Desert Soil Microbes as a Mineral Nutrient Acquisition Tool for Chickpea (Cicer arietinum L.) Productivity at Different Moisture Regimes

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Morphological Characterization of Isolated Strains
2.2. Biochemical Characterization of Isolated Strains
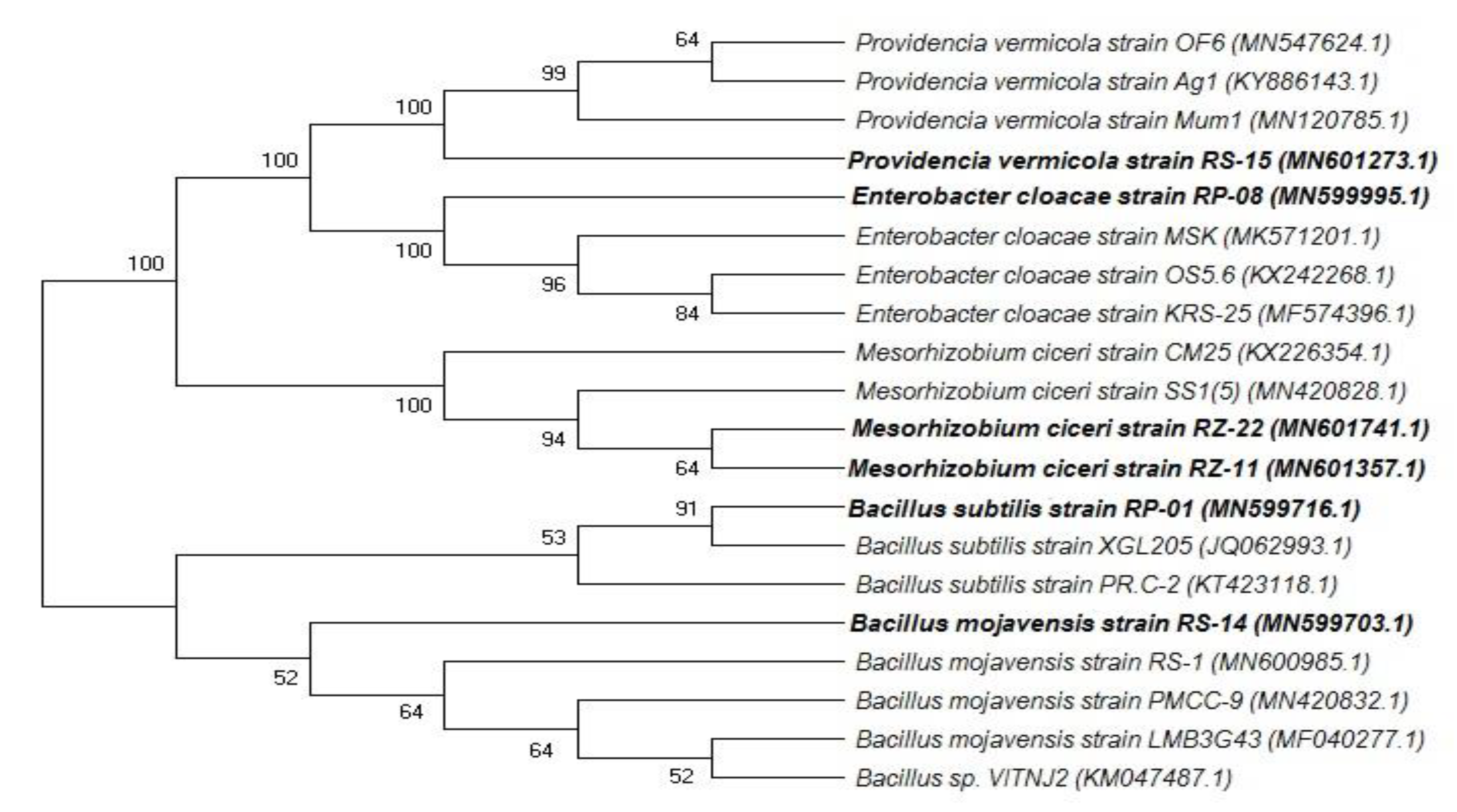
2.3. Molecular Identification of Isolates
2.4. Quantification Assay of ACC-Deaminase, EPS, IAA and P Solubilization
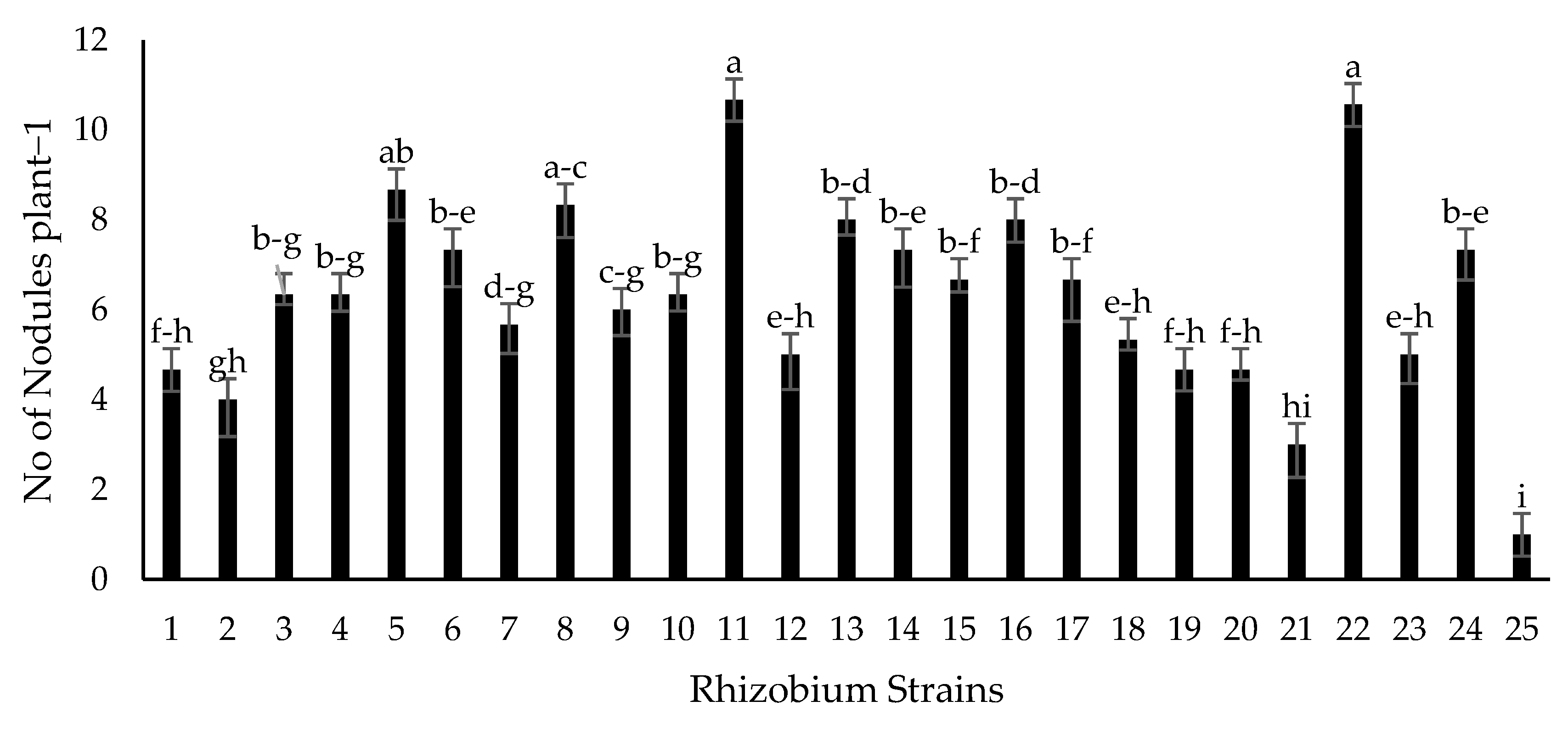
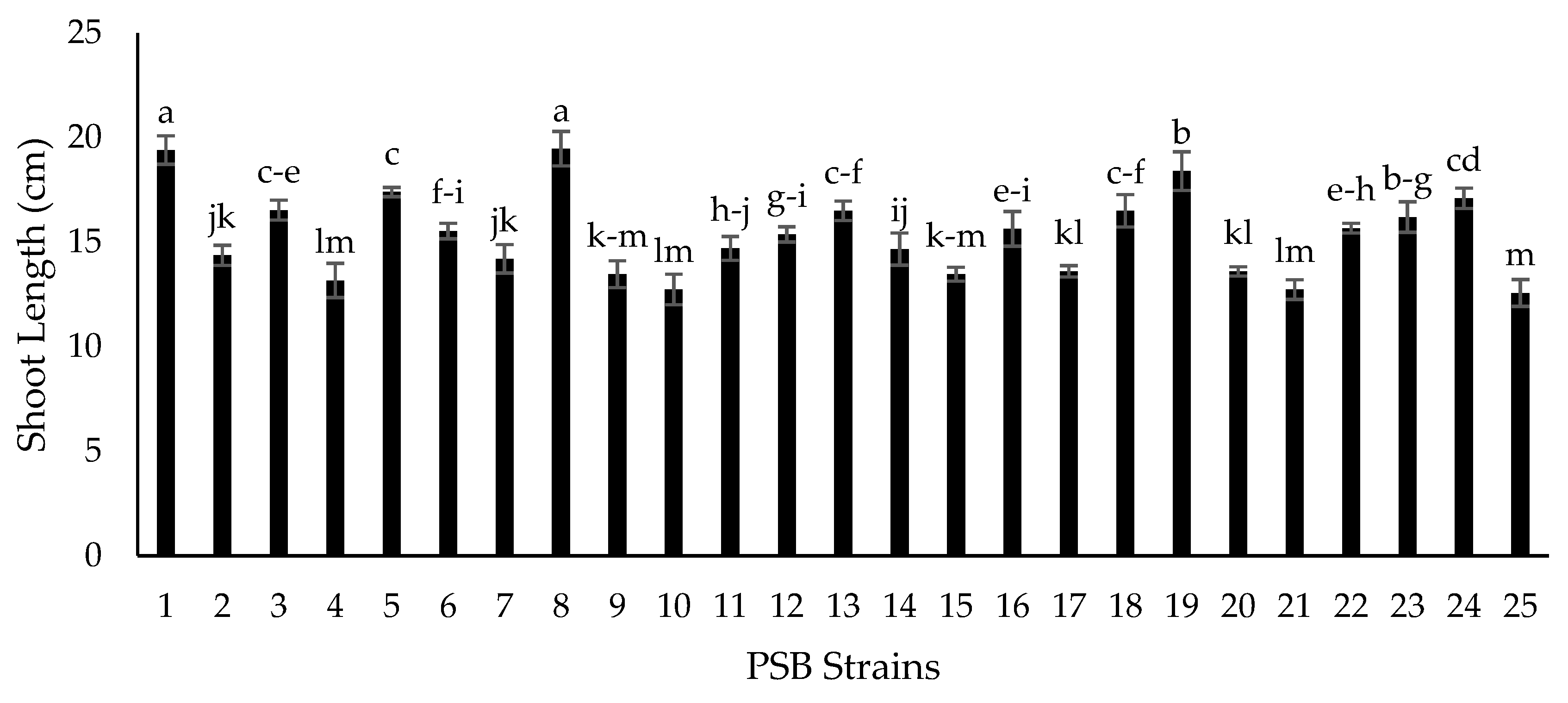
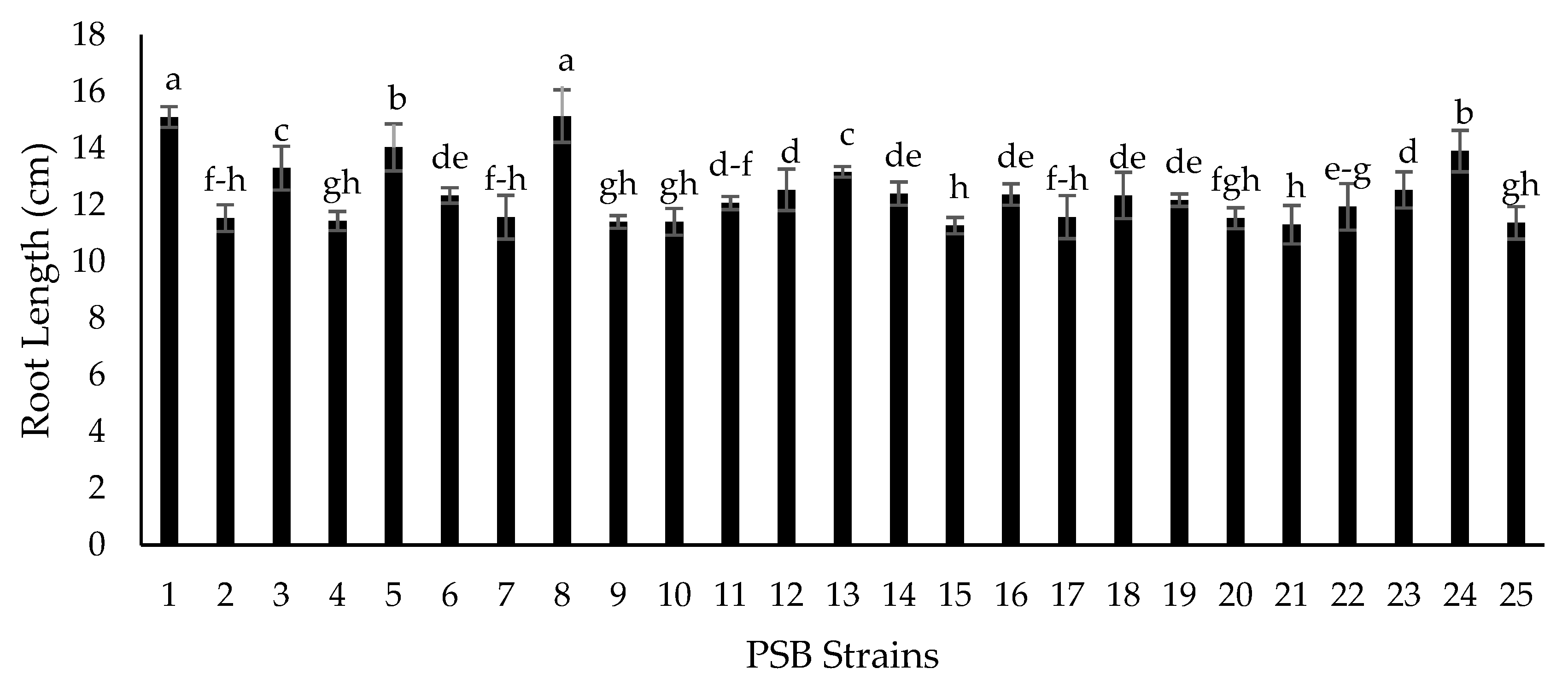
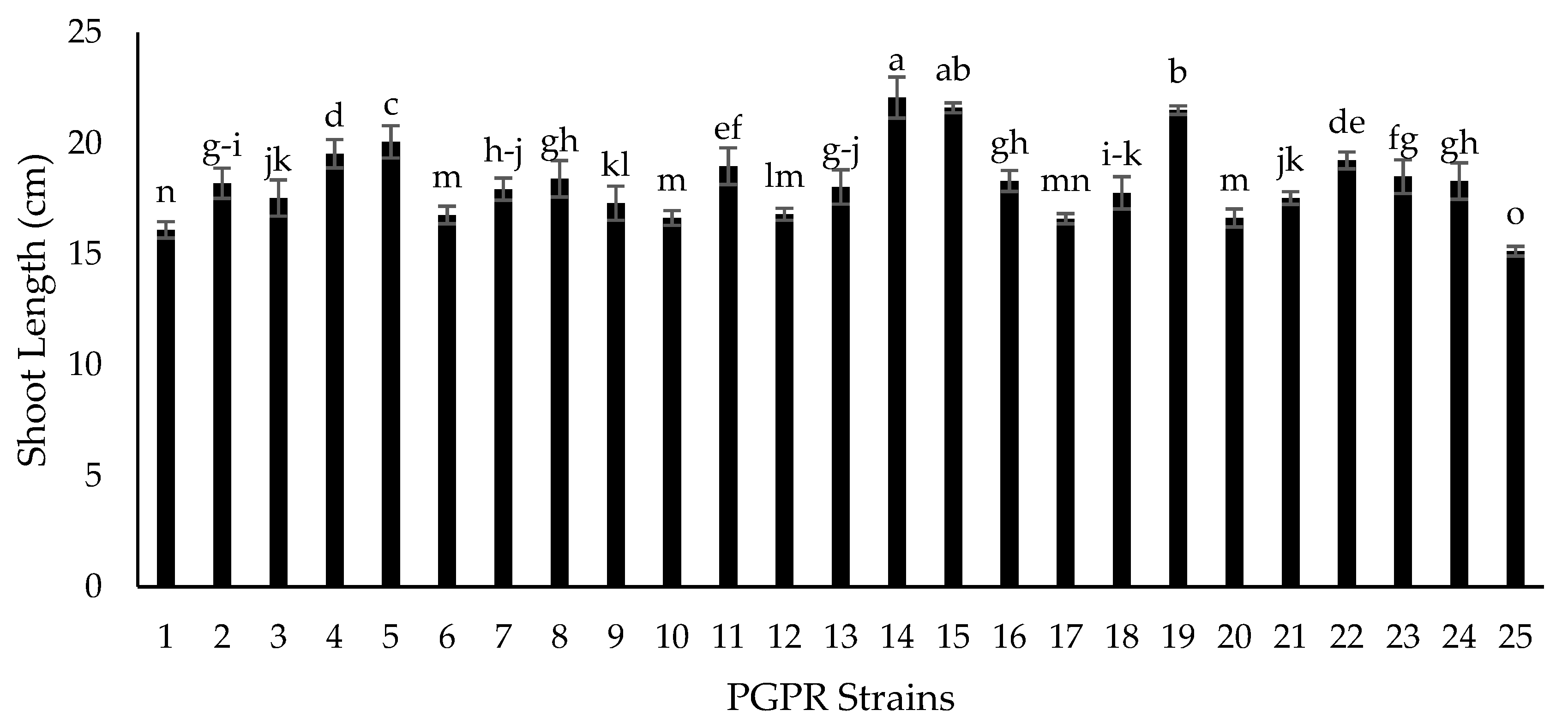
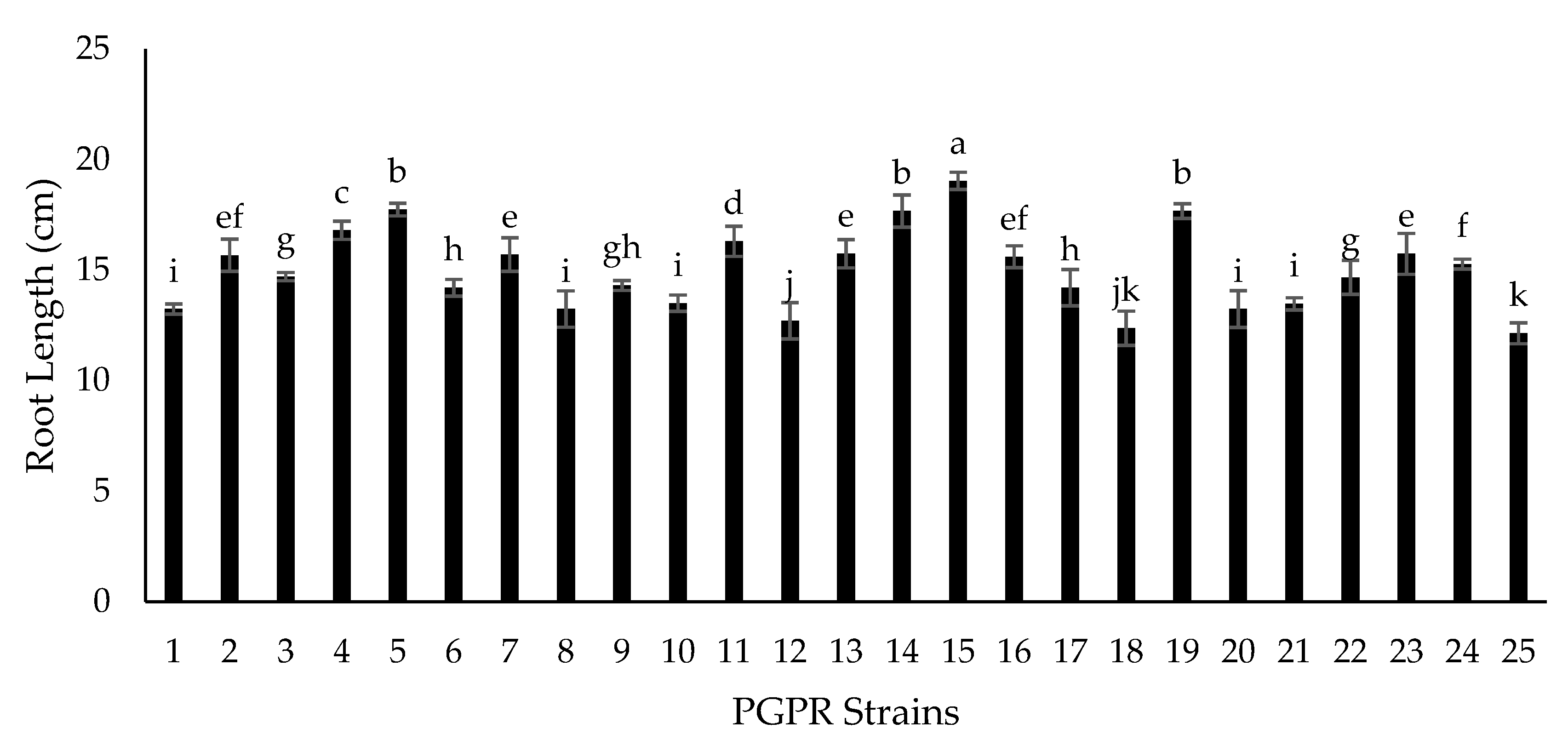
2.5. Growth Chamber Experiment for Screening of Isolates
2.6. Glass House Experiment at Different Moisture Regimes
2.6.1. Physiological Attributes and Nutrient Acquisition of Chickpea
2.6.2. Isolates Survival in Rhizospheric Soil
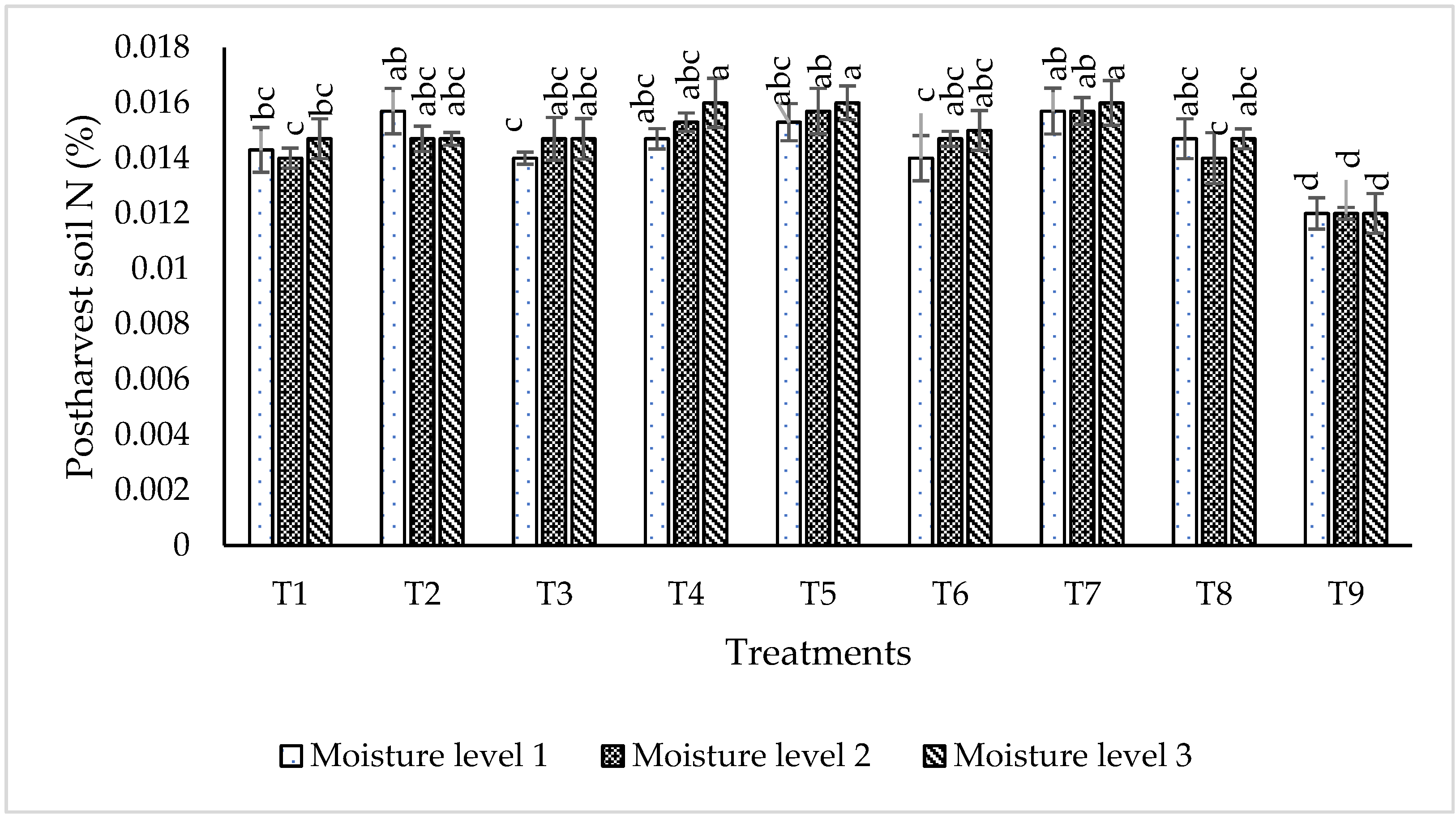
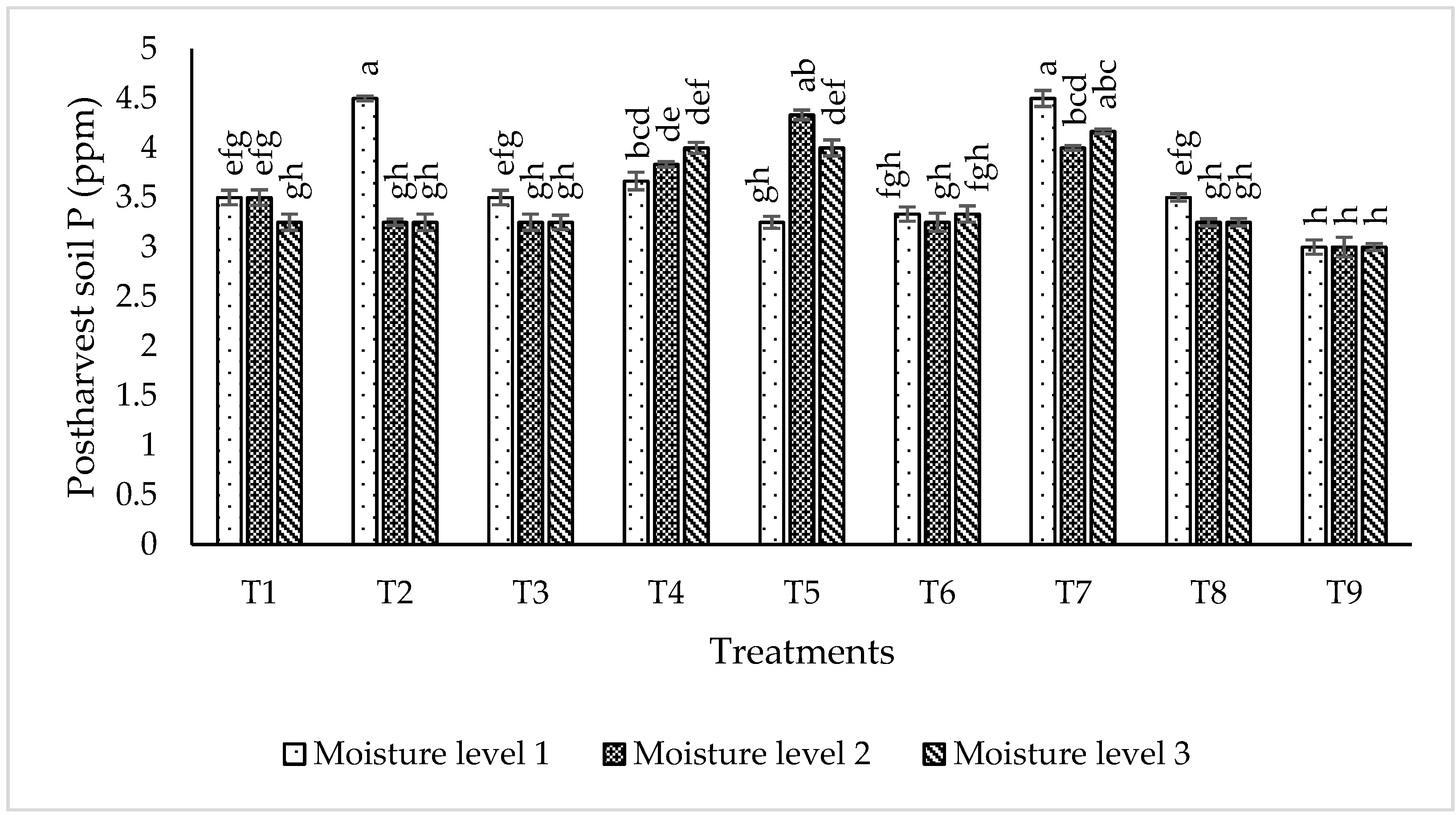
2.6.3. Post-Harvest Soil Nutrient Status
3. Discussion
4. Material and Methods
4.1. Study Area and Sample Collection
4.2. Isolation of Rhizobium and PGPRs
4.3. Morpho-Physiological Characterization of Isolated Strains
4.4. Biochemical Characterization of Isolates
4.5. Growth Chamber Experiment for Screening of Isolates
4.6. Molecular Characterization of Selected Strains
4.7. Compatibility of Isolates for Consortia
4.8. Seed Inoculation with Consortia
4.8.1. Isolate Survival in Rhizospheric Soil
4.8.2. Soil Properties
4.8.3. Leaves Proline and Grain Protein Contents
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kavadia, A.; Omirou, M.; Fasoula, D.; Trajanoski, S.; Andreou, E.; Ioannides, I.M. Genotype and soil water availability shape the composition of AMF communities at chickpea early growth stages. Appl. Soil Ecol. 2020, 150, 103443. [Google Scholar] [CrossRef]
- Aalipour, H.; Nikbakht, A.; Etemadi, N.; Rejali, F.; Soleimani, M. Biochemical response and interactions between arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria during establishment and stimulating growth of Arizona cypress (Cupressus arizonica G.) under drought stress. Sci. Hortic. 2020, 261, 108923. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Curá, J.A. Role of beneficial microorganisms and salicylic acid in improving rainfed agriculture and future food safety. Microorganisms 2020, 8, 1018. [Google Scholar] [CrossRef] [PubMed]
- Mancosu, N.; Snyder, R.L.; Kyriakakis, G.; Spano, D. Water scarcity and future challenges for food production. Water 2015, 7, 975–992. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant. Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Zuluaga, M.Y.; Milani, L.K.M.; Gonçalves, A.L.S.; de Martinez, O.A.L. Diversity and plant growth-promoting functions of diazotrophic/N-scavenging bacteria isolated from the soils and rhizospheres of two species of Solanum. PLoS ONE 2020, 15, e0227422. [Google Scholar] [CrossRef]
- FAOSTAT. Production Statistics of Agriculture Crops. 2020. Available online: http://faostat.fao.org/site (accessed on 15 August 2020).
- Aslam, M.; Mehmood, I.A.; Sultan, T.; Ahmad, S. Inoculation approach to legume crops and their production assessment in Pakistan—A review. Pak. J. Biol. Sci. 2000, 3, 193–195. [Google Scholar]
- Kantar, F.; Hafeez, F.Y.; Kumar, B.G.S.; Sundram, S.P.; Tejera, N.A.; Aslam, A.; Bano, A.; Raja, P. Chickpea: Rhizobium management and nitrogen fixation. In Chickpea Breeding and Management; Cromwell Press: Trowbridge, UK, 2007; pp. 179–192. [Google Scholar]
- Khan, N.; Bano, A.; Rahman, M.A.; Rathinasabapathi, B.; Babar, M.A. UPLC-HRMS-based untargeted metabolic profiling reveals changes in chickpea (Cicer arietinum) metabolome following long-term drought stress. Plant. Cell Environ. 2019, 42, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Bano, A. Exopolysaccharide producing rhizobacteria and their impact on growth and drought tolerance of wheat grown under rainfed conditions. PLoS ONE 2019, 14, e0222302. [Google Scholar] [CrossRef] [Green Version]
- Billah, M.; Khan, M.; Bano, A.; Hassan, T.U.; Munir, A.; Gurmani, A.R. Phosphorus and phosphate solubilizing bacteria: Keys for sustainable agriculture. Geomicrobiol. J. 2019, 36, 904–916. [Google Scholar] [CrossRef]
- Heydri, M.M.; Brook, R.M.; Jones, D.L. The role of phosphorus sources on root diameter, root length and root dry matter of barley (Hordeum vulgare L.). J. Plant Nutr. 2019, 42, 1–15. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of phosphatase enzymes in soil. In Phosphorus in Action, Soil Biology, 26th ed.; Bunemann, E., Oberson, A., Frossard, E., Eds.; Springer: Heidelberg, Germany, 2011; pp. 215–243. [Google Scholar]
- Sharma, S.B.; Sayed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springer Plus 2013, 2, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Yadav, N.; Kumar, V.; Kumar, A.; Sayyed, R.Z. Drought tolerance phosphorus solubilizing microbes. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Microorganisms for Sustainability; Springer: Berlin, Germany, 2019; Volume 12. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. growth and yield of field crops grown under drought stress condition is influenced by the application of PGPR. In Field Crops: Sustainable Management by PGPR; Springer: Cham, Switzerland, 2019; pp. 337–349. [Google Scholar]
- Li, Z.; Sheng, Y.; Yang, J.; Burton, E.D. Phosphorus release from coastal sediments: Impacts of the oxidation-reduction potential and sulfide. Mar. Pollut. Bull. 2016, 113, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Sayin, M.; Mermut, A.R.; Tessen, H. Phosphate sorption-desorption characteristics by magnetically separated soil fraction. Soil Sci. Soc. Am. J. 1990, 54, 1248–1304. [Google Scholar]
- Maliha, R.; Samina, K.; Najma, A.; Sadia, A.; Farooq, L. Organic acids production and phosphate solubilization by phosphate solubilizing microorganisms under in vitro conditions. Pak. J. Biol. Sci. 2004, 7, 187–196. [Google Scholar]
- Vurukonda, S.K.P.; Vardharjula, S.; Shrivastava, M.; Skz, A. Enhancement of drought stress tolerance in crops by plant growth promoting Rhizobacteria. J. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Kumar, S.; Glen, S.; Koichiro, T. MEGA-7, Molecular evolutionary genetics version 7.0 for bigger data sets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Naseem, H.; Ahsan, M.; Shahid, M.A.; Khan, N. Exopolysaccharides producing rhizobacteria and their role in plant growth and drought tolerance. J. Basic Microbiol. 2018, 58, 1009–1022. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Role of plant growth promoting rhizobacteria and Ag-nano particle in the bioremediation of heavy metals and maize growth under municipal wastewater irrigation. Int. J. Phytorem. 2016, 18, 211–221. [Google Scholar] [CrossRef]
- Majeed, A.; Abbasi, M.K.; Hameed, S.; Imran, A.; Rahim, N. Isolation and characterization of plant growth promoting rhizobacteria from wheat rhizosphere and their effect on plant growth promotion. Front. Microbiol. 2015, 6, 198. [Google Scholar] [CrossRef] [Green Version]
- Thakur, I.B.; Putatunda, C. In vitro Phosphate Solubilization by Enterobacter spp. Isolated from wheat phosphere. J. Pure Appl. Microb. 2017, 11, 2007–2015. [Google Scholar] [CrossRef]
- Paul, D.; Sinha, S.N. Isolation of Phosphate Solubilizing Bacteriaand Toatal Hetrotrophic actreria from River Water and study of Phosphatase Activity of PSB. Adv. Appl. Sci. Res. 2013, 4, 409–412. [Google Scholar]
- Khan, N.; Bano, A.; Zandi, P. Effects of exogenously applied plant growth regulators in combination with PGPR on the physiology and root growth of chickpea (Cicer arietinum) and their role in drought tolerance. J. Plant. Interact. 2018, 1, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Mishra, S.; Dixit, V.; Agarval, L.; Chohan, P.S.; Nautiyal, C.S. Synergistic effect of Pseudomonas putida and Bacillus amyloliquefaciens ameliorates drought stress in chickpea (Cicer arietinum L). Plant Signal. Behav. 2016, 11, e1071004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romdhane, S.B.; Tajini, F.; Tarbelsi, M.; Aouani, M.E.; Mohamdi, R. Competition for nodule formation between induced strains of Mesorhizobium ciceri and native rhizobia nodulating chickpea in Tunisia. World J. Microbiol. Biotech. 2007, 23, 1195–1201. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Rhizobacteria and Abiotic Stress Management. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Springer: Singapore, 2019; pp. 65–80. [Google Scholar]
- Dey, R.; Pal, K.K.; Bhatt, D.M.; Chauhan, S.M. Growth and yield enhancement of peanut (Arachis hypogaea L.) by application of plant growth-promoting rhizobacteria. Microbiol. Res. 2004, 159, 371–394. [Google Scholar] [CrossRef]
- Akhtar, M.S.; Siddiqui, Z.A. Effects of phosphate solubilizing microorganisms and Rhizobium sp. on the growth, nodulation, yield and root-rot disease complex of chickpea under field conditions. Afr. J. Biotechnol. 2009, 15, 3489–3496. [Google Scholar]
- Gull, M.; Hafeez, G.Y.; Saleem, M.; Malik, K.A. Phosphorus uptake and growth promotion of chickpea by co-inoculation of mineral PSB and a mixed rhizobial culture. Aust. J. Exp. Agric. 2004, 44, 623–628. [Google Scholar] [CrossRef]
- Marasco, R.; Roli, E.; Ethoumi, B.; Vigami, G.; Mapelli, F. A drought resistantace- promoting microbiome is selected by root system under drought farming. PLoS ONE 2012, 7, e48479. [Google Scholar] [CrossRef]
- Verma, J.P.; Yadav, J.; Tiwari, K.N. Enhancement in nodulation and yield of chickpea by co-inoculation by indigenous Mesorhizobium sp. And plant growth promoting rhizobacteria Eastern Uttar Pradesh. Commun. Soil Sci. Plant Anal. 2012, 43, 621–640. [Google Scholar] [CrossRef]
- Mandhurendra, N.P. Changes in certain organic metabolites during seedling growth of chickpea under salt stress and along with exogenous proline. Leg Res. 2009, 30, 41–44. [Google Scholar]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinative Bacteriology, 9th ed.; Williams and Wilkins: Baltimore, MS, USA, 1994. [Google Scholar]
- Mainatis, T.; Fritsch, E.F.; Sambrook, J. Molecular Cloning, A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1982. [Google Scholar]
- Smibert, R.M.; Kreig, N.R. Phenotypic characterization. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R., Wood, W., Krieg, N., Eds.; ASM Press: Washington, DC, USA, 1994. [Google Scholar]
- Vincent, J.M. A Manual for Practical Study of Root Nodule Bacteria (IBP Handbook 15); Blackwell’s Scientific Publication: Oxford, UK, 1970. [Google Scholar]
- Hussain, A.; Kamran, M.A.; Javed, M.T.; Hayat, K.; Farooq, M.A.; Ali, N.; Chaudhary, H.J. Individual and combinatorial application of Kocuriarhizophila and citric acid on phytoextraction of multi-metal contaminated soils by Glycine max L. Environ. Exp. Bot. 2019, 159, 23–33. [Google Scholar] [CrossRef]
- Dinesh, R.; Anandaraj, M.; Kumar, A.; Bini, Y.K.; Subila, K.P.; Aravind, R. Isolation, characterization, and evaluation of multi-trait plant growth promoting rhizobacteria for their growth promoting and disease suppressing effects on ginger. Microbiol. Res. 2015, 173, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of blue agar CAS assay for siderophore detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Kumar, A.; Devi, S.; Patil, S.; Payal, C.; Nagi, S. Isolation, screening and characterization of bacteria from Rhizospheric soils for different plant growth promotion (PGP) activities as in vitro study. Recent. Res. Sci. Technol. 2012, 4, 1–5. [Google Scholar]
- Qureshi, M.A.; Ahmed, Z.A.; Akhtar, N.; Iqbal, A.; Mujeeb, F.; Shakir, M.A. Role of phosphate solubilizing bacteria (PSB) in enhancing P availability and promoting cotton growth. J. Anim. Plant. Sci. 2012, 22, 204–210. [Google Scholar]
- Macfaddin, J.F. Biochemical Tests for Identification of Medical Bacteria, 3rd ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000. [Google Scholar]
- Ajayi, A.O.; Fagade, O.E. Growth pattern and structural nature of amylases produced by some bacillus species in starchy substrates. Afr. J. Biotechnol. 2006, 5, 440–449. [Google Scholar]
- Adinarayana, K.; Ellaih, P.; Parasde, D.S. Purification and partial characterization of thermostable scrine alkaline protease from a newly isolated Bacillus subtilis. AAPS PharmSciTech 2003, 4, 440–448. [Google Scholar] [CrossRef] [Green Version]
- Etesami, H.; Hosseini, H.M.; Alikhani, H.A. Bacterial biosynthesis of 1-aminocyclopropane-1-caboxylate (ACC) deaminase, a useful trait to elongation and endophytic colonization of the roots of rice under constant flooded conditions. Physiol. Mol. Biol. Plants 2014, 20, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Honma, M.; Shimomura, T. Metabolism of 1-aminocyclopropane-1-carboxylic acid. Agric. Biol. Chem. 1978, 42, 1825–1831. [Google Scholar]
- Subair, H. Isolation and screening bacterial exopolysaccharide (EPS) from potato 516 rhizosphere in highland and the potential as a producer indole acetic acid (IAA). Procedia Food. Sci. 2015, 3, 74–81. [Google Scholar]
- Hashmi, I.; Christophe, P.; Al-Dourobi, A.; Frederic, S.; Priscilla, D.; Thomas, J.; Pilar, J.; Saskia, B. Comparison of the plant growth-promotion performance of a consortium of Bacilli inoculated as endospores or as vegetative cells. FEMS Microbiol. Ecol. 2019, 95, 11. [Google Scholar] [CrossRef] [PubMed]
- Bache, B.; Chesworth, W.; Gessa, C.; Lewis, D.T. Bulk density. In Encyclopedia of Soil Science and Earth Sciences Series; Chestworth, W., Ed.; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- NASA, Glenn Research Center. Nose Cone Volumes of Frustum; NASA, Glenn Research Center: Cleveland, OH, USA, 1968.
- Molina-Romero, D.; Baez, A.; Quintero-Hernández, V.; Castañeda-Lucio, M.; Fuentes-Ramírez, L.E.; Bustillos-Cristales, M.R. Rodríguez-Andrade, O.; Morales-García, Y.E.; Munive, A.; Muñoz-Rojas, J. Compatible bacterial mixture, tolerant to desiccation, improves maize plant growth. PLoS ONE 2017, 12, e0187913. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.F.; Curl, E.A. Methods for Research on the Ecology of Soil-Borne Plant Pathogens; Burgess Publishing Company: Minneapolis, MN, USA, 1972. [Google Scholar]
- Koehler, F.E.; Moudre, C.D.; McNeal, B.L. Laboratory Manual for Soil Fertility; Washington State University: Pulman, WA, USA, 1984. [Google Scholar]
- Walkley, A.; Black, C.A. A critical examination of a rapid method for determination of organic carbon in soils. Effect of variations in digestion conditions and organic soil constituents. Soil Sci. 1934, 63, 251–263. [Google Scholar] [CrossRef]
- Olsen, S.R.; Sommers, L.E.; Page, A.L. Methods of Soil Analysis: Part 2; Soil Science Society of America, Inc.: Madison, WI, USA; American Society of Agronomy, Inc.: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Jackson, M.L. Nitrogen determination from soils and plant tissues. In Soil Chemical Analysis; Constable and Company Ltd.: London, UK, 1962; pp. 183–204. [Google Scholar]
- Soltonpour, P.N.; Schawab, A.P. A new soil test for simultaneous extraction of macro and micronutrient on alkaline soils. Commun. Soil Sci. Plant. Anal. 1977, 8, 195–207. [Google Scholar] [CrossRef]
- McClean, E.O. Soil pH and lime requirement. In Methods of Soil Analysis Part 2, 2nd ed.; Page, A.L., Miller, R.D., Keeney, D.R., Eds.; Agronomy: Madison, WI, USA, 1982; pp. 199–208. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dicky, D. Principles and Procedures of Statistics. A Biometrical Approach, 3rd ed.; McGraw-Hill Book Company, Inc.: New York, NY, USA, 1997. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Strain | Identified Name | Accession No. | Sample Collection Site | Gram Reaction | Bacterial Colony Shape | Form | Elevation | Color | Margin |
|---|---|---|---|---|---|---|---|---|---|
| RP-08 | Enterobacter cloacae | MN599995 | Hayderabad Thal | −ve | Round | Smooth | Raised | Yellowish | Entire |
| RP-01 | Bacillus subtilis | MN599716 | Mankaira | +ve | Irregular | Dry/rough | Umbonate | Yellowish | Undulate |
| RS-15 | Providencia vermicola | MN601273 | Rahdari | −ve | Round | Moist/Round | Convex | Light yellow | Entire |
| RS-14 | Bacillus mojavensis | MN599703 | Pulses research farm Kaloor kot | −ve | Round | Moist | Convex | Yellowish | Undulate |
| RZ-11 | Mesorhizobium ciceri | MN601357 | AZRI Bhakker | −ve | Round | Sticky | Raised | White | Entire |
| RZ-22 | Mesorhizobium ciceri | MN601741 | Chobara | −ve | Round | Sticky | Raised | White | Entire |
| Isolate Strain | Isolation From | IAA Test | HCN Test | Siderophore Test | Phosphate Solubilization | Ammonia Test | Amylase Test | Protease Test | Catalase Test | ACC-Deaminase | EPS |
|---|---|---|---|---|---|---|---|---|---|---|---|
| RP-08 | Rhizoplane soil | +++ | − | + | +++ | − | − | + | + | + | + |
| RP-01 | Rhizoplane soil | ++ | − | ++ | +++ | − | + | + | + | + | + |
| RS-15 | Rhizoplane soil | +++ | − | ++ | − | − | + | + | + | + | + |
| RS-14 | Rhizoplane soil | ++ | − | + | − | − | + | + | + | + | + |
| RZ-11 | Nodules | − | − | − | − | +++ | − | − | − | − | − |
| RZ-22 | Nodules | − | − | − | − | +++ | − | − | − | − | − |
| Isolate Strain Code/Treatment | Bacterial Strains | ACC-Deaminase (μM/mg Protein/h) | Exopolysaccharide (mg/mL) | IAA Production (μg/mL) | Phosphate Solubilization (ug/L) |
|---|---|---|---|---|---|
| RP-08 | Enteroabacter cloacae | 0.84 ± 0.016 | 0.80 ± 0.030 | 86 ± 1.99 | 13.4 ± 0.89 |
| RP-01 | Bacillus subtilis | 0.77 ± 0.024 | 0.74 ± 0.032 | 79 ± 1.60 | 14.2 ± 0.68 |
| RS-15 | Providencia vermicola | 0.69 ± 0.012 | 0.68 ± 0.038 | 71 ± 1.80 | ND |
| RS-14 | Bacillus mojavensis | 0.66 ± 0.022 | 0.61 ± 0.062 | 63 ± 1.40 | ND |
| RZ-11 | Mesorhizobium ciceri | ND | ND | ND | ND |
| RZ-22 | Mesorhizobium ciceri | ND | ND | ND | ND |
| T1 | RZ22 + RP01 + RS15, | 1.3 ± 0.012 | NT | 121 ± 2.30 | 14.2 ± 0.68 |
| T2 | RZ11 + RP01 + RS14, | 2.6 ± 0.011 | NT | 177 ± 2.60 | 14.2 ± 0.68 |
| T3 | RZ11 + RP08 + RS14, | 1.5 ± 0.011 | NT | 138 ± 1.50 | 13.4 ± 0.89 |
| T4 | RZ11 + RP08 + RS15, | 2.4 ± 0.015 | NT | 164 ± 2.10 | 13.4 ± 0.89 |
| T5 | RZ11 + RP01 + RS15 | 2.1 ± 0.017 | NT | 146 ± 2.20 | 14.2 ± 0.68 |
| T6 | RZ22 + RP08 + RS14 | 1.4 ± 0.019 | NT | 133 ± 1.60 | 13.4 ± 0.89 |
| T7 | RZ22 + RP08 + RS15 | 2.5 ± 0.014 | NT | 171 ± 2.10 | 13.4 ± 0.89 |
| T8 | RZ22 + RP01 + RS14 | 1.5 ± 0.011 | NT | 137 ± 1.70 | 14.2 ± 0.68 |
| Treatment (Consortium) | Moisture Level | No. Nodules Plant−1 | Plant Height (cm) | No. Pods Pot−1 | Root Length (cm) | 100 Grain Weight (g) | Biological Yield Pot−1 (g) | Economic Yield Pot−1 (g) | Harvest Index % | Proline (mg g−1 dw) |
|---|---|---|---|---|---|---|---|---|---|---|
| T1 | 1 | 5.00 ij | 29.24 hi | 17.00 ij | 27.23 jkl | 26.93 ij | 33.20 m | 14.20 kl | 39.33 cd | 3.7533 hij |
| 2 | 7.67 fgh | 35.84 efg | 19.33 fg | 34.03 d–h | 28.20 h | 45.00 fg | 16.17 ij | 38.33 d | 3.6167 jkl | |
| 3 | 9.44 e | 41.05 bcd | 21.67 d | 39.24 abc | 29.00 d–g | 49.17 cd | 19.00 c–f | 39.67 bcd | 3.4867 lm | |
| T2 | 1 | 12.33 d | 32.37 gh | 20.67 de | 29.61 ijk | 29.83 abc | 38.93 j | 17.90 fgh | 44.67 ab | 4.7967 a |
| 2 | 23.11 a | 37.59 def | 28.00 a | 42.03 ab | 28.37 gh | 45.01 fg | 20.08 bc | 44.00 abc | 4.2633 de | |
| 3 | 22.45 a | 44.07 ab | 24.33 c | 36.35 c–f | 29.10 c–g | 49.37 c | 20.57 b | 44.00 abc | 4.1000 ef | |
| T3 | 1 | 4.11 j | 28.22 i | 16.67 ijk | 26.77 kl | 25.67 lm | 31.60 no | 14.43 kl | 43.67 abc | 3.6833 ijk |
| 2 | 7.11 fgh | 35.20 efg | 18.33 gh | 32.64 ghi | 28.70 e–h | 44.00 gh | 18.85 d–g | 43.00 a–d | 3.5400 kl | |
| 3 | 8.56 ef | 40.78 bcd | 20.67 de | 38.40 abc | 29.20 b–f | 48.83 cd | 20.10 bc | 42.67 a–d | 3.4733 lm | |
| T4 | 1 | 6.78 gh | 29.14 hi | 19.67 ef | 27.39 jkl | 26.63 ijk | 36.47 k | 16.47 i | 43.33 a–d | 4.5233 bc |
| 2 | 19.00 b | 42.80 ab | 24.33 c | 39.23 abc | 28.57 fgh | 47.53 e | 20.47 b | 45.33 a | 4.4033 cd | |
| 3 | 23.89 a | 45.83 a | 25.67 b | 42.57 a | 29.90 ab | 52.07 a | 22.70 a | 43.33 a–d | 4.2700 de | |
| T5 | 1 | 5.00 ij | 29.80 hi | 17.33 hi | 27.04 kl | 29.33 b–e | 34.37 lm | 14.80 k | 43.00 a–d | 4.0900 ef |
| 2 | 11.89 d | 41.43 bcd | 23.33 c | 38.03 bcd | 29.87 ab | 45.00 fg | 19.90 bcd | 43.00 a–d | 3.9233 fgh | |
| 3 | 11.56 d | 36.13 efg | 19.67 ef | 33.28 e–h | 29.63 a–d | 48.00 de | 19.73 bcd | 43.00 a–d | 3.7167 ijk | |
| T6 | 1 | 3.78 j | 26.79 ij | 16.33 ijk | 26.03 kl | 24.37 n | 30.73 o | 13.60 l | 40.00 bcd | 3.9733 fg |
| 2 | 6.11 hi | 34.14 fg | 18.33 gh | 32.95 f–i | 25.97 klm | 45.00 fg | 19.00 c–f | 41.67 a–d | 3.7533 hij | |
| 3 | 7.56 fgh | 38.40 cde | 18.33 gh | 37.13 cde | 27.23 i | 49.80 bc | 18.00 fgh | 41.00 a–d | 3.6267 jkl | |
| T7 | 1 | 6.11 hi | 28.40 hi | 18.33 gh | 28.33 jkl | 28.57 fgh | 35.20 l | 15.30 jk | 43.33 a–d | 4.6067 b |
| 2 | 23.56 a | 43.53 ab | 29.00 a | 42.47 a | 29.43 a–e | 52.87 a | 23.17 a | 41.67 a–d | 4.5133 bc | |
| 3 | 14.56 c | 42.04 abc | 24.33 c | 36.87 c–f | 30.13 a | 50.00 bc | 19.87 bcd | 42.00 a–d | 4.1733 e | |
| T8 | 1 | 4.00 j | 27.78 ij | 16.67 ijk | 26.30 kl | 24.00 n | 32.00 n | 14.17 kl | 41.33 a–d | 3.8600 ghi |
| 2 | 6.67 gh | 34.39 fg | 18.67 fg | 31.27 hij | 25.36 m | 46.13 f | 19.30 cde | 39.67 bcd | 3.3333 mn | |
| 3 | 8.22 efg | 40.40 bcd | 18.33 gh | 36.24 c–f | 26.30 jkl | 50.83 b | 18.30 efg | 41.67 a–d | 3.2500 n | |
| T9 | 1 | 1.22 k | 22.43 k | 16.00 jk | 24.70 l | 23.07 o | 26.49 p | 11.92 m | 43.00 a–d | 1.4067 o |
| 2 | 1.44 k | 24.07 jk | 15.67 k | 24.90 l | 24.53 n | 40.73 i | 17.10 hi | 42.67 a–d | 1.2533 op | |
| 3 | 1.44 k | 24.06 jk | 16.00 jk | 25.13 l | 25.73 lm | 43.37 h | 17.80 gh | 42.67 a–d | 1.1533 p | |
| LSD (0.05) | 1.63 | 4.00 | 1.31 | 4.17 | 0.77 | 1.17 | 1.14 | 5.15 | 0.1845 | |
| Treatment (Consortium) | Moisture Level | N in Frain (%) | P in Grain (%) | Protein in Grain (%) | N-Contents in Shoot (%) | P-Contents in Shoot (%) |
|---|---|---|---|---|---|---|
| T1 | 1 | 4.0600 l | 0.2967 gh | 25.377 hi | 1.5267 f | 0.2267 jkl |
| 2 | 4.0867 kl | 0.3033 fgh | 25.543 ghi | 1.5300 ef | 0.2300 jkl | |
| 3 | 4.1233 hij | 0.3100 ef | 25.770 b–h | 1.5400 def | 0.2400 hij | |
| T2 | 1 | 4.1867 b | 0.3133 def | 26.167 bc | 1.6633 a | 0.2567 gh |
| 2 | 4.1767 bcd | 0.3033 fgh | 26.107 bcd | 1.6533 ab | 0.2867 def | |
| 3 | 4.1833 bc | 0.3167 cde | 26.150 bc | 1.6300 a–e | 0.3167 b | |
| T3 | 1 | 4.0900 k | 0.3067 efg | 25.563 f–i | 1.5300 ef | 0.2300 jkl |
| 2 | 4.1200 hij | 0.3067 efg | 25.750 c–h | 1.5567 b–f | 0.2567 gh | |
| 3 | 4.1667 b–e | 0.3233 bcd | 26.043 b–e | 1.5800 a–f | 0.2800 ef | |
| T4 | 1 | 4.1067 jk | 0.3033 fgh | 25.667 e–h | 1.6567 ab | 0.2433 hij |
| 2 | 4.1367 f–i | 0.3067 efg | 25.857 b–g | 1.6467 abc | 0.2767 ef | |
| 3 | 4.3133 a | 0.3333 b | 26.960 a | 1.6367 a–d | 0.3367 a | |
| T5 | 1 | 4.1133 ijk | 0.3067 efg | 25.710 d–h | 1.5300 ef | 0.2300 jkl |
| 2 | 4.1567 c–f | 0.3233 bcd | 25.980 b–f | 1.5500 c–f | 0.2500 hi | |
| 3 | 4.1700 b–e | 0.3233 bcd | 26.063 b–e | 1.5833 a–f | 0.2833 def | |
| T6 | 1 | 4.1267 g–j | 0.3033 fgh | 25.790 b–h | 1.5367 def | 0.2367 ijk |
| 2 | 4.1533 d–g | 0.3133 def | 25.960 b–g | 1.5700 a–f | 0.2700 fg | |
| 3 | 4.1767 bcd | 0.3233 bcd | 26.107 bcd | 1.5733 a–f | 0.2933 cde | |
| T7 | 1 | 4.1333 f–j | 0.3133 def | 25.833 b–g | 1.5833 a–f | 0.2400 hij |
| 2 | 4.2867 a | 0.3533 a | 26.793 a | 1.6533 ab | 0.3000 bcd | |
| 3 | 4.1900 b | 0.3267 bc | 26.190 b | 1.6100 a–f | 0.3100 bc | |
| T8 | 1 | 4.1433 e–h | 0.2967 gh | 25.900 b–g | 1.5333 ef | 0.2333 ijk |
| 2 | 4.1700 b–e | 0.3167 cde | 26.063 b–e | 1.5567 b–f | 0.2567 gh | |
| 3 | 4.1733 bcd | 0.3233 bcd | 26.083 b–e | 1.5867 a–f | 0.2867 def | |
| T9 | 1 | 3.8900 m | 0.2933 h | 24.313 j | 1.3100 g | 0.2000 m |
| 2 | 3.8967 m | 0.2933 h | 24.353 j | 1.3233 g | 0.2133 lm | |
| 3 | 3.9167 m | 0.3033 fgh | 25.147 i | 1.3233 g | 0.2200 kl | |
| LSD (0.05) | 0.77 | 0.0103 | 0.4249 | 0.1012 | 0.0183 | |
| Treatments | Isolated Strains | Identified Bacteria |
|---|---|---|
| T1 | RZ-22+RP-01+RS-15 | Mesorhizobium ciceri RZ-22 + Bacillus subtilis RP-01 + Providencia vermicola RS-15 |
| T2 | RZ-11+RP-01+RS-14 | Mesorhizobium ciceri RZ-11 + Bacillus subtilis RP-01 + Bacillus mojavensis RS-14 |
| T3 | RZ-11+RP-08+RS-14 | Mesorhizobium ciceri RZ-11 + Enterobacter cloacae RP-08 + Bacillus mojavensis RS-14 |
| T4 | RZ-11+RP-08+RS-15 | Mesorhizobium ciceri RZ-11 + Enterobacter cloacae RP-08 + Providencia vermicola RS-15 |
| T5 | RZ-11+RP-01+RS-15 | Mesorhizobium ciceri RZ-11 + Bacillus subtilis RP-01 + Providencia vermicola RS-15 |
| T6 | RZ-22+RP-08+RS-14 | Mesorhizobium ciceri RZ-22 + Enterobacter cloacae RP-08 + Bacillus mojavensis RS-14 |
| T7 | RZ-22+RP-08+RS-15 | Mesorhizobium ciceri RZ-22 + Enterobacter cloacae RP-08 + Providencia vermicola RS-15 |
| T8 | RZ-22+RP-01+RS-14 | Mesorhizobium ciceri RZ-22 + Bacillus subtilis RP-01 + Bacillus mojavensis RS-14 |
| T9 | Control |
| Characteristics | Value | Characteristics | Value |
|---|---|---|---|
| Sand (%) | 65.3 | Electrical Conductivity (dSm−1) | 0.45 |
| Silt (%) | 20 | Available P (mg/kg) | 3.29 |
| Clay (%) | 14.7 | Available K (mg/kg) | 68 |
| Texture | Sandy loam | Organic Matter (%) | 0.25 |
| Ph | 8.3 | Nitrogen (%) | 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmood Aulakh, A.; Qadir, G.; Hassan, F.U.; Hayat, R.; Sultan, T.; Billah, M.; Hussain, M.; Khan, N. Desert Soil Microbes as a Mineral Nutrient Acquisition Tool for Chickpea (Cicer arietinum L.) Productivity at Different Moisture Regimes. Plants 2020, 9, 1629. https://doi.org/10.3390/plants9121629
Mahmood Aulakh A, Qadir G, Hassan FU, Hayat R, Sultan T, Billah M, Hussain M, Khan N. Desert Soil Microbes as a Mineral Nutrient Acquisition Tool for Chickpea (Cicer arietinum L.) Productivity at Different Moisture Regimes. Plants. 2020; 9(12):1629. https://doi.org/10.3390/plants9121629
Chicago/Turabian StyleMahmood Aulakh, Azhar, Ghulam Qadir, Fayyaz Ul Hassan, Rifat Hayat, Tariq Sultan, Motsim Billah, Manzoor Hussain, and Naeem Khan. 2020. "Desert Soil Microbes as a Mineral Nutrient Acquisition Tool for Chickpea (Cicer arietinum L.) Productivity at Different Moisture Regimes" Plants 9, no. 12: 1629. https://doi.org/10.3390/plants9121629
APA StyleMahmood Aulakh, A., Qadir, G., Hassan, F. U., Hayat, R., Sultan, T., Billah, M., Hussain, M., & Khan, N. (2020). Desert Soil Microbes as a Mineral Nutrient Acquisition Tool for Chickpea (Cicer arietinum L.) Productivity at Different Moisture Regimes. Plants, 9(12), 1629. https://doi.org/10.3390/plants9121629