TUNEL Assay and DAPI Staining Revealed Few Alterations of Cellular Morphology in Naturally and Artificially Aged Seeds of Cultivated Flax



Abstract
:1. Introduction
2. Results
2.1. Seed Viability under NA and AA Treatments
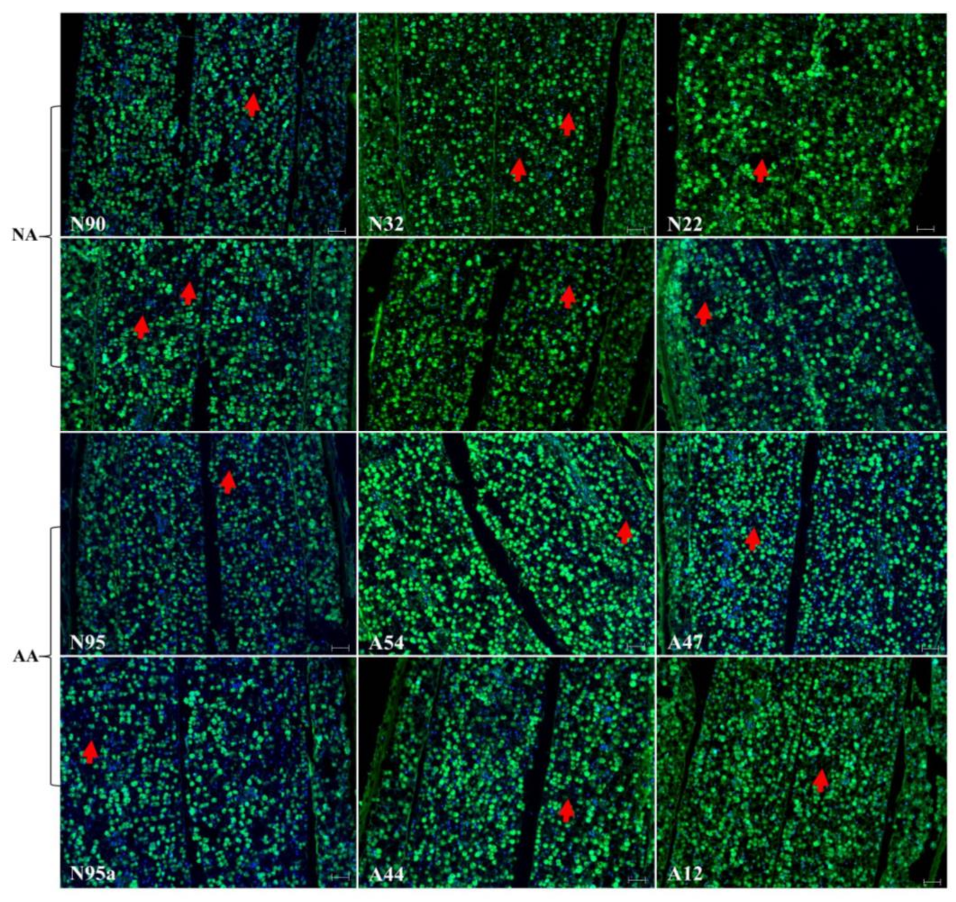
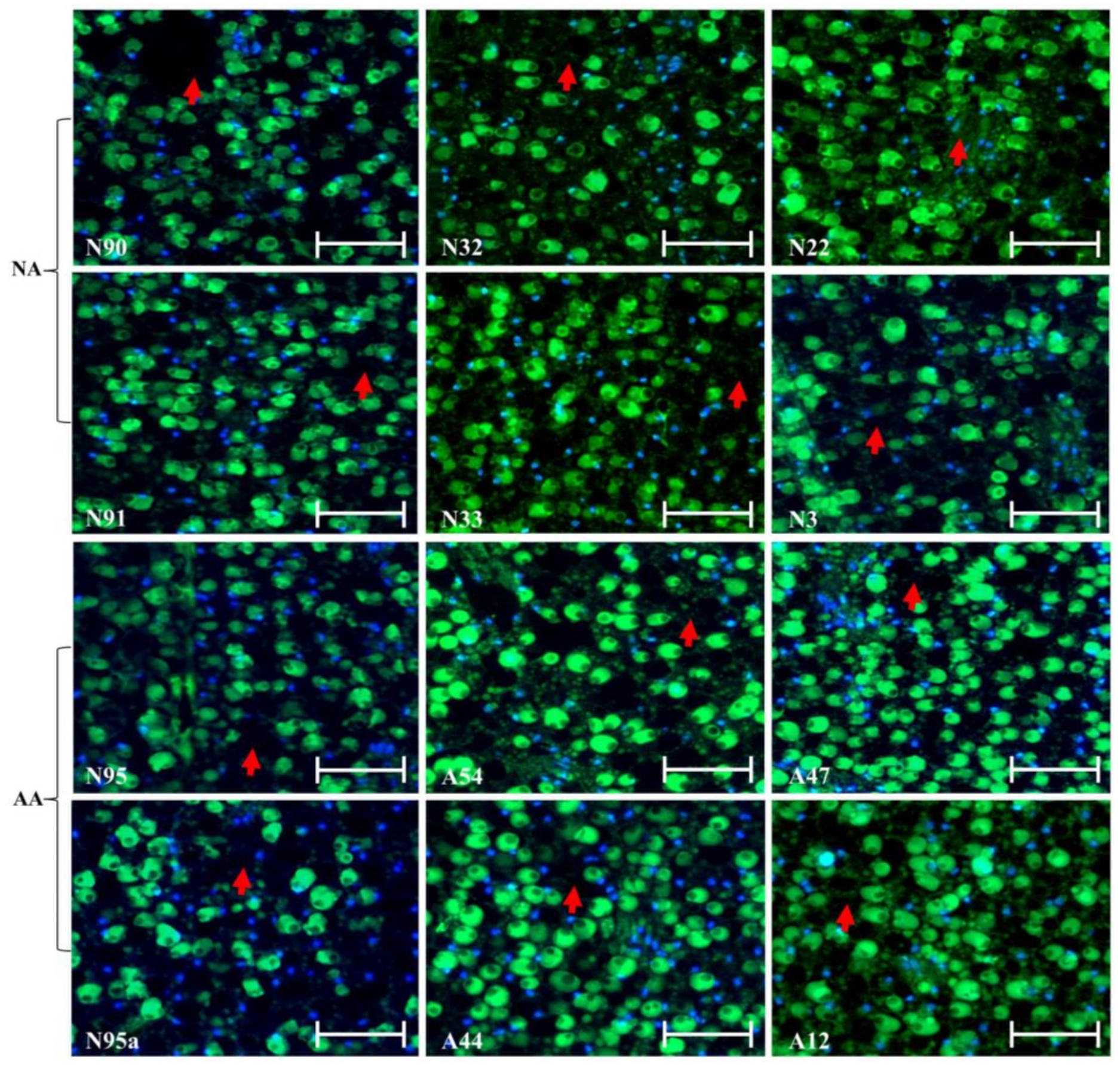
2.2. Assessment of Cellular Alterations Following NA and AA Treatments
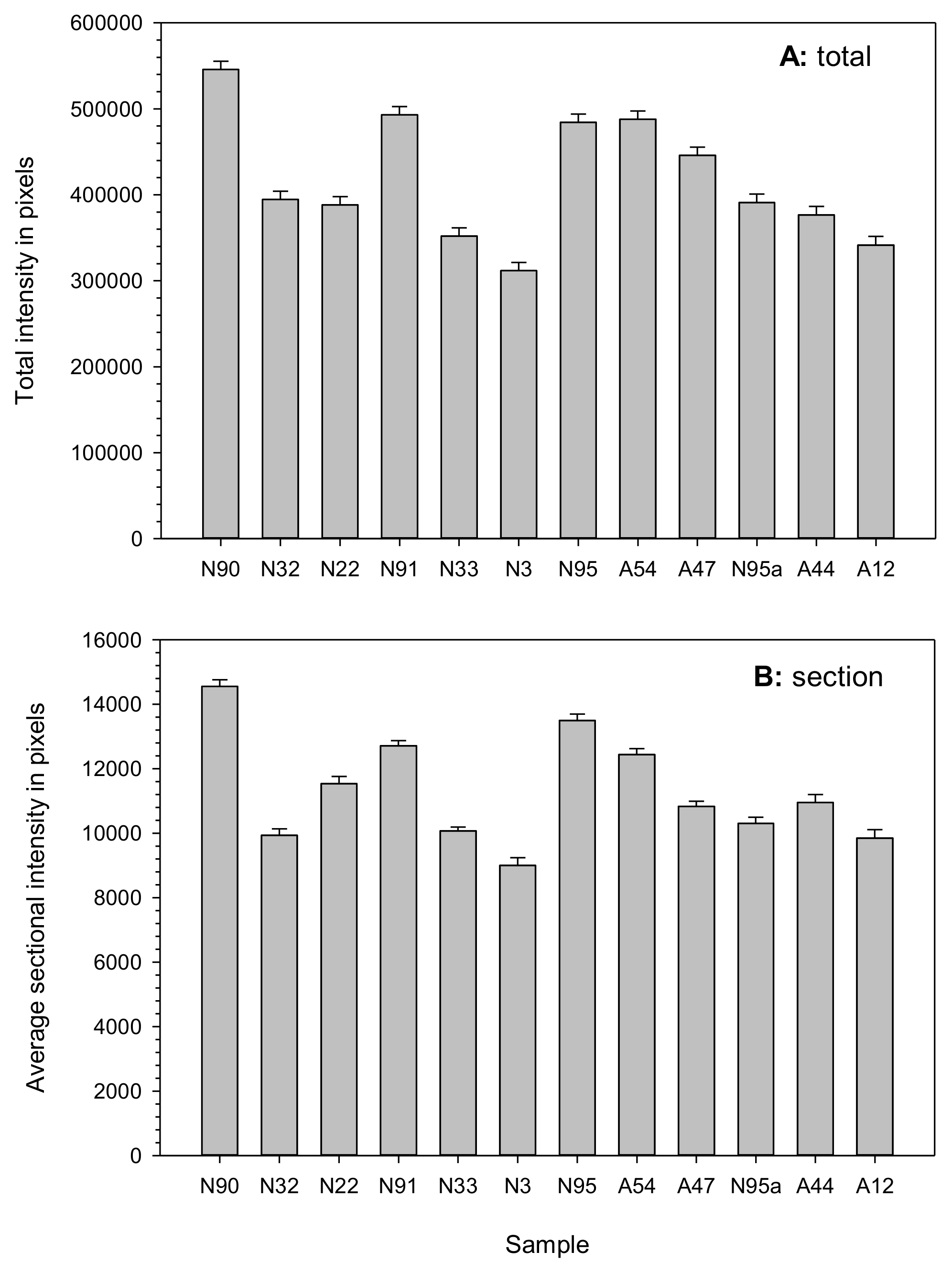
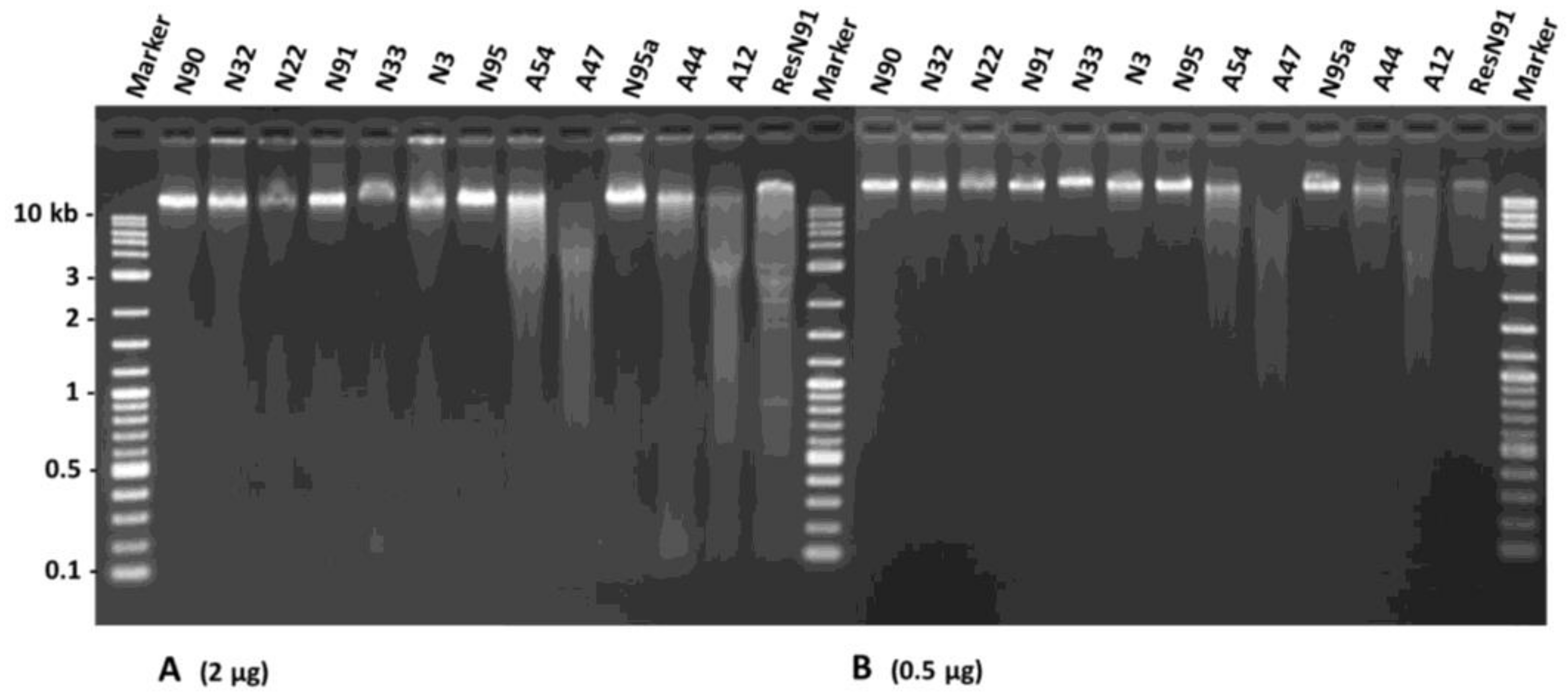
2.3. DNA Assessment of Nuclear Alterations
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Seed Fixation, Processing, and Tissue Sectioning
4.3. TUNEL Assay, DAPI Staining, and Microscopy
4.4. Analysis of Micrographs
4.5. Assessment of DNA Degradation
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fu, Y.B.; Ahmed, Z.; Diederichsen, A. Towards a better monitoring of seed ageing under ex situ seed conservation. Conserv. Physiol. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.D.; Dickie, J.D.; Linington, S.H.; Pritchard, H.W.; Probert, R.J. Seed Conservation: Turning Science into Practice 2003; Royal Botanic Gardens, Kew: London, UK, 2014. [Google Scholar]
- FAO. Draft Genebank Standards for Plant Genetic Resources for Food and Agriculture. Available online: http://www.fao.org/docrep/meeting/027/mf804e.pdf (accessed on 23 March 2018).
- Priestley, D.A. Seed Aging; Cornell University Press: Ithaca, NY, USA, 1986; pp. 39–197. [Google Scholar]
- Coolbear, P. Mechanism of seed deterioration. In Seed Quality: Basic Mechanisms and Agricultural Implications; Basra, A.S., Ed.; Food Product Press: New York, NY, USA, 1995; pp. 223–277. [Google Scholar]
- Cheah, K.S.E.; Osborne, D.J. DNA lesions occur with loss of viability in embryos of ageing rye seeds. Nature 1978, 272, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.T.; Berjak, P. Deteriorative changes associated with the loss of viability of stored desiccation-tolerant and desiccation-sensitive seeds. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 701–746. [Google Scholar]
- Walters, C. Understanding the mechanisms and kinetics of seed aging. Seed Sci. Res. 1998, 8, 223–244. [Google Scholar] [CrossRef]
- McDonald, M.B. Seed deterioration: Physiology, repair and assessment. Seed Sci. Technol. 1999, 27, 177–237. [Google Scholar]
- Chaudhury, A.M.; Koltunow, A.; Payne, T.; Luo, M.; Tucker, M.R.; Dennis, E.S.; Peacock, W.J. Control of early seed development. Annu. Rev. Cell Dev. Biol. 2001, 17, 677–699. [Google Scholar] [CrossRef] [PubMed]
- Kranner, I.; Minibayeva, F.V.; Beckett, R.P.; Seal, C.E. What is stress? Concepts, definitions and applications in seed science. New Phytol. 2010, 188, 655–673. [Google Scholar] [CrossRef] [PubMed]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Ramos, J.M.; Lopez, S.; Vazquez, E.; Murillo, E. DNA integrity and DNA polymerase activity in deteriorated maize embryo axes. J. Plant Physiol. 1988, 133, 600–604. [Google Scholar] [CrossRef]
- Corbineau, F.; Gay-Mathieu, C.; Vinel, D.; Côme, D. Decrease in sunflower (Helianthus annuus) seed viability caused by high temperature as related to energy metabolism, membrane damage and lipid composition. Physiol. Plant. 2002, 116, 489–496. [Google Scholar] [CrossRef]
- Kibinza, S.; Vinel, D.; Côme, D.; Bailly, C.; Corbineau, F. Sunflower seed deterioration as related to moisture content during ageing, energy metabolism and active oxygen species scavenging. Physiol. Plant. 2006, 128, 496–506. [Google Scholar] [CrossRef]
- Stein, J.C.; Hansen, G. Mannose induces an endonuclease responsible for DNA laddering in plant cells. Plant Physiol. 1999, 121, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Cacas, J.L. Devil inside: Does plant programmed cell death involve the endomembrane system? Plant Cell Environ. 2010, 33, 1453–1473. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.C.D.; Locato, V.; Gara, L.D. Redox regulation in plant programmed cell death. Plant Cell Environ. 2012, 35, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, F.; Cejudo, F.J. Programmed cell death (PCD): An essential process of cereal seed development and germination. Front. Plant Sci. 2014, 5, 366. [Google Scholar] [CrossRef] [PubMed]
- Gavrieli, Y.; Sherman, Y.; Ben-Sasson, S.A. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 1992, 119, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Kubista, M.; Åkerman, B.; Nordén, B. Characterization of interaction between DNA and 4′,6-diamidino-2-phenylindole by optical spectroscopy. Biochemistry 1987, 26, 4545–4553. [Google Scholar] [CrossRef] [PubMed]
- Crowley, L.C.; Marfell, B.J.; Waterhouse, N.J. Detection of DNA fragmentation in apoptotic cells by TUNEL. Cold Spring Harb. Protoc. 2016, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Ansari, B.; Coates, P.J.; Greenstein, B.D.; Hall, P.A. In situ endlabeling detects DNA strand breaks in apoptosis and other physiological and pathological states. J. Pathol. 1993, 170, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kanoh, M.; Takemura, G.; Misao, J.; Hayakawa, Y.; Aoyama, T.; Nishigaki, K.; Noda, T.; Fujiwara, T.; Fukuda, K.; Minatoguchi, S.; et al. Significance of myocytes with positive DNA in situ nick end-labeling (TUNEL) in hearts with dilated cardiomyopathy. Not apoptosis but DNA repair. Circulation 1999, 99, 2757–2764. [Google Scholar] [CrossRef] [PubMed]
- El-Maarouf-Bouteau, H.; Mazuy, C.; Corbineau, F.; Bailly, C. DNA alteration and programmed cell death during ageing of sunflower seed. J. Exp. Bot. 2011, 62, 5003–5011. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Ma, G.; Wang, Q.; Yao, J.; Wang, Y.; Pritchard, H.W.; Wang, X. Spatial and temporal nature of reactive oxygen species production and programmed cell death in elm (Ulmus pumila L.) seeds during controlled deterioration. Plant Cell Environ. 2012, 35, 2045–2059. [Google Scholar] [CrossRef] [PubMed]
- Kranner, I.; Birtić, S.; Anderson, K.M.; Pritchard, H.W. Glutathione half-cell reduction potential: A universal stress marker and modulator of programmed cell death? Free Radic. Biol. Med. 2006, 40, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.; Yang, H.; Fu, Y.B. The associative changes in scutellum nuclear content and morphology with viability loss of naturally aged and accelerated ageing wheat (Triticum aestivum) seeds. Front. Plant Sci. 2016, 7, 1474. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.B.; Yang, M.-Y.; Horbach, C.; Kessler, D.; Diederichsen, A.; You, F.M.; Wang, H. Patterns of SSR variation in bread wheat (Triticum aestivum L.) seeds under ex situ genebank storage and accelerated ageing. Genet. Resour. Crop Evol. 2017, 64, 277–290. [Google Scholar] [CrossRef]
- Hu, Z.-Y.; Hua, W.; Zhang, L.; Deng, L.-B.; Wang, X.-F.; Liu, G.-H.; Hao, W.-J.; Wang, H.-Z. Seed structure characteristics to form ultrahigh oil content in rapeseed. PLoS ONE 2013, 8, e62099. [Google Scholar] [CrossRef] [PubMed]
- Fleming, M.B.; Richards, C.M.; Walters, C. Decline in RNA integrity of dry-stored soybean seeds correlates with loss of germination potential. J. Exp. Bot. 2017, 68, 2219–2230. [Google Scholar] [CrossRef] [PubMed]
- Diederichsen, A.; Jones-Flory, L.L. Accelerated aging tests with seeds of 11 flax (Linum usitatissimum) cultivars. Seed Sci. Technol. 2005, 33, 419–429. [Google Scholar] [CrossRef]
- Association of Official Seed Analysts. Seedling Evaluation Handbook; AOSA: Lincoln, NE, USA, 1992; p. 98. [Google Scholar]
- Rasband, W.S.; ImageJ, U.S. National Institutes of Health, Bethesda, MA, USA. 2018. Available online: http://imagej.nih.gov/ij/ (accessed on 12 April 2018).
- Demeke, T.I.; Ratnayaka, M.; Holigroski, A.P. Assessment of DNA extraction methods for PCR testing of discontinued or unapproved biotech events in single seeds of canola, flax and soybean. Food Control 2012, 24, 44–49. [Google Scholar] [CrossRef]
- Sambrook, J.E.; Fritsch, F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microscopy | Sample Label | DNA Assessment | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Sample | Year | Aging | GR | NS | NSO | PT | DNA(ng/µL) | 260/280 | |
| CN101154 | 2000 | Natural | 90% | 9 | 180 | 54 | N90 | 302.7(74.6) | 1.65(0.01) |
| CN98246 | 1998 | Natural | 32% | 9 | 210 | 192 | N32 | 187.6(34.6) | 1.49(0.07) |
| CN98245 | 1998 | Natural | 22% | 10 | 230 | 124 | N22 | 144.1(28.5) | 1.44(0.04) |
| CN96846 | 1998 | Natural | 91% | 9 | 210 | 194 | N91 | 193.8(48.4) | 1.66(0.02) |
| CN98062 | 1998 | Natural | 33% | 7 | 126 | 68 | N33 | 142.5(6.6) | 1.57(0.05) |
| CN33389 | 1973 | Natural | 3% | 9 | 200 | 124 | N3 | 205.5(26.9) | 1.54(0.03) |
| CN116253 | 2014 | Natural | 95% | 10 | 240 | 106 | N95 | 287.4(62.7) | 1.61(0.02) |
| CN116253 | 2014 | Aged 96 h@43 °C | 54% | 9 | 190 | 55 | A54 | 218.5(22.3) | 1.53(0.05) |
| CN116253 | 2014 | Aged 120 h@44 °C | 47% | 10 | 240 | 142 | A47 | 221.0(20.3) | 1.57(0.04) |
| CN116255 | 2014 | Natural | 95% | 10 | 240 | 119 | N95a | 214.6(93.6) | 1.71(0.26) |
| CN116255 | 2014 | Aged 96 h@43 °C | 44% | 10 | 240 | 106 | A44 | 198.5(22.5) | 1.43(0.01) |
| CN116255 | 2014 | Aged 120 h@44 °C | 12% | 10 | 240 | 100 | A12 | 204.9(35.5) | 1.55(0.03) |
| Total | 112 | 2546 | 1384 | ||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.-B.; Ahmed, Z.; Yang, H.; Horbach, C. TUNEL Assay and DAPI Staining Revealed Few Alterations of Cellular Morphology in Naturally and Artificially Aged Seeds of Cultivated Flax. Plants 2018, 7, 34. https://doi.org/10.3390/plants7020034
Fu Y-B, Ahmed Z, Yang H, Horbach C. TUNEL Assay and DAPI Staining Revealed Few Alterations of Cellular Morphology in Naturally and Artificially Aged Seeds of Cultivated Flax. Plants. 2018; 7(2):34. https://doi.org/10.3390/plants7020034
Chicago/Turabian StyleFu, Yong-Bi, Zaheer Ahmed, Hui Yang, and Carolee Horbach. 2018. "TUNEL Assay and DAPI Staining Revealed Few Alterations of Cellular Morphology in Naturally and Artificially Aged Seeds of Cultivated Flax" Plants 7, no. 2: 34. https://doi.org/10.3390/plants7020034
APA StyleFu, Y. -B., Ahmed, Z., Yang, H., & Horbach, C. (2018). TUNEL Assay and DAPI Staining Revealed Few Alterations of Cellular Morphology in Naturally and Artificially Aged Seeds of Cultivated Flax. Plants, 7(2), 34. https://doi.org/10.3390/plants7020034