Gas Chromatography-Mass Spectrometry and Single Nucleotide Polymorphism-Genotype-By-Sequencing Analyses Reveal the Bean Chemical Profiles and Relatedness of Coffea canephora Genotypes in Nigeria

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
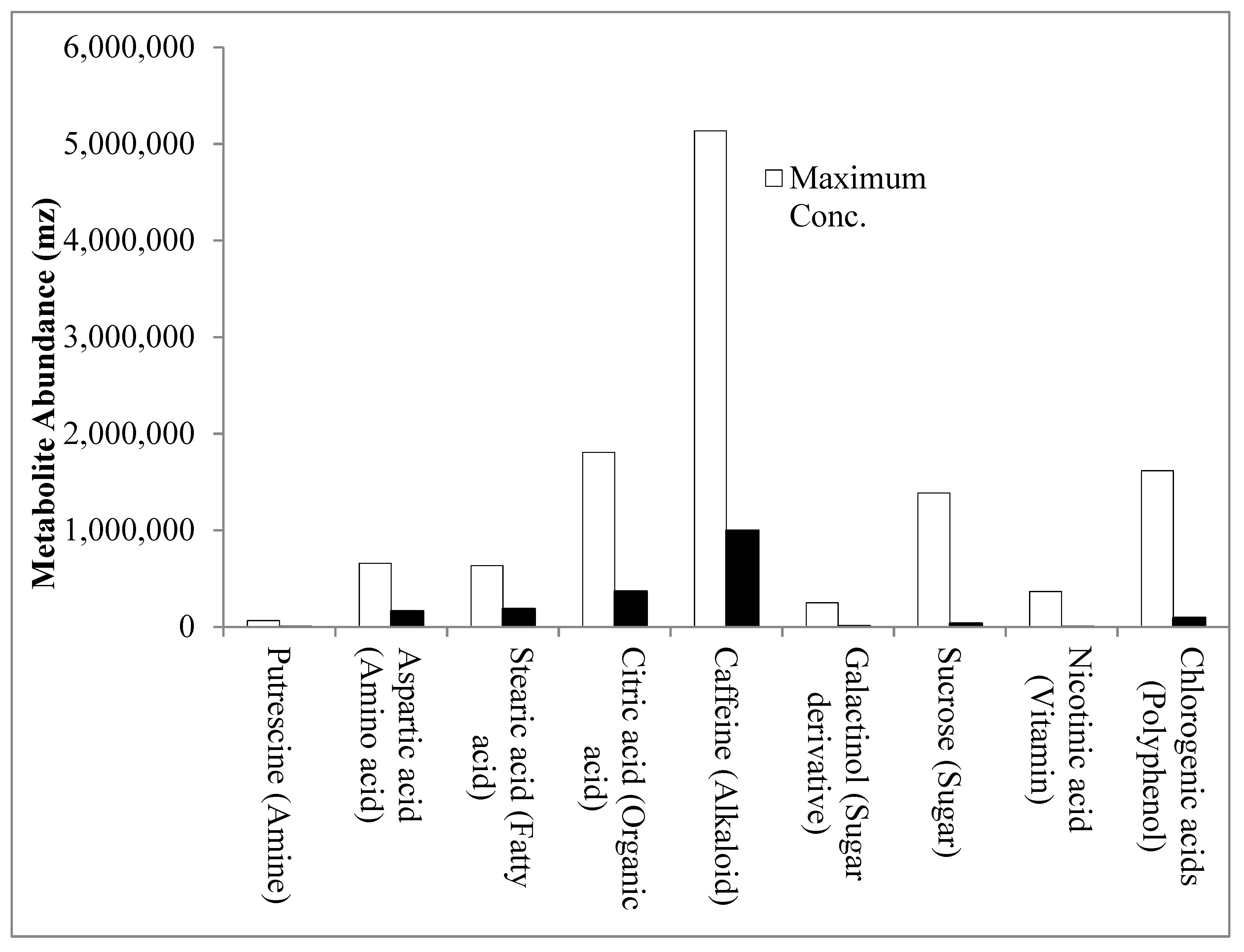
2.1. Metabolite Profiles of the Three C. canephora Genotypes: “Niaouli’, ‘Kouilou’ and ‘Java Robusta’
2.2. Metabolomic Markers for Differentiating Genotypes
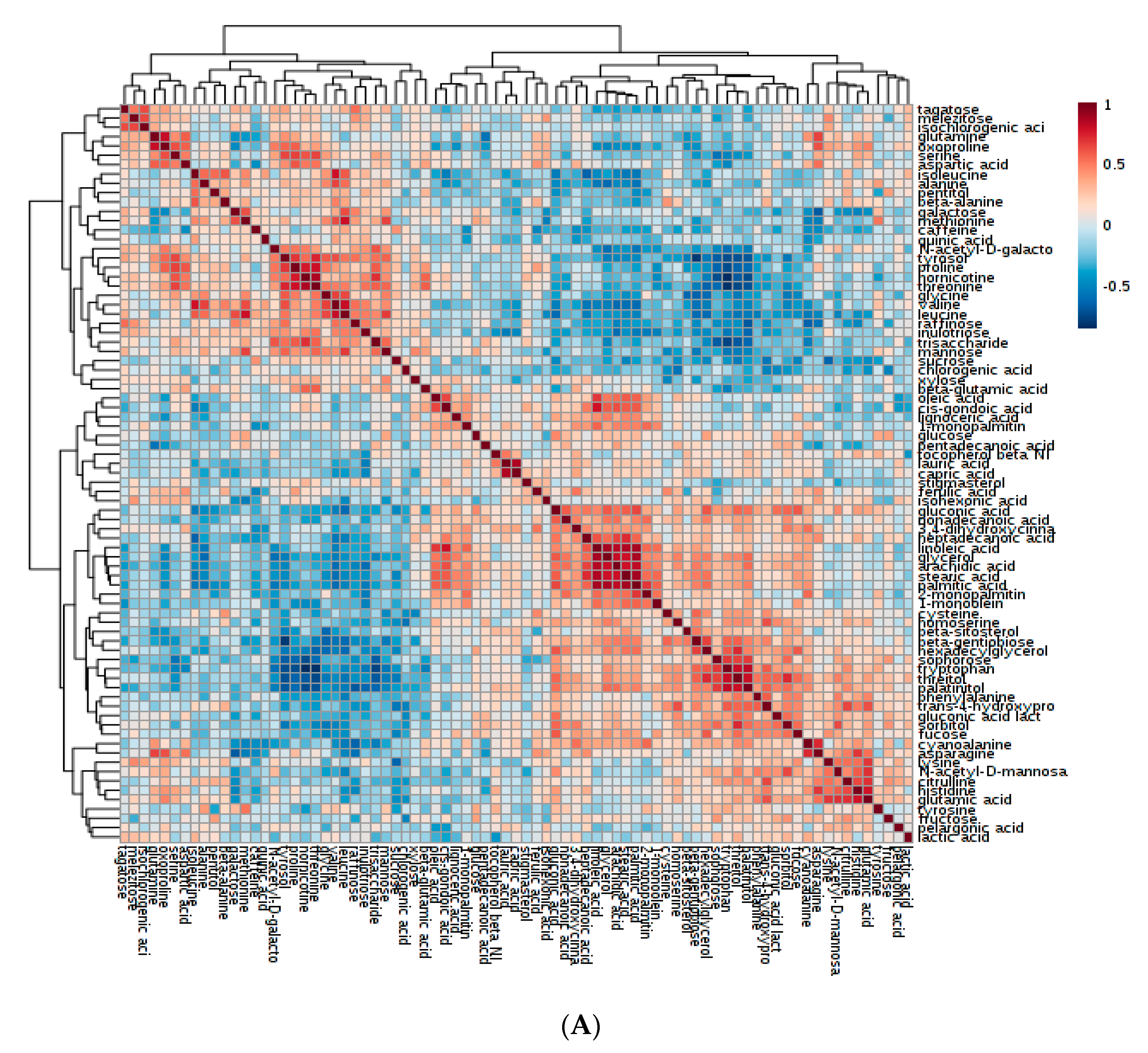
2.3. Metabolite-to-Metabolite Correlations and Their Potential Influence on Cup Quality and Other Beneficial Traits
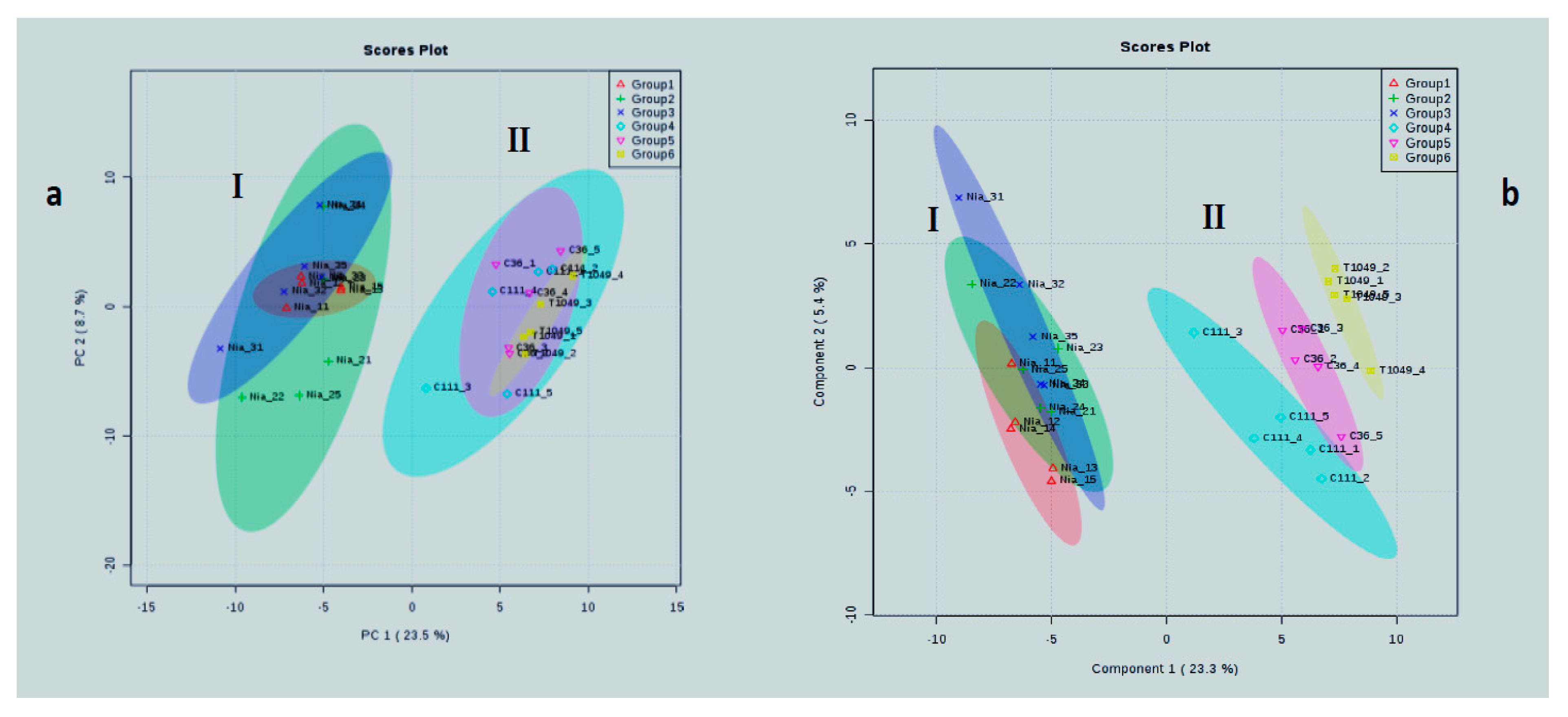
2.4. Metabolite and Genomic Diversity within and among Varieties
2.5. Genotypes with Favorable Bean Quality Traits
3. Materials and Methods
3.1. Single Nucleotide Polymorphism Genotype-By-Sequencing Analysis
3.2. GC-MS Analysis
3.3. Statistical Analysis of Metabolomics Data
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Dicum, G.; Luttinger, N. The Coffee Book: Anatomy of an Industry from the Crop to the Last Drop; The New Press: New York, NY, USA, 2006. [Google Scholar]
- Oxfam. The Coffee Market: A Background Study; Oxfam: London, UK, 2001. [Google Scholar]
- Davis, A.P.; Chadburn, H.; Moat, J.; O’Sullivan, R.; Hargreaves, S.; Lughadha, E.N. High extinction risk for wild coffee species and implications for coffee sector sustainability. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Ayegboyin, K.O.; Famaye, A.O.; Akinrinde, E.A.; Adejobi, K.B.; Akanbi, O.S.O. Growth performance and nutrient uptake of Coffea canephora Prierre Ex. Froehner grown in contrasting soils. Int. J. Res. 2015, 1, 25–31. [Google Scholar]
- Leroy, T.; Ribeyre, F.; Bertrand, B.; Charmetant, P.; Dufour, M.; Montagnon, C.; Marraccini, P.; Pot, D. Genetics of coffee quality. Br. J. Plant. Phys. 2006, 18, 229–242. [Google Scholar] [CrossRef]
- Omolaja, S.S. Biochemical Characterization, Improvement and Propagation of Coffea canephora. Cocoa Research Institute of Nigeria Annual Report; 2000; pp. 32–35. Available online: http://www.crin.gov.ng (accessed on 2 October 2019).
- Anagbogu, C.F.; Bhattacharjee, R.; Ilori, C.; Tongyoo, P.; Dada, K.E.; Muyiwa, A.A.; Beckles, D.M. Genetic diversity and re-classification of coffee (Coffea canephora Pierre ex A. Froehner) from South Western Nigeria through genotyping-by-sequencing-single nucleotide polymorphism analysis. Genet. Resour. Crop. Evol. 2019, 66, 685–695. [Google Scholar] [CrossRef]
- Choi, M.Y.; Choi, W.; Park, J.H.; Lim, J.; Kwon, S.W. Determination of coffee origins by integrated metabolomic approach of combining multiple analytical data. Food Chem. 2010, 121, 1260–1268. [Google Scholar] [CrossRef]
- Kwon, D.J.; Jeong, H.J.; Moon, H.; Kim, H.N.; Cho, J.H.; Lee, J.E.; Hong, K.S.; Hong, Y.S. Assessment of green coffee bean metabolites dependent on coffee quality using a H-1 NMR-based metabolomics approach. Food Res. Int. 2015, 67, 175–182. [Google Scholar] [CrossRef]
- Beckles, D.M.; Roessner, U. Plant metabolomics: Applications and opportunities for agricultural biotechnology. Plant Biotech. Agric. 2012, 1st Edition. 67–81. [Google Scholar]
- Flament, I. Coffee Flavor Chemistry; John Wiley & Sons Ltd.: New York, NY, USA, 2002; p. 424. [Google Scholar]
- Ribeiro, J.S.; Augusto, F.; Salva, T.J.G.; Thomaziello, R.A.; Ferreira, M.M.C. Prediction of sensory properties of Brazilian Arabica roasted coffees by headspace solid phase microextraction-gas chromatography and partial least squares. Anal. Chim. Acta 2009, 634, 172–179. [Google Scholar] [CrossRef]
- Dessalegn, Y.; Labuschagne, M.T.; Osthoff, G.; Herselman, L. Genetic diversity and correlation of bean caffeine content with cup quality and green bean physical characteristics in coffee (Coffea arabical L.). J. Sci. Food Agr. 2008, 88, 1726–1730. [Google Scholar] [CrossRef]
- Belay, A.; Gholap, A.V. Characterization and determination of chlorogenic acids (CGA) is coffee beans by UV-Vis spectroscopy. Afr. J. Pure Appl. Chem. 2009, 3, 234–240. [Google Scholar]
- Joet, T.; Bertrand, B.; Dussert, S. Environmental Effects on Coffee Seed Biochemical Composition and Quality Attributes: A Genomic Perspective. In Proceedings of the 25th International Conference on Coffee Science, Armenia, Colombia, 8–13 September 2014; pp. 42–49. [Google Scholar]
- Bertrand, B.; Boulanger, R.; Dussert, S.; Ribeyre, F.; Berthiot, L.; Descroix, F.; Joet, T. Climatic factors directly impact the volatile organic compound fingerprint in green Arabica coffee bean as well as coffee beverage quality. Food Chem. 2012, 135, 2575–2583. [Google Scholar] [CrossRef] [PubMed]
- Oksman-Caldentey, K.M.; Saito, K. Integrating genomics and metabolomics for engineering plant metabolic pathways. Curr. Opin. Biotech. 2005, 16, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Duarte, G.S.; Pereira, A.A.; Farah, A. Chlorogenic acids and other relevant compounds in Brazilian coffees processed by semi-dry and wet post-harvesting methods. Food Chem. 2010, 118, 851–855. [Google Scholar] [CrossRef]
- Farah, A.; Monteiro, M.C.; Calado, V.; Franca, A.S.; Trugo, L.C. Correlation between cup quality and chemical attributes of Brazilian coffee. Food Chem. 2006, 98, 373–380. [Google Scholar] [CrossRef]
- Arana, V.A.; Medina, J.; Alarcon, R.; Moreno, E.; Heintz, L.; Schafer, H.; Wist, J. Coffee’s country of origin determined by NMR: The Colombian case. Food Chem. 2015, 175, 500–506. [Google Scholar] [CrossRef]
- Jumhawan, U.; Putri, S.P.; Yusianto; Bamba, T.; Fukusaki, E. Application of gas chromatography/flame ionization detector-based metabolite fingerprinting for authentication of Asian palm civet coffee (Kopi Luwak). J. Biosci. Bioeng. 2015, 120, 555–561. [Google Scholar] [CrossRef]
- Dong, W.J.; Tan, L.H.; Zhao, J.P.; Hu, R.S.; Lu, M.Q. Characterization of Fatty Acid, Amino Acid and Volatile Compound Compositions and Bioactive Components of Seven Coffee (Coffea robusta) Cultivars Grown in Hainan Province, China. Molecules 2015, 20, 16687–16708. [Google Scholar] [CrossRef]
- CIRAD. Genes Involved in Coffee Quality; Montpellier University: Montpellier, France, 2006. [Google Scholar]
- Ky, C.L.; Louarn, J.; Dussert, S.; Guyot, B.; Hamon, S.; Noirot, M. Caffeine, trigonelline, chlorogenic acids and sucrose diversity in wild Coffea arabica L. and C-canephora P. accessions. Food Chem. 2001, 75, 223–230. [Google Scholar] [CrossRef]
- Steinfath, M.; Strehmel, N.; Peters, R.; Schauer, N.; Groth, D.; Hummel, J.; Steup, M.; Selbig, J.; Kopka, J.; Geigenberger, P.; et al. Discovering plant metabolic biomarkers for phenotype prediction using an untargeted approach. Plant. Biotech. J. 2010, 8, 900–911. [Google Scholar] [CrossRef]
- Liu, C.; Yang, N.; Yang, Q.; Ayed, C.; Linforth, R.; Fisk, I.D. Enhancing Robusta coffee aroma by modifying flavour precursors in the green coffee bean. Food Chem. 2019, 281, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Aziz, S.A.; Mohamed, S. Sensory aroma from Maillard reaction of individual and combinations of amino acids with glucose in acidic conditions. Int. J. Food Sci. Tech. 2008, 43, 1512–1519. [Google Scholar] [CrossRef]
- Setoyama, D.; Iwasa, K.; Seta, H.; Shimizu, H.; Fujimura, Y.; Miura, D.; Wariishi, H.; Nagai, C.; Nakahara, K. High-Throughput Metabolic Profiling of Diverse Green Coffea arabica Beans Identified Tryptophan as a Universal Discrimination Factor for Immature Beans. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, C.; Straface, E.; Modesti, D.; Coni, E.; Cantafora, A.; De Vincenzi, M.; Malorni, W.; Masella, R. Tyrosol, the major olive oil biophenol, protects against oxidized-LDL-induced injury in Caco-2 cells. J. Nutr. 1999, 129, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, L.P.; Borém, F.M.; Ribeiro, F.C.; Giomo, G.S.; Taveira, J.H.S.; Malta, M.R. Fatty acid profiles and parameters of quality of specialty coffees produced in different Brazilian regions. Afr. J. Agric. Res. 2015, 10, 3484–3493. [Google Scholar]
- Calvino, A.M.; Garciamedina, M.R.; Comettomuniz, J.E. Interactions in Caffeine-Sucrose and Coffee-Sucrose Mixtures - Evidence of Taste and Flavor Suppression. Chem. Senses 1990, 15, 505–519. [Google Scholar] [CrossRef]
- Nehlig, A.; Daval, J.L.; Debry, G. Caffeine and the Central-Nervous-System - Mechanisms of Action, Biochemical, Metabolic and Psychostimulant Effects. Brain Res. Rev. 1992, 17, 139–169. [Google Scholar] [CrossRef]
- Kumar, V.; Kaur, J.; Panghal, A.; Kaur, S.; Handa, V. Caffeine: A boon or bane. Nutr. Food Sci. 2018, 48, 61–75. [Google Scholar] [CrossRef]
- Caporaso, N.; Whitworth, M.B.; Grebby, S.; Fisk, I.D. Non-destructive analysis of sucrose, caffeine and trigonelline on single green coffee beans by hyperspectral imaging. Food Res. Int. 2018, 106, 193–203. [Google Scholar] [CrossRef]
- Hellmers, F.; Takors, R.; Thum, O. Robust enzyme immobilizates for industrial isomalt production. Mol. Catal. 2018, 445, 293–298. [Google Scholar] [CrossRef]
- Toci, A.T.; Farah, A. Volatile compounds as potential defective coffee beans’ markers. Food Chem. 2008, 108, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Coffee: Recent Developments; Clarke, R.J.; Vitzthum, O.G. (Eds.) Wiley-Blackwell Science Ltd.: Oxford, UK, 2001; pp. 50–67. [Google Scholar]
- Grosch, W.; Czerny, M.; Mayer, F.; Moors, A. Sensory Studies on the Key Odorants of Roasted Coffee. In Caffeinated Beverages; American Chemical Society: Washington, DC, USA, 2000; Volume 754, pp. 202–209. [Google Scholar]
- Arnold, U.; Ludiwg, E.; Kuhn, R.; Moschwitzer, U. Analysis of Free Amino-Acids in Green Coffee Beans. 1. Determination of Amino-Acids after Precolumn Derivatization Using 9-Fluorenylmethylchloroformate. Z Lebensm Unters For. 1994, 199, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant. Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.C.; Liong, C.Y.; Jemain, A.A. Partial least squares-discriminant analysis (PLS-DA) for classification of high-dimensional (HD) data: A review of contemporary practice strategies and knowledge gaps. Analyst 2018, 143, 3526–3539. [Google Scholar] [CrossRef] [PubMed]
- Stamova, B.S.; Roessner, U.; Suren, S.; Laudencia-Chingcuanco, D.; Bacic, A.; Beckles, D.M. Metabolic profiling of transgenic wheat over-expressing the high-molecular-weight Dx5 glutenin subunit. Metabolomics 2009, 5, 239–252. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics - the link between genotypes and phenotypes. Plant. Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Heuberger, A.L.; Lewis, M.R.; Chen, M.H.; Brick, M.A.; Leach, J.E.; Ryan, E.P. Metabolomic and Functional Genomic Analyses Reveal Varietal Differences in Bioactive Compounds of Cooked Rice. PLoS ONE 2010, 5, e12915. [Google Scholar] [CrossRef]
- Privat, I.; Foucrier, S.; Prins, A.; Epalle, T.; Eychenne, M.; Kandalaft, L.; Caillet, V.; Lin, C.W.; Tanksley, S.; Foyer, C.; et al. Differential regulation of grain sucrose accumulation and metabolism in Coffea arabica (Arabica) and Coffea canephora (Robusta) revealed through gene expression and enzyme activity analysis. New Phytol. 2008, 178, 781–797. [Google Scholar] [CrossRef]
- Perrois, C.; Strickler, S.R.; Mathieu, G.; Lepelley, M.; Bedon, L.; Michaux, S.; Husson, J.; Mueller, L.; Privat, I. Differential regulation of caffeine metabolism in Coffea arabica (Arabica) and Coffea canephora (Robusta). Planta 2015, 241, 179–191. [Google Scholar] [CrossRef]
- Geromel, C.; Ferreira, L.P.; Guerreiro, S.M.C.; Cavalari, A.A.; Pot, D.; Pereira, L.F.P.; Leroy, T.; Vieira, L.G.E.; Mazzafera, P.; Marraccini, P. Biochemical and genomic analysis of sucrose metabolism during coffee (Coffea arabica) fruit development. J. Exp. Bot. 2006, 57, 3243–3258. [Google Scholar] [CrossRef]
- Zheng, X.; Levine, D.; Shen, J.; Gogarten, S.M.; Laurie, C.; Weir, B.S. A high-performance computing toolset for relatedness and principal component analysis on SNP data. Bioinformatics 2012, 28, 3326–3328. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O.; Wohlgemuth, G.; Scholz, M.; Kind, T.; Lee, D.Y.; Lu, Y.; Moon, S.; Nikolau, B. Quality control for plant metabolomics: Reporting MSI-compliant studies. Plant. J. 2008, 53, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Yamamoto, M.; Xia, J.G. MetaboAnalystR 2.0: From Raw Spectra to Biological Insights. Metabolites 2019, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2014. Available online: https://www.R-project.org/ (accessed on 6 October 2019).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolomic Markers | f. Value | p. Value | −Log10(p) | FDR |
|---|---|---|---|---|
| 6404 | 37.478 | 1.41 × 10−10 | 9.8511 | 4.30 × 10−8 |
| 125788 | 23.897 | 1.35 × 10−8 | 7.8688 | 2.06 × 10−6 |
| Citramalic acid | 21.799 | 3.29 × 10−8 | 7.4826 | 2.96 × 10−6 |
| Tryptophan | 21.428 | 3.88 × 10−8 | 7.4112 | 2.96 × 10−6 |
| 34007 | 14.774 | 1.17 × 10−6 | 5.9327 | 4.29 × 10−5 |
| Palatinitol | 14.587 | 1.30 × 10−6 | 5.8846 | 4.29 × 10−5 |
| 134465 | 14.39 | 1.47 × 10−6 | 5.8333 | 4.29 × 10−5 |
| 2193 | 14.304 | 1.55 × 10−6 | 5.8109 | 4.29 × 10−5 |
| Erythritol | 13.379 | 2.74 × 10−6 | 5.5625 | 6.96 × 10−5 |
| 134464 | 13.05 | 3.38 × 10−6 | 5.4711 | 7.36 × 10−5 |
| Threitol | 12.92 | 3.68 × 10−6 | 5.4344 | 7.48 × 10−5 |
| 1-methylgalactose NIST | 12.663 | 4.35 × 10−6 | 5.3615 | 8.29 × 10−5 |
| 127358 | 12.044 | 6.58 × 10−6 | 5.1815 | 0.0001 |
| Gluconic acid | 11.973 | 6.91 × 10−6 | 5.1606 | 0.0001 |
| 2-hydroxyglutaric acid | 11.239 | 1.15 × 10−5 | 4.9383 | 0.000146 |
| 3182 | 11.072 | 1.30 × 10−5 | 4.8861 | 0.000152 |
| Maleic acid | 10.247 | 2.39 × 10−5 | 4.6219 | 0.000241 |
| Sophorose | 10.215 | 2.45 × 10−5 | 4.6114 | 0.000241 |
| 16594 | 9.5222 | 4.18 × 10−5 | 4.3787 | 0.000399 |
| 4850 | 9.3508 | 4.79 × 10−5 | 4.3195 | 0.000443 |
| 125830 | 8.252 | 0.000119 | 3.9236 | 0.00107 |
| Butane-2,3-diol NIST | 8.0335 | 0.000144 | 3.8414 | 0.001253 |
| 1,2-anhydro-myo-inositol NIST | 8.0039 | 0.000148 | 3.8301 | 0.001253 |
| Pseudo uridine | 7.6369 | 0.000205 | 3.6888 | 0.001688 |
| 6-deoxyglucitol | 7.2396 | 0.000294 | 3.5316 | 0.00236 |
| Mannitol | 7.0457 | 0.000352 | 3.4532 | 0.002755 |
| 102728 | 6.4686 | 0.000612 | 3.2131 | 0.004446 |
| 133018 | 5.6757 | 0.001363 | 2.8655 | 0.009037 |
| Hexadecylglycerol NIST | 4.8794 | 0.003208 | 2.4938 | 0.019967 |
| Arachidic acid | 4.5156 | 0.004832 | 2.3159 | 0.026794 |
| Sorbitol | 4.4837 | 0.005011 | 2.3001 | 0.02724 |
| trans-4-hydroxyproline | 4.0293 | 0.008518 | 2.0697 | 0.042538 |
| beta-gentiobiose | 3.8623 | 0.010403 | 1.9829 | 0.048073 |
| Fisher’s Least Square Difference (LSD) identified ‘Niaouli’ to be higher in content in the metabolites listed here compared to ‘Java/Kouilou’. | ||||
| Threonine | 16.164 | 5.27 × 10−7 | 6.2779 | 3.22 × 10−5 |
| Uric acid | 15.162 | 9.30 × 10−7 | 6.0315 | 4.29 × 10−5 |
| Nornicotine | 13.222 | 3.03 × 10−6 | 5.5189 | 7.10 × 10−5 |
| Adipic | 12.564 | 4.64 × 10−6 | 5.3331 | 8.33 × 10−5 |
| 17094 | 12.205 | 5.90 × 10−6 | 5.229 | 9.47 × 10−5 |
| Pentitol | 11.778 | 7.90 × 10−6 | 5.1025 | 0.000109 |
| 5-hydroxynorvaline NIST | 11.414 | 1.02 × 10−5 | 4.992 | 0.000135 |
| Tyrosol | 11.076 | 1.30 × 10−5 | 4.8875 | 0.000152 |
| Proline | 10.719 | 1.68 × 10−5 | 4.7747 | 0.00019 |
| Methanolphosphate | 10.547 | 1.91 × 10−5 | 4.7195 | 0.000201 |
| Trisaccharide | 5.7878 | 0.001214 | 2.9159 | 0.008225 |
| 110009 | 4.2989 | 0.006203 | 2.2074 | 0.032621 |
| Isocitric acid | 4.114 | 0.007704 | 2.1133 | 0.039825 |
| Fisher’s LSD identified ‘Java’ to be higher in content in the metabolites listed here compared to ‘Kouilou’ | ||||
| Lyxitol | 12.382 | 5.24 × 10−6 | 5.2807 | 8.88 × 10−5 |
| Glycerol | 5.4388 | 0.001748 | 2.7574 | 0.01111 |
| Metabolite Identified as Markers | Correlation Coefficient (r2) | |
|---|---|---|
| Sucrose | Caffeine | |
| Caffeine | 0.4079 | - |
| 16548 | 0.4044 | 0.5182 |
| 6404 | −0.3705 | - |
| Palatinitol | −0.4961 | - |
| 68 | 0.4991 | 0.4502 |
| Tryptophan | −0.4599 | - |
| Metabolites | Metabolite Levels | ||||
|---|---|---|---|---|---|
| Very High | High | Medium | Low | Very Low | |
| Caffeine | Nia_24, Nia_25 | Nia_15, Nia_14 | |||
| CGA | Nia_25 | Nia_14 | C111_2 | ||
| Sucrose | Nia_25, Nia_22, Nia_15, Nia_14 | Nia_22 | Nia_24 | C36_5 | |
| Quinic acid | Nia_25, Nia_31, Nia_34 | C36_1 | Nia_14 | ||
| Butane-2,3 diol | C36_2 | All Nia except Nia_25 | Nia_22 | ||
| Saccharic acid | Nia_15 | Nia_22, Nia_11, Nia_31 | |||
| Ferulic acid | Nia_33 | ||||
| Tryptophan | C36_5 | Nia_24 | |||
| Putrescine | C36_5 | Nia_22, Nia_25, Nia_32, Nia_33 | C111_5 | ||
| Proline | Nia_33 | C111_3 | |||
| Relative Metabolite Levels | |||
|---|---|---|---|
| Coffee Type and Perceived Quality | High | Low | Source |
| Palm civet (Superior) | Citric acid, malic acid, and glycolic acid | Quinic acid, caffeine, and caffeic acid | [21] |
| Coffea arabica (Good) | Sucrose, triglyceride, and threonine, proline | Caffeine, chlorogenic acid, aminobutyric acid (GABA), quinic acid, choline, acetic acid, and fatty acid | [9,19,27] |
| C. canephora (Poor) | Caffeine and chlorogenic acid | Sucrose | [45,46] |
| Variety | ‘Niaouli’ | ‘Kouilou’ | ‘Java Robusta’ | |||
|---|---|---|---|---|---|---|
| Group/Genotype | Group 1 (Nia_1) | Group 2 (Nia_2) | Group 3 (Nia_3) | Group 4 (C111) | Group 5 (C36) | Group 6 (T1049) |
| Sample Symbols | Nia_11 | Nia_21 | Nia_31 | C111_1 | C36_1 | T1049_1 |
| Nia_12 | Nia_22 | Nia_32 | C111_2 | C36_2 | T1049_2 | |
| Nia_13 | Nia_23 | Nia_33 | C111_3 | C36_3 | T1049_3 | |
| Nia_14 | Nia_24 | Nia_34 | C111_4 | C36_4 | T1049_4 | |
| Nia_15 | Nia_25 | Nia_35 | C111_5 | C36_5 | T1049_5 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anagbogu, C.F.; Ilori, C.O.; Bhattacharjee, R.; Olaniyi, O.O.; Beckles, D.M. Gas Chromatography-Mass Spectrometry and Single Nucleotide Polymorphism-Genotype-By-Sequencing Analyses Reveal the Bean Chemical Profiles and Relatedness of Coffea canephora Genotypes in Nigeria. Plants 2019, 8, 425. https://doi.org/10.3390/plants8100425
Anagbogu CF, Ilori CO, Bhattacharjee R, Olaniyi OO, Beckles DM. Gas Chromatography-Mass Spectrometry and Single Nucleotide Polymorphism-Genotype-By-Sequencing Analyses Reveal the Bean Chemical Profiles and Relatedness of Coffea canephora Genotypes in Nigeria. Plants. 2019; 8(10):425. https://doi.org/10.3390/plants8100425
Chicago/Turabian StyleAnagbogu, Chinyere F., Christopher O. Ilori, Ranjana Bhattacharjee, Olufemi O. Olaniyi, and Diane M. Beckles. 2019. "Gas Chromatography-Mass Spectrometry and Single Nucleotide Polymorphism-Genotype-By-Sequencing Analyses Reveal the Bean Chemical Profiles and Relatedness of Coffea canephora Genotypes in Nigeria" Plants 8, no. 10: 425. https://doi.org/10.3390/plants8100425
APA StyleAnagbogu, C. F., Ilori, C. O., Bhattacharjee, R., Olaniyi, O. O., & Beckles, D. M. (2019). Gas Chromatography-Mass Spectrometry and Single Nucleotide Polymorphism-Genotype-By-Sequencing Analyses Reveal the Bean Chemical Profiles and Relatedness of Coffea canephora Genotypes in Nigeria. Plants, 8(10), 425. https://doi.org/10.3390/plants8100425