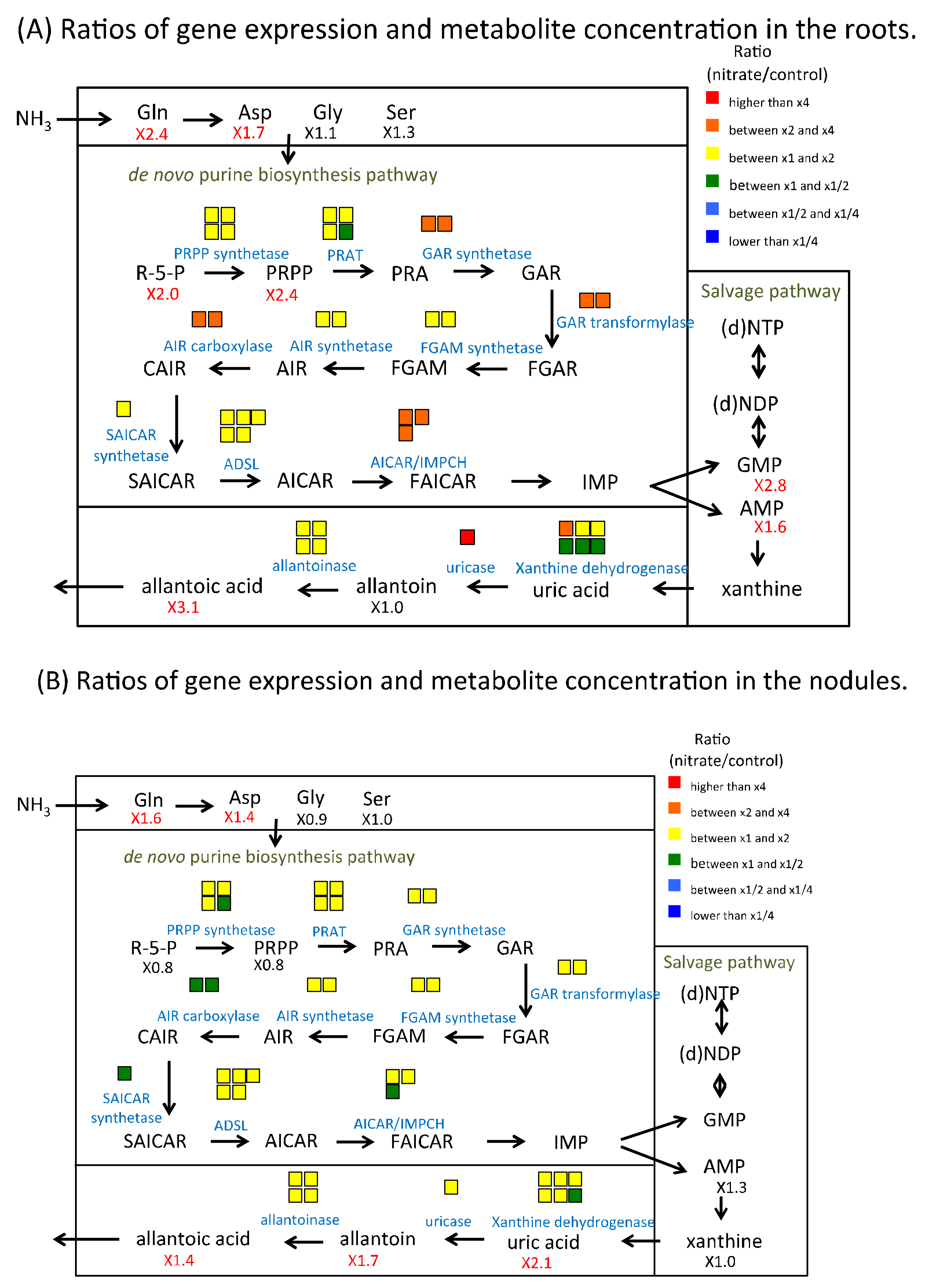
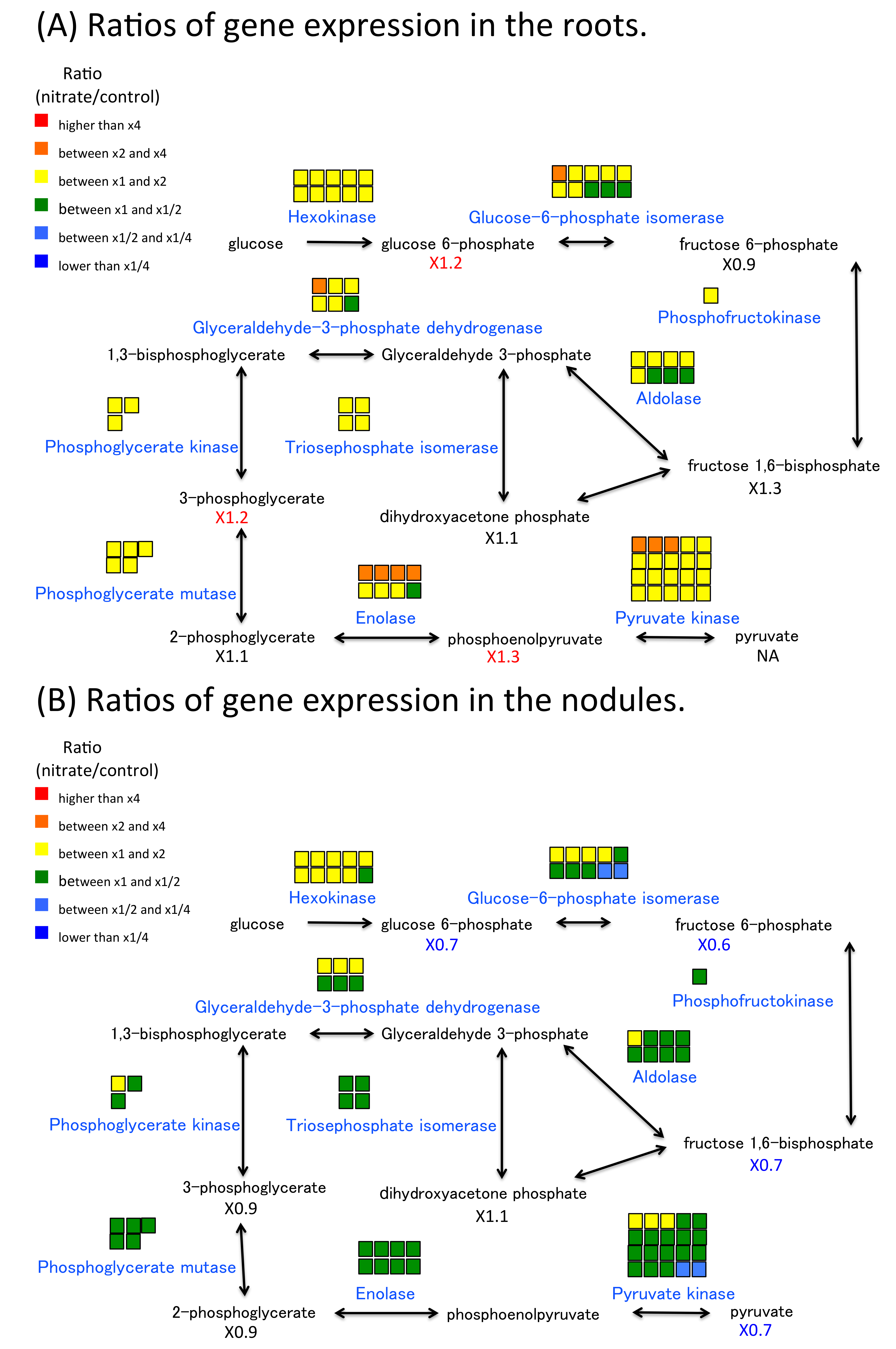
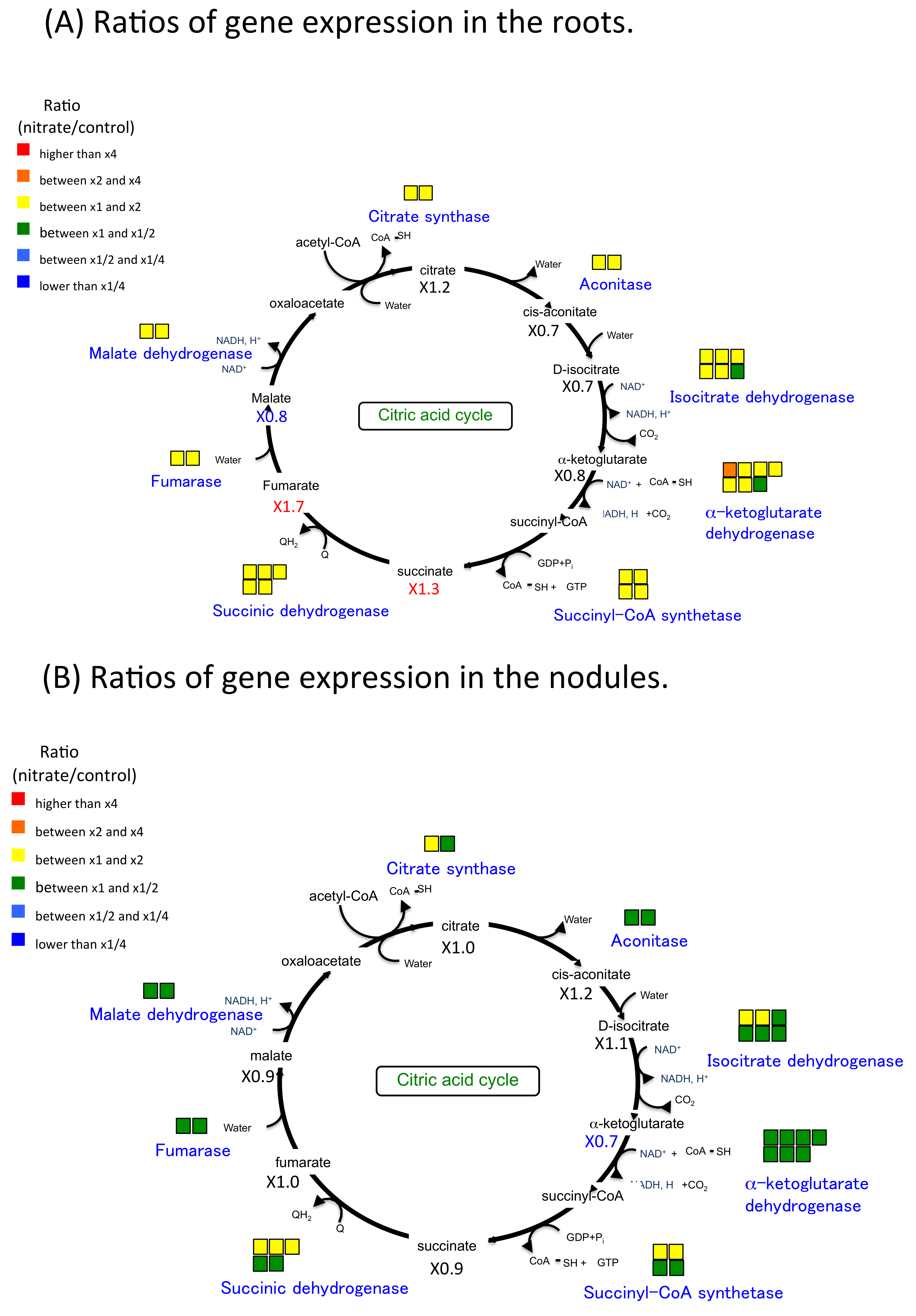
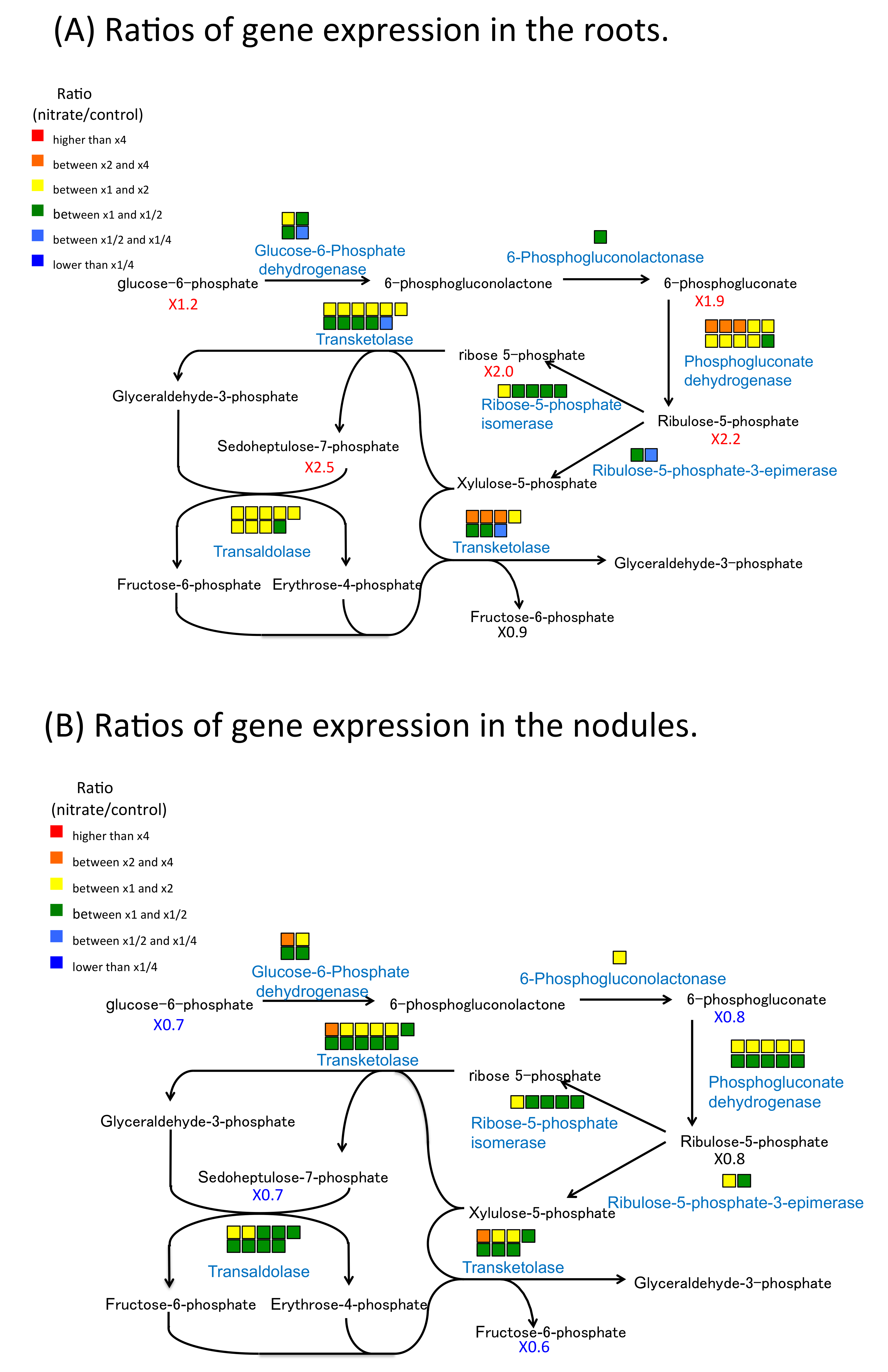
2.6. Nitrate Effect of Gene Expression and Metabolite Concentration on Nitrogen and Carbon Metabolism in Roots and Nodules
3. Discussion
3.1. The Effect of Nitrate on Nitrate Absorption and Assimilation and Carbon Metabolism
3.2. The Effect of Nitrate on Asparagine and Ureide Metabolism
3.3. The Effect of Nitrate Treatment on Nodulin and Hormone Related Probesets
4. Materials and Methods
4.1. Plant Growth and the Nitrate Treatment
4.2. Transcriptome Analysis
4.3. Metabolome Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AICAR | 5-aminoimidazole-4-carboxamide ribonucleotide |
| AIR | 5-aminoimidazole ribotide |
| AON | autoregulation of nodulation |
| AS | asparagine synthetase |
| Asn | asparagine |
| Asp | aspartate |
| CAIR | 5′-phosphoribosyl-4-carboxy-5-aminoimidazole |
| FAICAR | 5-formamidoimidazole-4-carboxamide ribotide |
| FGAM | 5′-phosphoribosylformylglycinamideine |
| FGAR | phosphoribosyl-N-formylglycineamide |
| Frc | fructose |
| GAR | glycineamide ribonucleotide |
| G-6-P DH | glucose-6-phosphate dehydrogenase |
| Gln | glutamine |
| Glu | glutamate |
| GMP | guanosine 5′-monophosphate |
| GOGAT | glutamate synthase |
| GS | glutamine synthetase |
| IMP | inosinic acid |
| NAR2 | high-affinity nitrate transporter-activating protein |
| (d)NDP | (deoxy) nucleotidediphosphate |
| NR | nitrate reductase |
| NRT | nitrate transporter |
| (d)NTP | (deoxy) nucleotidetriphosphate |
| OAA | oxaloacetate |
| 2-OG | 2-oxoglutarate |
| PEP | phosphoenolpyruvate |
| PEPC | phosphoenol pyruvate carboxylase |
| PPP | pentose phosphate pathway |
| R-5-P | ribose-5-phosphate |
| PRPP | 5-phosphoribosyl 1-diphosphate |
| PRA | 5-phosphoribosylamine |
| Pyr | pyruvate |
| SAICAR | phosphoribosylaminoimidazolesuccinocarboxyamide |
| SS | sucrose synthase |
| Suc | sucrose |
References
- Ohyama, T.; Minagawa, R.; Ishikawa, S.; Yamamoto, M.; Hung, N.V.P.; Ohtake, N.; Sueyoshi, K.; Sato, T.; Nagumo, Y.; Takahashi, Y. Soybean Seed Production and Nitrogen Nutrition. In A Comprehensive Survey of International Soybean Research-Genetics, Physiology, Agronomy and Nitrogen Relationships; Board, J.E., Ed.; InTech: Rijeka, Croatia, 2013; pp. 115–158. [Google Scholar] [CrossRef]
- Harper, J.E. Soil and symbiotic nitrogen requirement for optimum soybean production. Crop Sci. 1974, 14, 255–260. [Google Scholar] [CrossRef]
- Gibson, A.H.; Harper, J.E. Nitrate effect on nodulation of soybean by Bradyrhizobium japonicum. Crop Sci. 1985, 25, 497–501. [Google Scholar] [CrossRef]
- Streeter, J. Inhibition of legume nodule formation and N2 fixation by nitrate. Crit. Rev. Plant Sci. 1988, 7, 1–23. [Google Scholar] [CrossRef]
- Ohyama, T.; Fujikake, H.; Yashima, H.; Tanabata, S.; Ishikawa, S.; Sato, T.; Nishiwaki, T.; Ohtake, N.; Sueyoshi, K.; Ishii, S.; et al. Effect of Nitrate on Nodulation and Nitrogen Fixation of soybean. In Soybean Physiology and Biology; El-Shemy, H.A., Ed.; InTech: Rijeka, Croatia, 2012; pp. 333–364. [Google Scholar] [CrossRef]
- Harper, J.E.; Gibson, A.H. Differential nodulation tolerance to nitrate among legume species. Crop Sci. 1984, 24, 797–801. [Google Scholar] [CrossRef]
- Davidson, I.A.; Robson, M.J. Effect of contrasting patterns of nitrate application on the nitrate uptake, N2-fixation, nodulation and growth of white clover. Ann. Bot. 1986, 57, 331–338. [Google Scholar] [CrossRef]
- Harper, J.E. Nitrogen metabolism. In Soybeans: Improvement, Production, and Uses, 2nd ed.; Wilcox, J.R., Ed.; Agronomy Monograph No.16; American Society of Agronomy: Madison, WI, USA, 1987; pp. 497–533. [Google Scholar]
- Mizukoshi, K.; Nishiwaki, T.; Ohtake, N.; Minagawa, R.; Ikarashi, T.; Ohyama, T. Nitrate transport pathway into soybean nodules traced by tungstate and 15NO3−. Soil Sci. Plant Nutr. 1995, 41, 75–88. [Google Scholar] [CrossRef]
- Fujikake, H.; Yamazaki, A.; Ohtake, N.; Sueyoshi, K.; Matsuhashi, S.; Ito, T.; Mizuniwa, C.; Kume, T.; Hashimoto, S.; Ishioka, N.S.; et al. Quick and reversible inhibition of soybean root nodule growth by nitrate involves a decrease in sucrose supply to nodules. J. Exp. Bot. 2003, 54, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Fujikake, H.; Tamura, Y.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Photoassimilate partitioning in hypernodulation mutant of soybean (Glycine max (L.) Merr.) NOD1-3 and its parent Williams in relation to nitrate inhibition of nodule growth. Soil Sci. Plant Nutr. 2003, 49, 583–590. [Google Scholar] [CrossRef]
- Yashima, H.; Fujikake, H.; Sato, T.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Systemic and local effects of long-term application of nitrate on nodule growth and N2 fixation in soybean (Glycine max (L.) Merr.). Soil Sci. Plant Nutr. 2003, 49, 825–834. [Google Scholar] [CrossRef]
- Yashima, H.; Fujikake, H.; Yamazaki, A.; Ito, S.; Sato, T.; Tewari, K.; Ohtake, N.; Sueyoshi, K.; Takahashi, Y.; Ohyama, T. Long-term effect of nitrate application from lower part of roots on nodulation and N2 fixation in upper part of roots of soybean (Glycine max (L.) Merr.) in two-layered pot experiment. Soil Sci. Plant Nutr. 2005, 51, 981–990. [Google Scholar] [CrossRef]
- Carroll, B.J.; Mathews, A. Nitrate inhibition of nodulation in legumes. In Molecular Biology of Symbiotic Nitrogen Fixation; Gresshoff, P.M., Ed.; CRC Press: Boca Raton, FL, USA, 1990; pp. 159–180. [Google Scholar]
- Vessey, J.K.; Walsh, K.B; Layzell, D.B. Can a limitation in phloem supply to nodules account for the inhibitory effect of nitrate on nitrogenase activity? Physiol. Plant. 1988, 74, 137–146. [Google Scholar] [CrossRef]
- Thorpe, M.R.; Walsh, K.B.; Minchin, P.E.H. Photoassimilate partitioning in nodulated soybean I. 11C methodology. J. Exp. Bot. 1998, 49, 1805–1815. [Google Scholar] [CrossRef]
- Neo, H.H.; Layzell, D.B. Phloem glutamine and the regulation of O2 diffusion in legume nodules. Plant Physiol. 1997, 113, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Bacanamwo, M.; Harper, J.E. The feedback mechanism of nitrate inhibition of nitrogenase activity in soybean may involve asparagine and/or products of its metabolism. Physiol. Plant. 1997, 100, 371–377. [Google Scholar] [CrossRef]
- Gordon, A.J.; Skøt, L.; James, C.L.; Minchin, F.R. Short-term metabolic responses of soybean root nodules to nitrate. J. Exp. Bot. 2002, 53, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Kanahama, K.; Kanayama, Y. Involvement of nitric oxide in the inhibition of nitrogenase activity by nitrate in Lotus root nodules. J. Plant Physiol. 2010, 167, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Fujikake, H.; Yashima, H.; Sato, T.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Rapid and reversible nitrate inhibition of nodule growth and N2 fixation activity in soybean (Glycine max (L.) Merr.). Soil Sci. Plant Nutr. 2002, 48, 211–217. [Google Scholar] [CrossRef]
- Saito, A.; Tanabata, S.; Tanabata, T.; Tajima, S.; Ueno, M.; Ishikawa, S.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Effect of nitrate on nodule and root growth of soybean (Glycine max (L.) Merr.). Int. J. Mol. Sci. 2014, 15, 4464–4480. [Google Scholar] [CrossRef] [PubMed]
- Tanabata, S.; Tanabata, T.; Saito, A.; Tajima, S.; Watanabe, S.; Ishikawa, K.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Computational image analysis method for measuring size of nodule growth in soybean. Jpn. J. Soil Sci. Plant Nutr. 2014, 85, 43–47. [Google Scholar]
- Ruffel, S.; Freixes, S.; Balzergue, S.; Tillard, P.; Jeudy, C.; Martin-Magniette, M.L.; van der Merwe, M.J.; Kakar, K.; Gouzy, J.; Fernie, A.R.; et al. Systemic Signaling of the Plant Nitrogen Status Triggers Specific Transcriptome Responses Depending on the Nitrogen Source in Medicago truncatula. Plant Physiol. 2008, 146, 2020–2035. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R.; Koester, B.; Liese, R.; Lingner, A.; Baumgarten, V.; Dirks, J.; Salinas-Riester, G.; Pommerenke, C.; Dittert, K.; Schulze, J. An RNA sequencing transcriptome analysis reveals novel insights into molecular aspects of the nitrate impact on the nodule activity of Medicago truncatula. Plant Physiol. 2014, 164, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Caetano-Anolles, G.; Gresshoff, P.M. Plant genetic control of nodulation. Ann. Rev. Microbiol. 1991, 45, 345–382. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.E.; Ferguson, B.J.; Hayashi, S.; Lin, Y.H.; Gresshoff, P.M. Molecular mechanisms controlling legume autoregulation of nodulation. Ann. Bot. 2011, 108, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Soyano, T.; Kawaguchi, M. Systemic Regulation of Root Nodule Formation. In Advances in Biology and Ecology of Nitrogen Fixation; Ohyama, T., Ed.; InTech: Rijeka, Croatia, 2014; pp. 73–88. [Google Scholar] [CrossRef]
- Delves, A.C.; Mathews, A.; Day, D.A.; Carter, A.S.; Carroll, B.J.; Gresshoff, P.M. Regulation of the soybean-Rhizobium nodule symbiosis by shoot and root factors. Plant Physiol. 1986, 82, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Gremaud, M.F.; Harper, J.E. Selection and initial characterization of partially nitrate tolerant nodulation mutants of soybean. Plant Physiol. 1989, 89, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Akao, S.; Kouchi, H. A supernodulating mutant isolated from soybean cultivar Enrei. Soil Sci. Plant Nutr. 1992, 38, 183–187. [Google Scholar] [CrossRef]
- Ohyama, T.; Nicholas, J.C.; Harper, J.E. Assimilation of 15N2 and 15NO3− by partially nitrate-tolerant nodulation mutants of soybean. J. Exp. Bot. 1993, 44, 1739–1747. [Google Scholar] [CrossRef]
- Okamoto, S.; Shinohara, H.; Mori, T.; Matsubayashi, Y.; Kawaguchi, M. Root-derived CLE glycopeptides control nodulation by direct binding to HAR1 receptor kinase. Nat. Commun. 2013. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, T.; Sakurai, T.; Totoki, Y.; Toyoda, A.; Seki, M.; Ishiwata, A.; Akiyama, K.; Kurotani, A.; Yoshida, T.; Mochida, K.; et al. Sequencing and analysis of approximately 40,000 soybean cDNA clones from a fulllength-enriched cDNA library. DNA Res. 2008, 15, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Hirsch, A.M. Studying early nodulin gene ENOD40 expression and induction by nodulation factor and cytokinin in transgenic alfalfa. Plant Physiol. 1998, 116, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Nagata, M.; Suzuki, A. Effect of phytohormones on nodulation and nitrogen fixation in legumes plants. In Advances in Biology and Ecology of Nitrogen Fixation; Ohyama, T., Ed.; InTech: Rijeka, Croatia, 2014; pp. 111–128. [Google Scholar] [CrossRef]
- Gordon, A.J.; Minchin, F.R.; James, C.L.; Komina, O. Sucrose synthase in legume nodules is essential for nitrogen fixation. Plant Physiol. 1999, 120, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, T.; Kumazawa, K. Assimilation and transport of nitrogenous compounds originated from 15N2 fixation and 15NO3 abosorption. Soil. Sci. Plant Nutr. 1979, 25, 9–19. [Google Scholar] [CrossRef]
- Bojsen, K.; Abildsten, D.; Jensen, E.O.; Paludan, K.; Marcker, K.A. The chromosomal arrangement of six soybean leghemoglobin genes. EMBO J. 1983, 2, 1165–1168. [Google Scholar] [PubMed]
- Nishiwaki, T.; Sato, T.; Yashima, H.; Ikarashi, T.; Ohyama, T.; Harper, J.E.; Akao, S.; Kouchi, H. Changes in concentration of leghemoglobin components in hypernodulation mutants of soybean. Soil Sci. Plant Nutr. 1997, 43, 1091–1096. [Google Scholar] [CrossRef]
- Kouch, H.; Hata, S. Isolation and characterization of novel nodulin cDNAs representing genes expressed at early stages of soybean nodule development. Mol. Gen. Genet. 1993, 238, 106–119. [Google Scholar]
- Soga, T.; Heiger, D.N. Amino acid analysis by capillary electrophoresis electrospray ionization mass spectrometry. Anal. Chem. 2000, 72, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer–specific profiles. Metabolomics 2010, 6, 78–95. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nitrogen Compound | Roots | Nodules |
|---|---|---|
| Allantoic acid | 3.1 *** | 1.4 * |
| Glutamine | 2.4 * | 1.6 * |
| Asparagine | 2.1 * | 3.3 |
| Alanine | 2.0 *** | 0.9 |
| Homoserine | 2.8 * | 0.8 |
| 2-Aminobutyric acid | 1.9 ** | 0.8 * |
| Glutamic acid | 1.8 ** | 1.1 |
| Aspartic acid | 1.8 * | 1.4 * |
| Allantoin | 1.0 | 1.9 ** |
| N-Acetylglucosamine | 1.1 | 3.6 * |
| Phosphorous Compound | Roots | Nodules |
|---|---|---|
| ATP | 3.9 ** | 0.7 |
| ADP | 2.9 * | 0.9 |
| GMP | 2.8 ** | N.A. |
| CDP | 2.4 | 0.8 |
| UDP | 2.1 * | 0.9 |
| CTP | 2.1 * | N.A. |
| Sedoheptulose 7-phosphate | 2.5 ** | 0.7 * |
| Ribulose 5-phosphate | 2.2 *** | 0.8 |
| NAD+ | 2.2 * | 0.9 |
| Ribose 5-phosphate | 2.0 *** | 0.8 |
| 6-Phosphogluconic acid | 1.9 *** | 0.8 ** |
| Spermidine | 1.8 * | 1.2 |
| O-Acetylserine | 1.7 * | N.A. |
| AMP | 1.6 * | 1.3 |
| UMP | 1.6 * | 0.8 |
| CMP | 1.5 * | 1.0 |
| UDP-glucuronic acid | 1.4 * | 0.7 |
| Phosphoenolpyruvic acid | 1.3 * | 0.9 |
| Glucose 1-phosphate | 1.2 * | 0.8 * |
| Glucose 6-phosphate | 1.2 * | 1.0 |
| 3-Phosphoglyceric acid | 1.2 * | 0.9 |
| Organic Acid | Roots | Nodules |
|---|---|---|
| 2-Isopropylmalic acid | 2.4 * | 1.0 |
| 2-Aminoadipic acid | 2.1 ** | 1.1 |
| N-Acetylglutamic acid | 1.9 ** | 1.1 |
| Citramalic acid | 1.9 ** | 0.9 |
| Fumaric acid | 1.7 *** | 1.0 |
| Argininosuccinic acid | 1.4 ** | 1.3 * |
| Succinic acid | 1.3 * | 0.9 |
| Phosphoenolpyruvic acid | 1.3 * | 0.9 |
| Malic acid | 0.8 * | 0.9 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishikawa, S.; Ono, Y.; Ohtake, N.; Sueyoshi, K.; Tanabata, S.; Ohyama, T. Transcriptome and Metabolome Analyses Reveal That Nitrate Strongly Promotes Nitrogen and Carbon Metabolism in Soybean Roots, but Tends to Repress It in Nodules. Plants 2018, 7, 32. https://doi.org/10.3390/plants7020032
Ishikawa S, Ono Y, Ohtake N, Sueyoshi K, Tanabata S, Ohyama T. Transcriptome and Metabolome Analyses Reveal That Nitrate Strongly Promotes Nitrogen and Carbon Metabolism in Soybean Roots, but Tends to Repress It in Nodules. Plants. 2018; 7(2):32. https://doi.org/10.3390/plants7020032
Chicago/Turabian StyleIshikawa, Shinji, Yuki Ono, Norikuni Ohtake, Kuni Sueyoshi, Sayuri Tanabata, and Takuji Ohyama. 2018. "Transcriptome and Metabolome Analyses Reveal That Nitrate Strongly Promotes Nitrogen and Carbon Metabolism in Soybean Roots, but Tends to Repress It in Nodules" Plants 7, no. 2: 32. https://doi.org/10.3390/plants7020032
APA StyleIshikawa, S., Ono, Y., Ohtake, N., Sueyoshi, K., Tanabata, S., & Ohyama, T. (2018). Transcriptome and Metabolome Analyses Reveal That Nitrate Strongly Promotes Nitrogen and Carbon Metabolism in Soybean Roots, but Tends to Repress It in Nodules. Plants, 7(2), 32. https://doi.org/10.3390/plants7020032