Toward the Genetic Basis and Multiple QTLs of Kernel Hardness in Wheat



Abstract
:1. Introduction
2. PINs Are the Major Causal Genes for Wheat Kernel Hardness
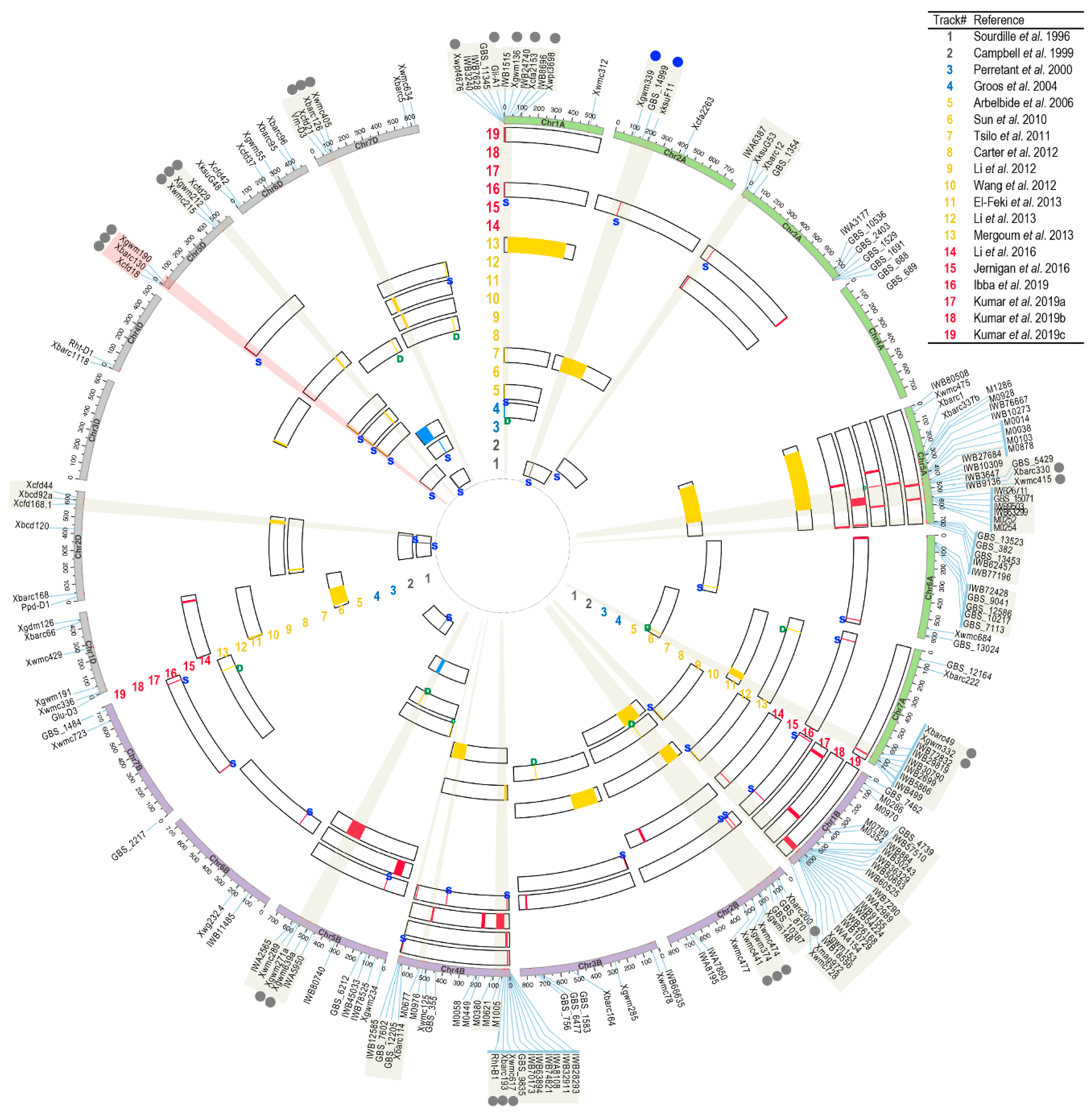
3. Dissecting the Genetic Loci Controlling Wheat Kernel Hardness
4. Technological Improvements Benefit the Mapping of Kernel Hardness
5. Conclusions and Future Perspective
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AFLP | Amplified fragment length polymorphism |
| chr. | Chromosome |
| CRP | Crush response profile |
| CS | Chinese Spring |
| DArT | Diversity array technology |
| DH | Double haploid |
| GBS | Genotyping-by-sequencing |
| GSP | Grain Softness Protein |
| Ha | Hardness |
| HI | Hardness Index |
| HRS | Hard Red Spring |
| HRW | Hard Red Winter |
| HW | Hard White |
| IWGSC | International Wheat Genome Sequencing Consortium |
| KASP | Kompetitive allele-specific PCR |
| MAGIC | Multiparent Advanced Generation Inter-Cross |
| NAM | Nested association mapping |
| NIRS | Near-infrared spectroscopy |
| PCR | Polymerase chain reaction |
| PIN | Puroindoline |
| PINA | Puroindoline A |
| PINB | Puroindoline B |
| QTL | Quantitive trait loci |
| RFLP | Restriction Fragment Length Polymorphism |
| RIL | Recombinant inbred lines |
| SNP | Single-Nucleotide Polymorphism |
| STS | Sequence-Tagged Sites |
| SKCS | Single Kernel Characterization System |
| SSR | Simple Sequence Repeats |
| SRW | Soft Red Winter |
| SW | Soft White |
| USDA | United States Department of Agriculture |
References
- FAO Cereal Supply and Demand Brief. Available online: http://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 13 November 2020).
- IWGSC. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- North Dakota Wheat Commission. World Wheat Production. 2015, North Dakota Wheat Commission. Available online: https://www.ndwheat.com/uploads/6/worldweb-charts.pdf (accessed on 15 August 2020).
- Morris, C.F.; Kiszonas, A.M.; Murray, J.; Boehm, J., Jr.; Ibba, M.I.; Zhang, M.; Cai, X. Re-evolution of durum wheat by introducing the Hardness and Glu-D1 loci. Front. Sustain. Food. Syst. 2019, 3, 103. [Google Scholar] [CrossRef]
- Hourston, J.E.; Ignatz, M.; Reith, M.; Leubner-Metzger, G.; Steinbrecher, T. Biomechanical properties of wheat grains: The implications on milling. J. R. Soc. Interface 2016, 14, 20160828. [Google Scholar] [CrossRef] [PubMed]
- Glenn, G.M.; Johnston, R.K. Moisture-dependent Changes in the Mechanical Properties of Isolated Wheat Bran. J. Cereal Sci. 1992, 15, 223–236. [Google Scholar] [CrossRef]
- Mabille, F.; Gril, J.; Abecassis, J. Mechanical Properties of Wheat Seed Coats. Cereal Chem. 2001, 78, 231–235. [Google Scholar] [CrossRef]
- Antoine, C.; Peyron, S.; Mabille, F.; Lapierre, C.; Bouchet, B.; Abecassis, J.; Rouau, X. Individual Contribution of Grain Outer Layers and Their Cell Wall Structure to the Mechanical Properties of Wheat Bran. J. Agric. Food. Chem. 2003, 51, 2026–2033. [Google Scholar] [CrossRef]
- Greffeuille, V.; Mabille, F.; Rousset, M.; Oury, F.X.; Abecassis, J.; Lullien-Pellerin, V. Mechanical properties of outer layers from near-isogenic lines of common wheat differing in hardness. J. Cereal Sci. 2007, 45, 227–235. [Google Scholar] [CrossRef]
- Martelli, M.R.; Barron, C.; Delaporte, P.; Viennois, G.; Rouau, X.; Sadoudi, A. Pulsed laser ablation: A new approach to reveal wheat outer layer properties. J. Cereal Sci. 2009, 49, 354–362. [Google Scholar] [CrossRef]
- Martelli, M.R.; Barron, C.; Mabille, F.; Rouau, X.; Sadoudi, A. Adherence within biological multilayered systems: Development and application of a peel test on wheat grain peripheral tissues. J. Cereal Sci. 2010, 52, 83–89. [Google Scholar] [CrossRef]
- Chichti, E.; George, M.; Delenne, J.Y.; Radjai, F.; Lullien-Pellerin, V. Nano-mechanical properties of starch and gluten biopolymers from atomic force microscopy. Eur. Polym. J. 2013, 49, 3788–3795. [Google Scholar] [CrossRef]
- Chichti, E.; George, M.; Delenne, J.Y.; Lullien-Pellerin, V. Changes in the starch-protein interface depending on common wheat grain hardness revealed using Atomic Force Microscopy. Plant Sci. 2015, 239, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacRitchie, F. Advances in Cereal Science and Technology; Pomeranz, Y., Ed.; American Association of Cereal Chemists: St. Paul, MN, USA, 1980; p. 271. [Google Scholar]
- Barlow, K.K.; Buttrose, M.S.; Simmonds, D.H.; Vesk, M. The nature of the starch-protein interface in wheat endosperm. Cereal Chem. 1973, 50, 443–454. [Google Scholar]
- Simmonds, D.H.; Barlow, K.K.; Wrigley, C.W. The biochemical basis of grain hardness in wheat. Cereal Chem. 1973, 50, 553–563. [Google Scholar]
- Glenn, G.M.; Saunders, R.M. Physical and structural properties of wheat endosperm associated with grain texture. Cereal Chem. 1990, 67, 176–181. [Google Scholar]
- Glenn, G.M.; Younce, F.L.; Pitts, M.J. Fundamental physical properties characterizing the hardness of wheat endosperm. J. Cereal Sci. 1991, 13, 179–194. [Google Scholar] [CrossRef]
- Finnie, S.M.; Jeannotte, R.; Morris, C.F.; Faubion, J.M. Variation in polar lipid composition among near-isogenic wheat lines possessing different puroindoline haplotypes. J. Cereal Sci. 2010, 51, 66–72. [Google Scholar] [CrossRef]
- Qin, H.; Ma, D.; Huang, X.; Zhang, J.; Sun, W.; Hou, G.; Wang, C.; Guo, T. Accumulation of glycolipids in wheat grain and their role in hardness during grain development. Crop J. 2019, 7, 19–29. [Google Scholar] [CrossRef]
- Bhave, M.; Morris, C.F. Molecular genetics of puroindolines and related genes: Allelic diversity in wheat and other grasses. Plant Mol. Biol. 2008, 66, 205–219. [Google Scholar] [CrossRef]
- Bhave, M.; Morris, C.F. Molecular genetics of puroindolines and related genes: Regulation of expression, membrane binding properties and applications. Plant Mol. Biol. 2008, 66, 221–231. [Google Scholar] [CrossRef]
- Pauly, A.; Pareyt, B.; Fierens, E.; Delcour, J.A. Wheat (Triticum aestivum L. and T. turgidum L. ssp. durum) kernel hardness: I. Current view on the role of Puroindolines and polar lipids. Comp. Rev. Food Sci. Food Saf. 2013, 12, 413–426. [Google Scholar]
- Pauly, A.; Pareyt, B.; Fierens, E.; Delcour, J.A. Wheat (Triticum aestivum L. and T. turgidum L. ssp. durum) kernel hardness: II. Implications for end-product quality and role of Puroindolines therein. Comp. Rev. Food Sci. Food Saf. 2013, 12, 427–438. [Google Scholar] [CrossRef]
- Lullien-Pellerin, V.; Haraszi, R.; Anderssen, R.S.; Morris, C.F. Understanding the mechanics of wheat grain fractionation and the impact of Puroindolines on milling and product quality. In Wheat Quality for Improving Processing and Human Health; Igrejas, G., Ikeda, T., Guzmán, C., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Feiz, L.; Martin, J.M.; Giroux, M.J. Relationship between wheat (Triticum aestivum L.) grain hardness and wet-milling quality. Cereal Chem. 2008, 85, 44–50. [Google Scholar] [CrossRef]
- Pasha, I.; Anjum, F.M.; Morris, C.F. Grain hardness: A major determinant of wheat quality. Food Sci. Tech. Int. 2010, 16, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Oury, F.; Lasme, P.; Michelet, C.; Rousset, M.; Abecassis, J.; Lullien-Pellerin, V. Relationships between wheat grain physical characteristics studied through near-isogenic lines with distinct puroindoline-b allele. Theor. Appl. Genet. 2015, 128, 913–929. [Google Scholar] [CrossRef]
- Salmanowicz, B.P.; Adamski, T.; Surma, M.; Kaczmarek, Z.; Krystkowiak, K.; Kuczyńska, A.; Banaszak, Z.; Ługowska, B.; Majcher, M.; Obuchowski, W.; et al. The relationship between grain hardness dough mixing parameters and bread-making quality in winter wheat. Int. J. Mol. Sci. 2012, 13, 4186–4201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, P.C.; Sobering, D.C. Attempts at standardization of hardness testing of wheat. I. The grinding/sieving (particle size index) method. Cereal Foods World 1986, 31, 362–364. [Google Scholar]
- Norris, K.H.; Hruschka, W.R.; Bean, M.M.; Slaughter, D.C. A definition of wheat hardness using near infrared reflectance spectroscopy. Cereal Foods World 1989, 34, 696–705. [Google Scholar]
- Gaines, C.S.; Finney, P.F.; Fleege, L.M.; Andrews, L.M. Predicting a hardness measurement using the single-kernel characterization system. Cereal Chem. 1996, 73, 278–283. [Google Scholar]
- Symes, K.J. The inheritance of grain harness in wheat as measured by the particle size index. Aust. J. Agric. Res. 1965, 16, 113–123. [Google Scholar] [CrossRef]
- Baker, R.J. Inheritance of kernel hardness in spring wheat. Crop Sci. 1977, 17, 960–962. [Google Scholar] [CrossRef]
- Giroux, M.J.; Morris, C.F. Wheat grain hardness results from highly conserved mutations in the friabilin components puroindoline a and b. Proc. Natl. Acad. Sci. USA 1998, 95, 6262–6266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sourdille, P.; Perretant, M.R.; Charmet, G.; Leroy, P.; Gautier, M.F.; Joudrier, P.; Nelson, J.C.; Sorrells, M.E.; Bernard, M. Linkage between RFLP markers and genes affecting kernel hardness in wheat. Theor. Appl. Genet. 1996, 93, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.G.; Bergman, C.J.; Gualberto, D.G.; Anderson, J.A.; Giroux, M.J.; Hareland, G.; Fulcher, R.G.; Sorrells, M.E.; Finney, P.L. Quantitative trait loci associated with kernel traits in a soft × hard wheat cross. Cereal. Chem. 1999, 39, 1184–1195. [Google Scholar] [CrossRef]
- Greenwell, P.; Schofield, J.D. A starch granule protein associated with endosperm softness in wheat. Cereal Chem. 1986, 63, 379–380. [Google Scholar]
- Greenwell, P. Wheat starch granule proteins and their technological significance. In Proceedings of the 37th Australian Cereal Chemistry Conference, Melbourne, Australia, 9–22 October 1987; Murray, L., Ed.; Cereal Chemistry Division, Royal Australian Chemical Institute: Parkville, Australia, 1987; pp. 100–103. [Google Scholar]
- Schofield, J.D.; Greenwell, P. Wheat starch granule proteins and their technological significance. In Cereals in a European Context; Morton, I.D., Ed.; Ellis Horwood: Chichester, UK, 1987; pp. 407–420. [Google Scholar]
- Greenwell, P.; Schofield, J.D. The chemical basis of grain hardness and softness. In Proceedings of the Wheat End-Use Properties, Proceedings ICC ’89 Symposium, Lahti, Finland, 13–15 June 1989. [Google Scholar]
- Blochet, J.E.; Kaboulou, A.; Compoint, J.P.; Marion, D. Amphiphilic proteins from wheat flour: Specific extraction, structure and lipid binding properties. In Gluten Proteins 1990; Bushuk, W., Tkachuk, R., Eds.; American Association of Cereal Chemists: St. Paul, MN, USA, 1991; pp. 314–325. [Google Scholar]
- Blochet, J.E.; Chevalier, C.; Forest, E.; Pebay-Peyroula, E.; Gautier, M.F.; Joudrier, P.; Pezolet, M.; Marion, D. Complete amino acid sequence of puroindoline, a new basic and cystinerich protein with a unique tryptophan-rich domain, isolated from wheat endosperm by Triton X-114 phase partitioning. FEBS Lett. 1993, 329, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.F.; Greenblatt, G.A.; Malkawi, H.I. Enhanced electrophoretic detection and isolation of friabilin, a starch granule protein. Cereal Chem. 1992, 69, 467–468. [Google Scholar]
- Morris, C.F.; Greenblatt, G.A.; Bettge, A.D.; Malkawi, H.I. Isolation and characterization of multiple forms of friabilin. J. Cereal Sci. 1994, 21, 167–174. [Google Scholar] [CrossRef]
- Jolly, C.; Rahman, S.; Kortt, A.A.; Higgins, T.J.V. Characterisation of the wheat Mr 15,000 “grain-softness protein” and analysis of the relationship between its accumulation in the whole seed and grain softness. Theor. Appl. Genet. 1993, 86, 589–597. [Google Scholar] [CrossRef]
- Jolly, C. The Biochemistry and Molecular Genetics of Grain Softness and Hardness in Wheat, Triticum aestivum. Ph.D. Thesis, Macquarie University, Sydney, Australia, 1991. [Google Scholar]
- Rahman, S.; Jolly, C.J.; Kortt, A.A.; Walloschek, A.; Higgins, T.J. Molecular characterisation of grain softness protein. In Proceedings of the Cereals International 91 Conference, Brisbane, Australia, 9–13 September 1991; Martin, D.J., Wrigley, C.W., Eds.; Cereal Chemistry Division, Royal Chemical Institute: Parkville, Australia, 1991; pp. 288–289. [Google Scholar]
- Greenwell, P. Biochemical studies of endosperm texture in wheat. Chorleywood Digest 1992, 118, 74–76. [Google Scholar]
- Oda, S.; Komae, K.; Yasui, T. Relation between starch granule protein and endosperm softness in Japanese wheat (Triticum aestivum L.) cultivars. Jpn. J. Breed. 1992, 42, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Oda, S.; Schofield, J.D. Characterization of friabilin polypeptides. J. Cereal Sci. 1997, 26, 29–36. [Google Scholar] [CrossRef]
- Morris, C.F. Puroindolines: The molecular genetic basis of wheat grain hardness. Plant Mol. Biol. 2002, 48, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M.-F.; Aleman, M.-E.; Guirao, A.; Marion, D.; Joudier, P. Triticum aestivum puroindolines, two basic cystineyrich seed proteins: cDNA analysis and developmental gene expression. Plant Mol. Biol. 1994, 25, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.; Jolly, C.J.; Skerritt, J.H.; Wallosheck, A. Cloning of a wheat 15-kDa grain softness protein (GSP). GSP is a mixture of puroindoline-like polypeptides. Eur. J. Biochem. 1994, 223, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Giroux, M.J.; Morris, C.F. A glycine to serine change in puroindoline b is associated with wheat grain hardness and low levels of starch-surface friabilin. Theor. Appl. Genet. 1997, 95, 857–864. [Google Scholar] [CrossRef]
- Morris, C.F.; King, G.E.; Allan, R.E.; Simeone, M.C. Identification and characterization of near-isogenic hard and soft hexaploid wheats. Crop Sci. 2001, 41, 211–217. [Google Scholar] [CrossRef]
- Morris, C.F.; Allan, R.E. Registration of hard and soft near-isogenic lines of hexaploid wheat genetic stocks. Crop Sci. 2001, 41, 935–936. [Google Scholar] [CrossRef]
- Jolly, C.J.; Glenn, G.M.; Rahman, S. GSP-1 genes are linked to the grain hardness locus (Ha) on wheat chromosome 5D. Proc. Natl. Acad. Sci. USA 1996, 93, 2408–2413. [Google Scholar] [CrossRef] [Green Version]
- Charles, M.; Tang, H.; Belcram, H.; Paterson, A.; Gornick, P.; Chalhoub, B. Sixty million years in evolution of soft grain trait in grasses: Emergence of the softness locus in the common ancestor of pooideae and ehrhartoideae, after their divergence from panicoideae. Mol. Biol. Evol. 2009, 26, 1651–1661. [Google Scholar] [CrossRef]
- Juhasz, A.; Belova, T.; Florides, C.G.; Maulis, C.; Fischer, I.; Gell, G.; Birinyi, Z.; Ong, J.; Keeble-Gagnere, G.; Maharajan, A.; et al. Genome mapping of seed-borne allergens and immunoresponsive proteins in wheat. Sci. Adv. 2018, 4, eaar8602. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, Y.; Yu, X.; Sun, F.; Yang, G.; He, G. Genomics-enabled analysis of Puroindoline b2 genes identifies new alleles in wheat and related Triticeae species. Int. J. Mol. Sci. 2020, 21, 1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beecher, B.; Bettge, A.; Smidansky, E.; Giroux, M. Expression of wild-type pinB sequence in transgenic wheat complements a hard phenotype. Theor. Appl. Genet. 2002, 105, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Hogg, A.C.; Sripo, T.; Beecher, B.; Martin, J.M.; Giroux, M.J. Wheat puroindolines interact to form friabilin and control wheat grain hardness. Theor. Appl. Genet. 2004, 108, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.M.; Meyer, F.D.; Smidansky, E.D.; Wanjugi, H.; Blechl, A.E.; Giroux, M.J. Complementation of the pina (null) allele with the wild type Pina sequence restores a soft phenotype in transgenic wheat. Theor. Appl. Genet. 2006, 113, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, K.; Giroux, M.J. Expression of wheat puroindoline genes in transgenic rice enhances grain softness. Nat. Biotechnol. 2001, 19, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Martin, J.M.; Beecher, B.; Morris, C.F.; Hannah, L.C.; Giroux, M.J. Seed-specific expression of the wheat puroindoline genes improves maize wet milling yields. Plant Biotechnol. J. 2009, 7, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Mao, X.; Wang, Q.; Zhang, J.R.; Li, X.Y.; Ma, F.Y.; Sun, F.S.; Chang, J.L.; Chen, M.J.; Wang, Y.S.; et al. Overexpression of Puroindoline a gene in transgenic durum wheat (Triticum turgidum ssp. durum) leads to a medium–hard kernel texture. Mol. Breed. 2014, 33, 545–554. [Google Scholar]
- Morris, C.F.; Simeone, M.C.; King, G.E.; Lafiandra, D. Transfer of soft kernel texture from Triticum aestivum to durum wheat, Triticum turgidum ssp. durum. Crop Sci. 2011, 51, 114–122. [Google Scholar] [CrossRef]
- Morris, C.F.; Casper, J.; Kiszonas, A.M.; Fuerst, E.P.; Murray, J.; Simeone, M.C. Soft kernel durum wheat: A new bakery ingredient? Cereal Foods World 2015, 60, 76–83. [Google Scholar] [CrossRef]
- Heinze, K.; Kiszonas, A.M.; Murray, J.C.; Morris, C.F.; Lullien-Pellerin, V. Puroindoline genes introduced into durum wheat reduce milling energy and change milling behavior similar to soft common wheats. J. Cereal Sci. 2016, 71, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.C.; Kiszonas, A.M.; Wilson, J.; Morris, C.F. The effect of soft kernel texture on the milling properties of soft durum wheat. Cereal Chem. 2016, 93, 513–517. [Google Scholar] [CrossRef]
- Quayson, E.T.; Atwell, W.; Morris, C.F.; Marti, A. Empirical rheology and pasting properties of soft-textured durum wheat (Triticum turgidum ssp. durum) and hard-textured common wheat (T. aestivum). J. Cereal Sci. 2016, 69, 252–258. [Google Scholar] [CrossRef]
- Boehm, J.D.; Ibba, M.I.; Kiszonas, A.M.; Morris, C.F. End-use quality of CIMMYT-derived soft-kernel durum wheat germplasm: I. grain, milling, and soft wheat quality. Crop Sci. 2017, 57, 1475–1484. [Google Scholar] [CrossRef]
- Boehm, J.D.; Ibba, M.I.; Kiszonas, A.M.; Morris, C.F. End-use quality of CIMMYT-derived soft-kernel durum wheat germplasm: II. dough strength and pan bread quality. Crop Sci. 2017, 57, 1485–1494. [Google Scholar] [CrossRef]
- Ibba, M.I.; Zhang, M.; Cai, X.; Morris, C.F. Identification of a conserved ph1b-mediated 5DS–5BS crossing over site in soft-kernel durum wheat (Triticum turgidum subsp. durum) lines. Euphytica 2019, 215, 200. [Google Scholar] [CrossRef]
- Lillemo, M.; Chen, F.; Xia, X.C.; William, M.; Peña, R.J.; Trethowan, R.; He, Z.H. Puroindoline grain hardness alleles in CIMMYT bread wheat germplasm. J. Cereal Sci. 2006, 44, 86–92. [Google Scholar] [CrossRef]
- Ali, I.; Sardar, Z.; Rasheed, A.; Mahmood, T. Molecular characterization of the puroindoline-a and b alleles in synthetic hexaploid wheats and in silico functional and structural insights into Pina-D1. J. Theor. Biol. 2015, 376, 1–7. [Google Scholar] [CrossRef]
- Chen, F.; He, Z.H.; Xia, X.C.; Xia, L.Q.; Zhang, X.Y.; Lillemo, M.; Morris, C.F. Molecular and biochemical characterization of puroindoline a and b alleles in Chinese landraces and historical cultivars. Theor. Appl. Genet. 2006, 112, 400–409. [Google Scholar] [CrossRef]
- Kumar, R.; Arora, S.; Singh, K.; Garg, M. Puroindoline allelic diversity in Indian wheat germplasm and identification of new allelic variants. Breed. Sci. 2015, 65, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Sajjad, M.; Wang, J.; Yang, W.; Sun, J.; Li, X.; Zhang, A.; Liu, D. Diversity, distribution of Puroindoline genes and their effect on kernel hardness in a diverse panel of Chinese wheat germplasm. BMC Plant Biol. 2017, 17, 158. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sun, J.Z.; Liu, D.C.; Yang, W.L.; Wang, D.W.; Tong, Y.P.; Zhang, A.M. Analysis of Pina and Pinb alleles in the microcore collections of Chinese wheat germplasm by ecotilling and identification of a novel Pinb allele. J. Cereal Sci. 2008, 48, 836–842. [Google Scholar] [CrossRef] [Green Version]
- Feiz, L.; Beecher, B.S.; Martin, J.M.; Giroux, M.J. In planta mutagenesis determines the functional regions of the wheat puroindoline proteins. Genetics 2009, 183, 853–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feiz, L.; Martin, J.M.; Giroux, M.J. Creation and functional analysis of new Puroindoline alleles in Triticum aestivum. Theor. Appl. Genet. 2009, 118, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhang, F.Y.; Xia, X.C.; Dong, Z.D.; Cui, D.Q. Distribution of puroindoline alleles in bread wheat cultivars of the Yellow and Huai valley of China and discovery of a novel puroindoline a allele without PINA protein. Mol Breed. 2012, 29, 371–378. [Google Scholar] [CrossRef]
- Chen, F.; Li, H.H.; Cui, D.Q. Discovery, distribution and diversity of Puroindoline-D1 genes in bread wheat from five countries (Triticum aestivum L.). BMC Plant Biol. 2013, 13, 125. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, Y.; Zhang, M.; Yu, X.; Hu, R.; Chang, J.; Yang, G.; Wang, Y.; He, G. Diversity of Puroindoline genes and their association with kernel hardness in Chinese wheat cultivars and landraces. Mol. Breed. 2019, 39, 1–13. [Google Scholar] [CrossRef]
- Martin, J.M.; Meyer, F.D.; Morris, C.F.; Giroux, M.J. Pilot scale milling characteristics of transgenic isolines of a hard wheat over-expressing puroindolines. Crop Sci. 2007, 47, 497–506. [Google Scholar] [CrossRef]
- Brites, C.M.; Bagulho, A.S.; Maria, L.B. Effect of wheat puroindoline alleles on functional properties of starch. Eur. Food Res. Technol. 2008, 226, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Lasme, P.; Oury, F.; Michelet, C.; Abecassis, J.; Mabille, F.; Bar, L.; Lullien-Pellerinm, V. A study of puroindoline b gene involvement in the milling behavior and hard-type common wheats. Cereal Chem. 2012, 89, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Kammeraad, J.D.; Giroux, M.J.; Hogg, A.C.; Martin, J.M. Mutagenesis derived puroindoline alleles in Triticum aestivum and their impacts on milling and bread quality. Cereal Chem. 2016, 93, 3125–3144. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Sun, F.; Li, X.; Wang, P.; Yang, G.; He, G. Expression of Puroindoline a in durum wheat affects milling and pasting properties. Front. Plant Sci. 2019, 10, 482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Martin, J.M.; Beecher, B.; Lu, C.; Hannah, L.C.; Wall, M.L.; Altosaar, I.; Giroux, M.J. The ectopic expression of the wheat Puroindoline genes increase germ size and seed oild content in transgenic corn. Plant Mol. Biol. 2010, 74, 353–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, Q.; Li, Y.; Xiao, X.; Sun, S.; Wang, C.; Hu, W.; Feng, J.; Chang, L.; Chen, J.; et al. Coexpression of the high molecular weight glutenin subunit 1Ax1 and puroindoline improves dough mixing properties in durum wheat (Triticum turgidum L. ssp. durum). PLoS ONE 2012, 7, e50057. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, Y.; Sun, F.; Li, X.; Wang, P.; Chang, J.; Wang, Y.; Yang, G.; He, G. Co-expression of high-molecular-weight glutenin subunit 1Ax1 and Puroindoline a (Pina) genes in transgenic durum wheat (Triticum turgidum ssp. durum) improves milling and pasting quality. BMC Plant Biol. 2019, 19, 126. [Google Scholar] [CrossRef]
- Kiszonas, A.M.; Morris, C.F. Wheat breeding for quality: A historical review. Cereal Chem. 2018, 95, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Lullien-Pellerin, V. Both genetic and environmental conditions affect wheat grain texture: Consequences for grain fractionation and flour properties. J. Cereal Sci. 2020, 92, 102917. [Google Scholar] [CrossRef]
- Perretant, M.R.; Cadalen, T.; Charmet, G.; Sourdille, P.; Nicolas, P.; Boeuf, C.; Tixier, M.H.; Branlard, G.; Bernard, S.; Bernard, M. QTL analysis of bread-making quality in wheat using a doubled haploid population. Theor. Appl. Genet. 2000, 100, 1167–1175. [Google Scholar] [CrossRef]
- Arbelbide, M.; Bernardo, R. Mixed-model QTL mapping for kernel hardness and dough strength in bread wheat. Theor. Appl. Genet. 2006, 112, 885–890. [Google Scholar] [CrossRef]
- Sun, X.; Marza, F.; Ma, H.; Carver, B.F.; Bai, G. Mapping quantitative trait loci for quality factors in an inter-class cross of US and Chinese wheat. Theor. Appl. Genet. 2010, 120, 1041–1051. [Google Scholar] [CrossRef]
- Groos, C.; Bervas, E.; Charmet, G. Genetic analysis of grain protein content, grain hardness and dough rheology in a hard x hard bread wheat progeny. J. Cereal Sci. 2004, 40, 93–100. [Google Scholar] [CrossRef]
- Turner, A.S.; Bradburne, R.P.; Fish, L.; Snape, J.W. New quantitative trait loci influencing grain texture and protein content in bread wheat. J. Cereal Sci. 2004, 40, 51–60. [Google Scholar] [CrossRef]
- Tsilo, T.J.; Hareland, G.A.; Simsek, S.; Chao, S.; Anderson, J.A. Genome mapping of kernel characteristics in hard red spring wheat breeding lines. Theor. Appl. Genet. 2010, 121, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Tsilo, T.J.; Simsek, S.; Ohm, J.B.; Hareland, G.A.; Chao, S.; Anderson, J.A. Quantitative trait loci influencing endosperm texture, dough-mixing strength, and bread-making properties of the hard red spring wheat breeding lines. Genome 2011, 54, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Leonard, J.M.; Ross, A.S.; James Peterson, C.; Zemetra, R.S.; Garland-Campbell, K.; Riera-Lizarazu, O. Identification of genetic factors controlling kernel hardness and related traits in a recombinant inbred population derived from a soft X ‘extra-soft’ wheat (Triticum aestivum L.) cross. Theor. Appl. Genet. 2012, 124, 207–211. [Google Scholar] [CrossRef]
- Li, J.; Cui, F.; Ding, A.; Zhao, C.; Wang, X.; Wang, L.; Bao, Y.; Qi, X.; Li, X.; Gao, J.; et al. QTL detection of seven quality traits in wheat using two related recombinant inbred line populations. Euphytica 2012, 183, 207–226. [Google Scholar] [CrossRef]
- Carter, A.H.; Garland-Campbell, K.; Morris, C.F.; Kidwell, K.K. Chromosomes 3B and 4D are associated with several milling and baking quality traits in a soft white spring wheat (Triticum aestivum L.) population. Theor. Appl. Genet. 2012, 124, 1079–1096. [Google Scholar] [CrossRef]
- El-Feki, W.M.; Byrne, P.F.; Reid, S.D.; Lapitan, N.L.; Haley, S.D. Quantitative trait locus mapping for end-use quality traits in hard winter wheat under contrasting soil moisture levels. Crop Sci. 2013, 53, 1953–1967. [Google Scholar] [CrossRef]
- Li, H.M.; Liang, H.; Tang, Z.; Zhang, H.; Yan, B.; Ren, Z. QTL Analysis for grain pentosans and hardness index in a Chinese 1RS.1BL× non-1RS.1BL wheat cross. Plant Mol. Biol. Rep. 2013, 31, 477–484. [Google Scholar] [CrossRef]
- Mergoum, M.; Harilal, V.E.; Simsek, S.; Alamri, M.S.; Schatz, B.G.; Kianian, S.F.; Elias, E.; Kumar, A.; Bassi, F.M. Agronomic and quality QTL mapping in spring wheat. J. Plant Breed. Genet. 2013, 1, 19–33. [Google Scholar]
- Li, C.; Bai, G.; Chao, S.; Carver, B.; Wang, Z. Single nucleotide polymorphisms linked to quantitative trait loci for grain quality traits in wheat. Crop J. 2016, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Boehm, J.D., Jr.; Ibba, M.I.; Kiszonas, A.M.; See, D.R.; Skinner, D.Z.; Morris, C.F. Genetic analysis of kernel texture (grain hardness) in a hard red spring wheat (Triticum aestivum L.) bi-parental population. J. Cereal Sci. 2018, 79, 57–65. [Google Scholar] [CrossRef]
- Kumar, N.J.; Orenday-Ortiz, J.M.; Kiszonas, A.M.; Boehm, J.D., Jr.; Morris, C.F. Genetic analysis of a unique ‘super soft’ kernel texture phenotype in soft white spring wheat. J. Cereal Sci. 2019, 85, 162–167. [Google Scholar] [CrossRef]
- Kumar, N.; Kiszonas, A.M.; Ibba, M.I.; Morris, C.F. Identification of loci and molecular markers associated with Super Soft kernel texture in wheat. J. Cereal Sci. 2019, 87, 286–291. [Google Scholar] [CrossRef]
- Ibba, M.I.; Kiszonas, A.M.; See, D.R.; Skinner, D.Z.; Morris, C.F. Mapping kernel texture in a soft durum (Triticum turgidum subsp. durum) wheat population. J. Cereal Sci. 2019, 85, 20–26. [Google Scholar] [CrossRef]
- Kumar, A.; Mantovani, E.E.; Simsek, S.; Jain, S.; Elias, E.M.; Mergoum, M. Genome wide genetic dissection of wheat quality and yield related traits and their relationship with grain shape and size traits in an elite × non-adapted bread wheat cross. PLoS ONE 2019, 14, e0221826. [Google Scholar] [CrossRef]
- Wang, G.; Leonard, J.M.; Zitzewitz, J.; James Peterson, C.; Ross, A.S.; Riera-Lizarazu, O. Marker–trait association analysis of kernel hardness and related agronomic traits in a core collection of wheat lines. Mol. Breed. 2014, 34, 177–184. [Google Scholar] [CrossRef]
- Jernigan, K.L.; Goddy, J.V.; Huang, M.; Zhou, Y.; Morris, C.F.; Garland-Campbell, K.A.; Zhang, Z.; Carter, A.H. Genetic dissection of end-use quality traits in adapted soft white winter wheat. Front. Plant Sci. 2018, 9, 271. [Google Scholar] [CrossRef] [Green Version]
- Avni, R.; Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Hale, I.; Mascher, M.; Spannagl, M.; Wiebe, K.; et al. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 2017, 357, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.C.; Gu, Y.; Puiu, D.; Wang, H.; Twardziok, S.O.; Deal, K.R.; Huo, N.; Zhu, T.; Wang, L.; Wang, Y.; et al. Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature 2017, 551, 498–502. [Google Scholar] [CrossRef]
- Zhao, G.; Zou, C.; Li, K.; Wang, K.; Li, T.; Gao, L.; Zhang, X.; Wang, H.; Yang, Z.; Liu, X.; et al. The Aegilops tauschii genome reveals multiple impacts of transposons. Nat. Plants 2017, 3, 946–955. [Google Scholar] [CrossRef]
- Ling, H.; Ma, B.; Shi, X.; Liu, H.; Dong, L.; Sun, H.; Cao, Y.; Gao, Q.; Zheng, S.; Li, Y.; et al. Genome sequence of the progenitor of wheat A subgenome Triticum Urartu. Nature 2018, 557, 424–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hu, X.; Islam, S.; She, M.; Peng, Y.; Yu, Z.; Wylie, S.; Juhasz, A.; Dowla, M.; Yang, R.; et al. New insights into the evolution of wheat avenin-like proteins in wild emmer wheat (Triticum dicoccoides). Proc. Natl. Acad. Sci. USA 2018, 115, 13312–13317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilling, S.; Kennedy, A.; Pan, S.; Jermiin, L.S.; Melzer, R. Genome-wide analysis of MIKC-type MADS-box genes in wheat: Pervasive duplications, functional conservation and putative neofunctionalization. New Phytol. 2020, 225, 511–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marone, D.; Panio, G.; Ficco, D.B.M.; Russo, M.A.; De Vita, P.; Papa, R.; Rubiales, D.; Cattivelli, L.; Mastrangelo, A.M. Characterization of wheat DArT markers: Genetic and functional features. Mol. Genet. Genom. 2012, 287, 741–753. [Google Scholar] [CrossRef] [Green Version]
- Wen, W.; He, Z.; Gao, F.; Liu, J.; Jin, H.; Zhai, S.; Qu, Y.; Xia, X. A high-density consensus map of common wheat integrating four mapping populations scanned by the 90K SNP array. Front. Plant Sci. 2017, 8, 1389. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Soriano, J.M.; Alvaro, F. Discovering consensus genomic regions in wheat for root-related traits by QTL meta-analysis. Sci. Rep. 2019, 9, 10537. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Liu, C.; Wang, D.; St Amand, P.; Bernardo, A.; Li, W.; He, F.; Li, L.; Wang, L.; Yuan, X.; et al. High-resolution genome-wide association study identifies genomic regions and candidate genes for important agronomic traits in wheat. Mol. Plant. 2020. [Google Scholar] [CrossRef] [PubMed]
- Juliana, P.; Poland, J.; Huerta-Espino, J.; Shrestha, S.; Crossa, J.; Crespo-Herrera, L.; Toledo, F.H.; Govindan, V.; Mondal, S.; Kumar, U.; et al. Improving grain yield, stress resilience and quality of bread wheat using large-scale genomics. Nat. Genet. 2019, 51, 1530–1539. [Google Scholar] [CrossRef] [PubMed]
- Chitchi, E.; Carrere, M.; George, M.; Delenne, J.Y.; Lullien-Pellerin, V. A wheat grain quantitative evaluation of vitreousness by light transmission analysis. J. Cereal Sci. 2018, 83, 58–62. [Google Scholar]
- Oury, F.; Lasme, P.; Michelet, C.; Rousset, M.; Dubat, A.; Gardet, O.; Heumze, E.; Rolland, B.; Rousset, M.; Abecassis, J.; et al. Bread wheat milling behavior: Effects of genetic and environmental factors, and modeling using grain mechanical resistance traits. Theor. Appl. Genet. 2017, 130, 929–950. [Google Scholar] [CrossRef]
- Ohm, J.B.; Chung, O.K.; Deyoe, C.E. Single kernel characteristics of hard winter wheats in relation to milling and baking quality. Cereal Chem. 1998, 75, 156–161. [Google Scholar] [CrossRef]
- Martin, J.M.; Frohberg, R.C.; Morris, C.F.; Talbert, L.E.; Giroux, M.J. Milling and bread baking traits associated with puroindoline sequence type in hard red spring wheat. Crop Sci. 2001, 41, 228–234. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.R.; Rousser, R.; Brabec, D.L. Rapid Single Kernel Grain Characterization Ststem. U.S. Patent 5,005,774, 9 April 1991. [Google Scholar]
- Martin, C.R.; Rousser, R.; Brabec, D.L. Device for Singulating Particles. U.S. Patent 5,082,141, 21 January 1992. [Google Scholar]
- Martin, C.R.; Rousser, R.; Brabec, D.L. Development of a single-kernel wheat characterization system. Trans. ASAE 1993, 36, 399–1404. [Google Scholar] [CrossRef]
- Morris, C.F.; Anderson, J.A.; King, G.E.; Bettge, A.D.; Garland-Campbell, K.; Allan, R.E.; Fuerst, E.P.; Beecher, B.S. Characterization of a qnique “Super Soft” kernel trait in wheat. Cereal Chem. 2011, 88, 576–583. [Google Scholar] [CrossRef]
- Yu, J.; Buckler, E.S. Genetic association mapping and genome organization of maize. Curr. Opin. Biotechnol. 2006, 17, 155–160. [Google Scholar] [CrossRef]
- Cavanagh, C.R.; Chao, S.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A.; et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.M.; Winfield, M.O.; Burridge, A.J.; Downie, R.C.; Benbow, H.R.; Barker, G.L.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; et al. Characterization of a wheat breeders’ array suitable for high-throughput SNP genotyping of global accessions of hexaploid bread wheat (Triticum aestivum). Plant Biotechnol. J. 2017, 15, 390–401. [Google Scholar] [CrossRef] [Green Version]
- Rasheed, A.; Xia, X. From markers to genome-based breeding in wheat. Theor. Appl. Genet. 2019, 132, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Dong, Z.; Zhao, L.; Ren, Y.; Zhang, N.; Chen, F. The Wheat 660K SNP array demonstrates great potential for marker-assisted selection in polyploid wheat. Plant Biotech. J. 2020, 18, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Wu, J.; Yang, S.; Jin, Y.; Liu, J.; Yang, M.; Rasheed, A.; Zhang, Y.; Xia, X.; Jing, R.; et al. Genome-wide association analysis of stem water-soluble carbohydrate content in bread wheat. Theor. Appl. Genet. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Shi, X.; Zhao, G.; Qin, M.; Ibba, M.I.; Wang, Y.; Li, W.; Yang, P.; Wu, Z.; Lei, Z.; et al. Identification of novel genomic regions and superior alleles associated with Zn accumulation in wheat using a genome-wide association analysis method. Int. J. Mol. Sci. 2020, 21, 1928. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Chen, L.; Zhang, W.; Yang, L.; Zhu, W.; Li, J.; Liu, Y.; Tong, H.; Fu, L.; Liu, J.; et al. Genome-wide association analysis of Fusarium head blight resistance in Chinese elite wheat lines. Front. Plant Sci. 2020, 11, 206. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Liu, J.; Wen, J.; Nie, X.; Xu, L.; Chen, N.; Li, Z.; Wang, Q.; Zheng, Z.; Li, M.; et al. Frequent intra- and inter-species introgression shapes the landscape of genetic variation in bread wheat. Genome Biol. 2019, 20, 136. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yin, H.; Jiao, C.; Fang, X.; Wang, G.; Li, G.; Ni, F.; Li, P.; Su, P.; Ge, W.; et al. Sympatric speciation of wild emmer wheat driven by ecology and chromosomal rearrangements. Proc. Natl. Acad. Sci. USA 2020, 117, 5955–5963. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, X.; Li, Y.; Xu, J.; Bi, A.; Kang, L.; Chen, H.; Wang, Y.; Wang, Y.; Liu, S.; et al. Convergence within divergence: Insights of wheat adaptation from Triticum population sequencing. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Pont, C.; Leroy, T.; Seidel, M.; Tondelli, A.; Duchemin, W.; Armisen, D.; Lang, D.; Bustos-Korts, D.; Goue, N.; Balfourier, F.; et al. Tracing the ancestry of modern bread wheats. Nat. Genet. 2019, 51, 905–911. [Google Scholar] [CrossRef]
- He, F.; Pasam, R.; Shi, F.; Kant, S.; Keeble-Gagnere, G.; Kay, P.; Forrest, K.; Fritz, A.; Hucl, P.; Wiebe, K.; et al. Exome sequencing highlights the role of wild-relative introgression in shaping the adaptive landscape of the wheat genome. Nat. Genet. 2019, 51, 896–904. [Google Scholar] [CrossRef]
- Ramirez-Gonzalez, R.H.; Borrill, P.; Lang, D.; Harrington, S.A.; Brinton, J.; Venturini, L.; Davey, M.; Jacobs, J.; Van Ex, F.; Pasha, A.; et al. The transcriptional landscape of polyploid wheat. Science 2018, 361, eaar6089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, Q.; Guo, L.; Ma, M.; Zhang, L.; Mao, H.; Wu, B.; Liu, X.; Ramirez-Gonzalez, R.H.; Uauy, C.; Appels, R.; et al. Global transcriptome analysis uncovers the gene co-expression regulation network and key genes involved in grain development of wheat (Triticum aestivum L.). Funct. Integr. Genom. 2019, 19, 853–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrill, P.; Ramirez-Gonzalez, R.; Uauy, C. expVIP: A customisable RNA-seq data analysis and visualisation platform. Plant Physiol. 2016, 170, 2172–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wang, M.; Lin, K.; Xie, Y.; Guo, J.; Ye, L.; Zhuang, Y.; Teng, W.; Ran, X.; Tong, Y.; et al. The bread wheat epigenomic map reveals distinct chromatin architectural and evolutionary features of functional genetic elements. Genome Biol. 2019, 20, 139. [Google Scholar] [CrossRef] [Green Version]
- Ran, X.; Zhao, F.; Wang, Y.; Liu, J.; Zhuang, Y.; Ye, L.; Qi, M.; Cheng, J.; Zhang, Y. Plant Regulomics: A data-driven interface for retrieving upstream regulators from plant multi-omics data. Plant J. 2020, 101, 237–248. [Google Scholar] [CrossRef]
- Gardinar, L.J.; Joynson, R.; Omony, J.; Rusholme-Pilcher, R.; Olohan, L.; Lang, D.; Bai, C.; Hawkesford, M.; Salt, D.; Spannagl, M.; et al. Hidden variation in polyploid wheat drives local adaptation. Genome Res. 2018, 28, 1319–1332. [Google Scholar] [CrossRef] [Green Version]
- Krasileva, K.V.; Vasquez-Gross, H.A.; Howell, T.; Bailey, P.; Paraiso, F.; Clissold, L.; Simmonds, J.; Ramirez-Gonzalez, R.H.; Wang, X.; Borrill, P.; et al. Uncovering hidden variation in polyploid wheat. Proc. Natl. Acad. Sci. USA 2017, 114, E913–E921. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Chen, Z.; Guo, Y.; Sun, H.; Zhang, G.; Kuang, M.; Yang, S.; Li, X.; Diaz de la Garza, R.I.; Gou, J. Isolation of wheat mutants with higher grain phenolics to enhance anti-oxidant potential. Food Chem. 2020, 303, 125363. [Google Scholar] [CrossRef]
- Wilkinson, M.D.; Tosi, P.; Lovegrove, A.; Corol, D.I.; Ward, J.L.; Palmer, R.; Powers, S.; Passmore, D.; Webster, G.; Marcus, S.E.; et al. The Gsp-1 genes encode the wheat arabinogalactan peptide. J. Cereal Sci. 2017, 74, 155–164. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| References | Track# Genet Map 1 | Track# Genom Map 2 | Description of the Mapping Population | Type of Pop. 3 | Pop. Size 4 | Genetic Map Size 5 | Marker Type | No. QTL | Projected No. QTL Genetic Map | Projected No. QTL Genomic Map | 5DS Ha Locus |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sourdille et al., 1996 [37] | 1 | 1 | W-7984 X Opata85 Soft X Hard | RIL | 86 | NM | RFLP | 5 | 5 | 3 | Yes |
| Campbell et al., 1999 [38] | 2 | 2 | NY18 X CC Soft X Hard | RIL | 78 | 1806 | RFLP | 4 | 1 | 4 | Yes |
| Perretant et al., 2000 [100] | 3 | 3 | Courtot X Chinese Spring (CS) Hard X Soft | DH | 169 | ~2900 | RFLP, AFLP, SSR, functional markers | 3 | 3 | 1 | Yes |
| Groos et al., 2004 [103] | 4 | 4 | Renan X Recital Hard X Hard | RIL | 165 | 2722 | RFLP, AFLP, SSR, | 5 6 | 3 | 3 | |
| Arbelbide et al., 2006 [101] | 5 | 5 | 158 F2 crosses made from 80 parental lines, and SSD to generate 373 RILs | RIL | 373 | NM | SSR, functional markers | 2 | 2 | 2 | Yes |
| Sun et al., 2010 [102] | 6 | 6 | Ning7840 X Clark Hard X Soft | RIL | 132 | 2203 | AFLP, SSR | 2; 4; 7 | 4 | 5 | Yes |
| Tsilo et al., 2011 [106] | 7 | 7 | MN98550 X MN99394 Hard X Hard | RIL | 139 | 2505 | SSR, DArT, functional markers | 6 | 6 | 6 | Yes |
| Wang et al., 2012 [107] | 10 | 10 | OS9A X Q36 Soft X Extra Soft | RIL | 164 | 1821 | SSR, DArT, functional markers | 4 | 4 | 4 | |
| Li et al., 2012 [108] | 9 | 9 | Weimai 8 X Jimai20 Weimai 8 X Yangnong19 All three parents are hard kernel | RIL | 485-WJ; 229-WY; | 2855-WJ; 3010-WY; | SSR, STS, SRAP, RAPD, functional markers | 3; 5; 8 | 4 | 6 | |
| Carter et al., 2012 [109] | 8 | 8 | Louise X Penawawa Soft X soft | RIL | 188 | NM | SSR, SNP (only 1), functional marker | 4 | 4 | 4 | |
| El-Feki et al., 2013 [110] | 11 | 11 | CO940610 X Platte Hard X Hard | DH | 185 | 2117 | SSR, STS, DArT | 6 | 6 | 5 | |
| Li et al., 2013 [111] | 12 | 12 | R146 x R97 Hard X Hard | RIL | 103 | NM | SSR | 3 | 3 | 2 | |
| Mergoum et al., 2013 [112] | 13 | 13 | SteeleND X ND735 Hard X Hard | RIL | 129 | 1789 | SSR, DArT | 4 | 4 | 4 | |
| Wang et al., 2014 [119] | - | GWAS population. 94 diverse lines with phenotypes of hard, soft and extra soft kernels | GWAS | 94 | 1193 * | SSR, DArT | 2 | 0 | 0 | ||
| Li et al., 2016 [113] | 14 | 14 | Ning7840 X Clark Hard X Soft | RIL | 127 | 4225 | SSR, SNP | 2; 5; 9 | 4 | 6 | Yes |
| Jernigan et al., 2018 [120] | - | 15 | GWAS population. including 469 lines with historical phenotypes | GWAS | 469 | 15,229 * | SNP | 5 | 0 | 5 | |
| Boehm Jr. et al., 2018 [114] | - | Butte86 X ND2603 Hard X Hard | RIL | 132 | 1813 | SSR, SNP, Functional markers | 4 | na | na | ||
| Ibba et al., 2019 [117] | - | 16 | Creso X Langdon 1-67 8Hard X Soft | RIL | 428 | 8495 ** | SNP, TAS, Functional markers | 2; 24; 10 | 0 | 20 | |
| Kumar et al., 2019a [115] | - | 17 | Alpowa X BC2F5SS163 Soft X Extra Soft | RIL (F3:F5) | 125 | 913 | SNP | 10 | 0 | 8 | |
| Kumar et al., 2019b [116] | 15 | 18 | Alpowa X BC2F5SS163 Soft X Extra Soft | RIL (F6) | 229 | NA | KASP, SSR | 4 | 1 | 4 | |
| Kumar et al., 2019c [118] | - | 19 | ND705 X PI414566 Hard X Hard | RIL | 160 | 4676 | SNP | 7 11 | 0 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, M.; Li, Y. Toward the Genetic Basis and Multiple QTLs of Kernel Hardness in Wheat. Plants 2020, 9, 1631. https://doi.org/10.3390/plants9121631
Tu M, Li Y. Toward the Genetic Basis and Multiple QTLs of Kernel Hardness in Wheat. Plants. 2020; 9(12):1631. https://doi.org/10.3390/plants9121631
Chicago/Turabian StyleTu, Min, and Yin Li. 2020. "Toward the Genetic Basis and Multiple QTLs of Kernel Hardness in Wheat" Plants 9, no. 12: 1631. https://doi.org/10.3390/plants9121631
APA StyleTu, M., & Li, Y. (2020). Toward the Genetic Basis and Multiple QTLs of Kernel Hardness in Wheat. Plants, 9(12), 1631. https://doi.org/10.3390/plants9121631