Research Trends and Methodological Approaches of the Impacts of Windstorms on Forests in Tropical, Subtropical, and Temperate Zones: Where Are We Now and How Should Research Move Forward?




Abstract
:1. Introduction
2. Results
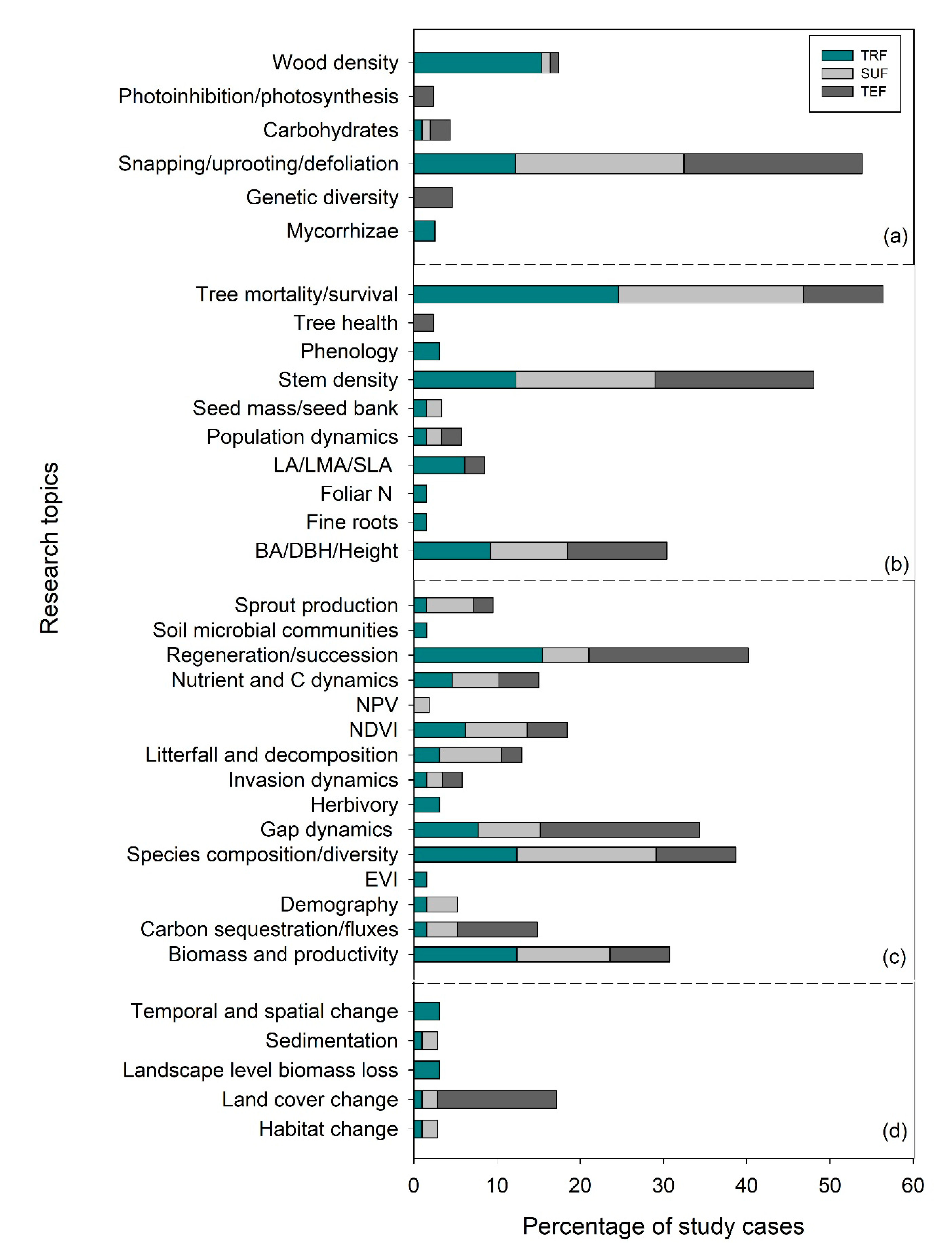
2.1. Research Topics Across Levels of Biological Organization
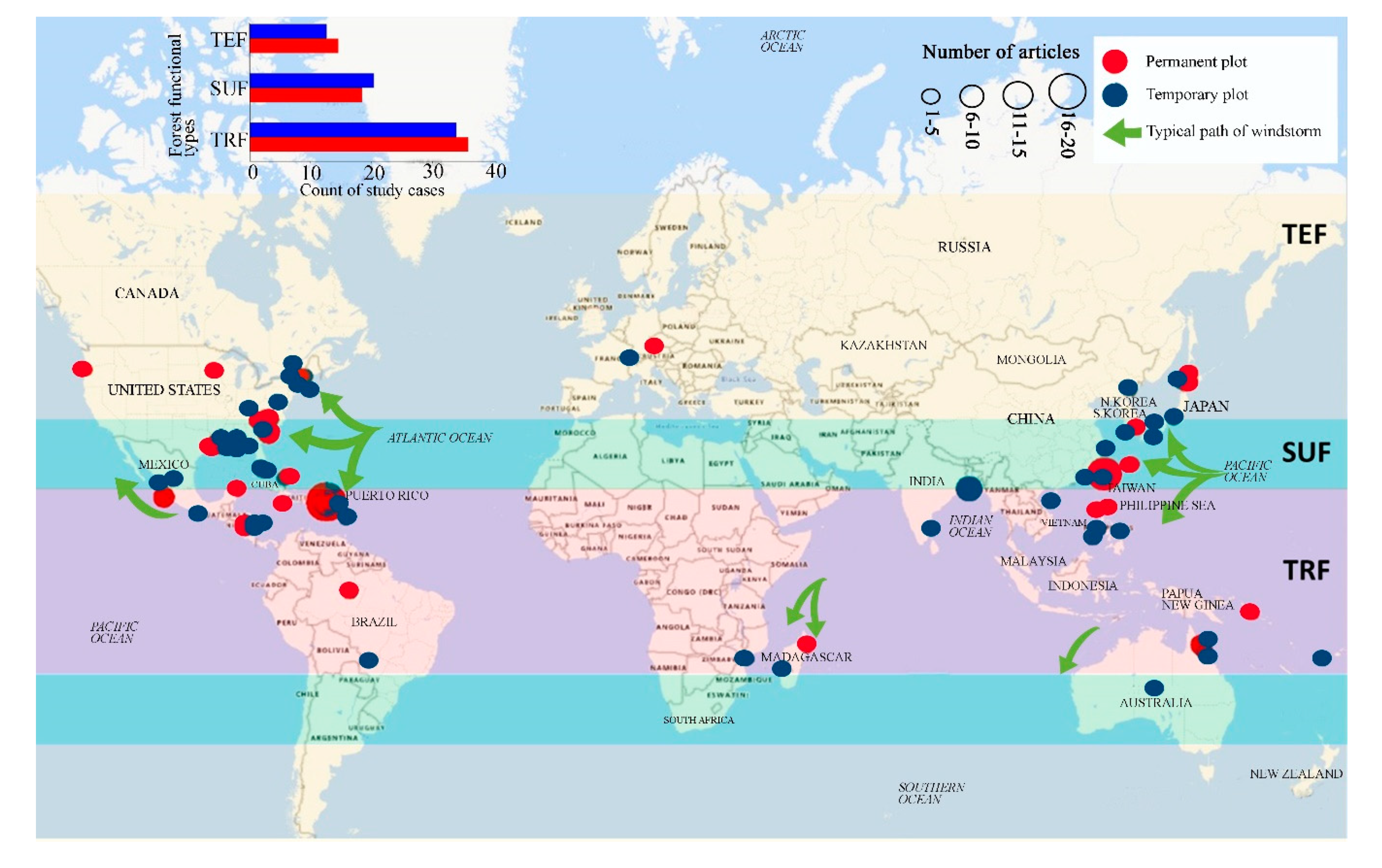
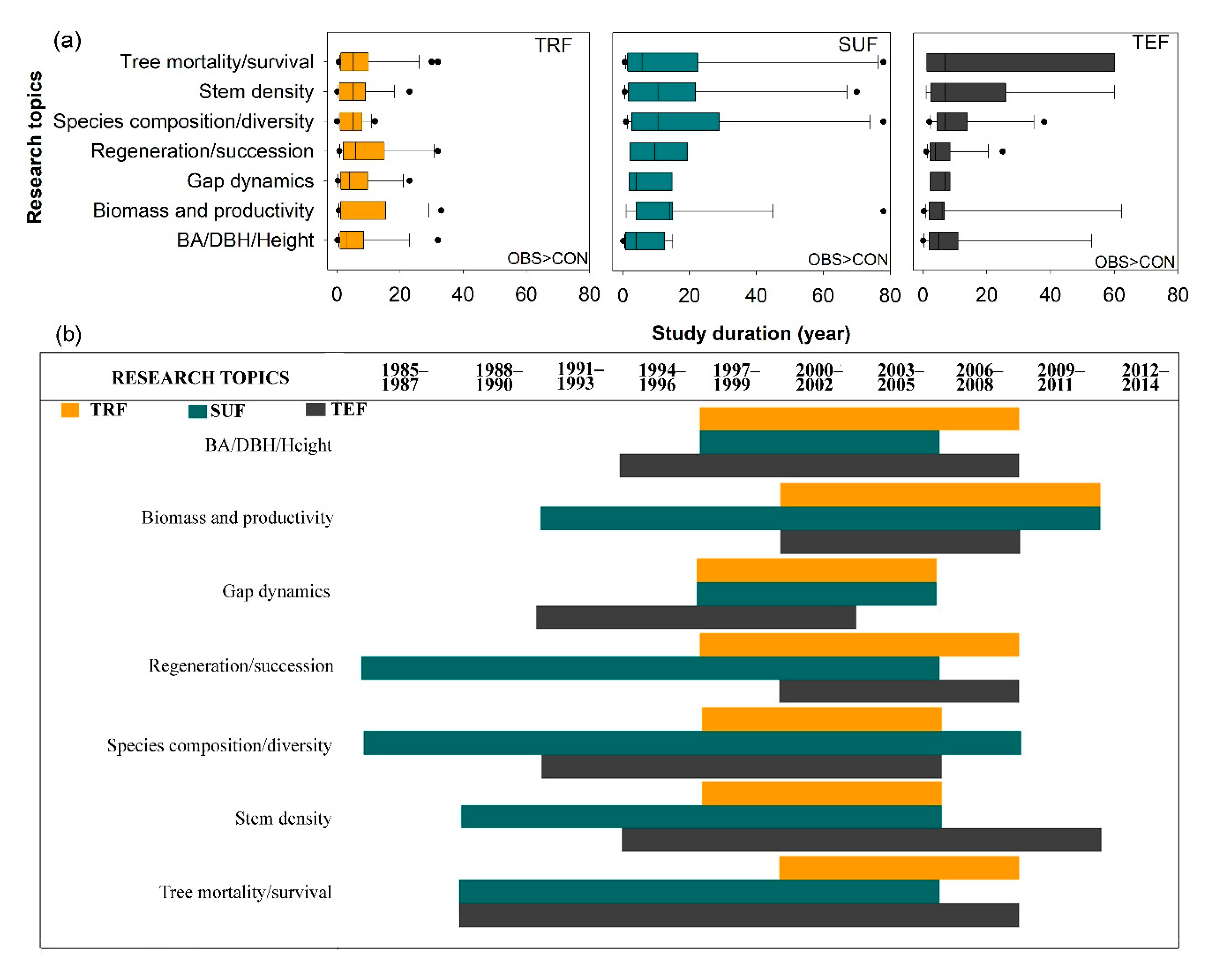
2.2. Study Approaches to Determine Windstorm Effects Across Forest Types
3. Discussion
3.1. Synergistic Effects of Catastrophic Windstorms on Forest Across Levels of Biological Organization
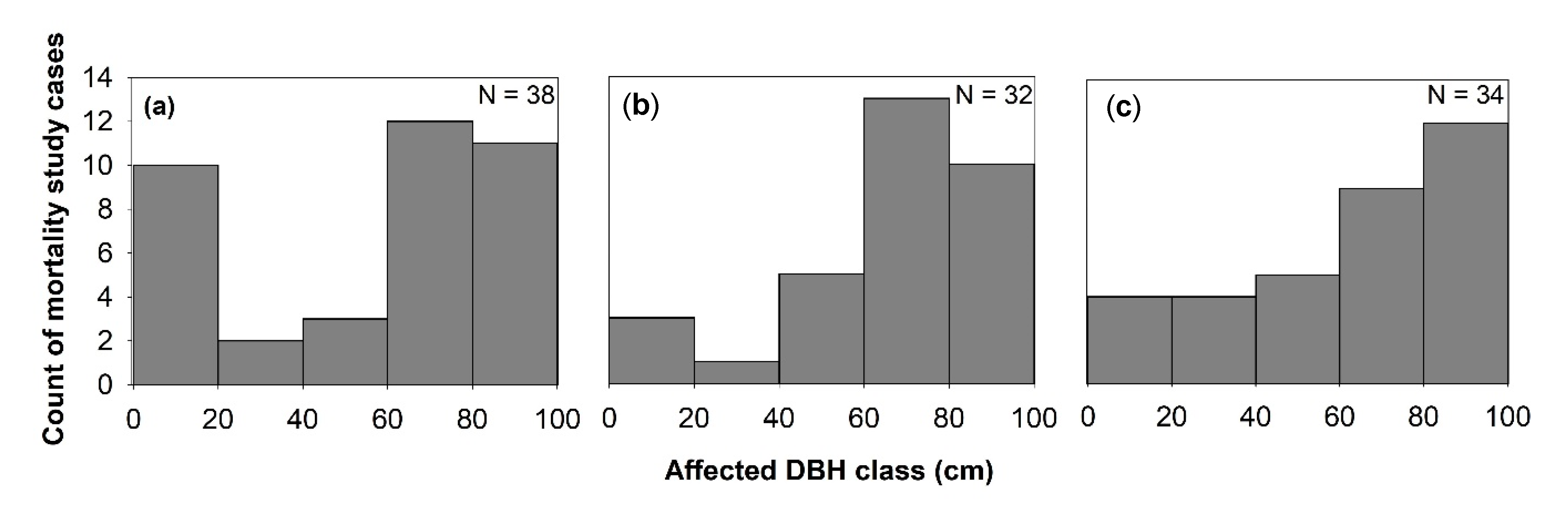
3.1.1. Individual-Level Effects
3.1.2. Species and Population-Level Effects
3.1.3. Community and Ecosystem-Level Effects
3.1.4. Landscape-Level Effects
3.2. Where Are We Now and How Should Research Move Forward?
3.2.1. Knowledge Gaps
3.2.2. Gaps in Methodological Approaches
4. Methodology
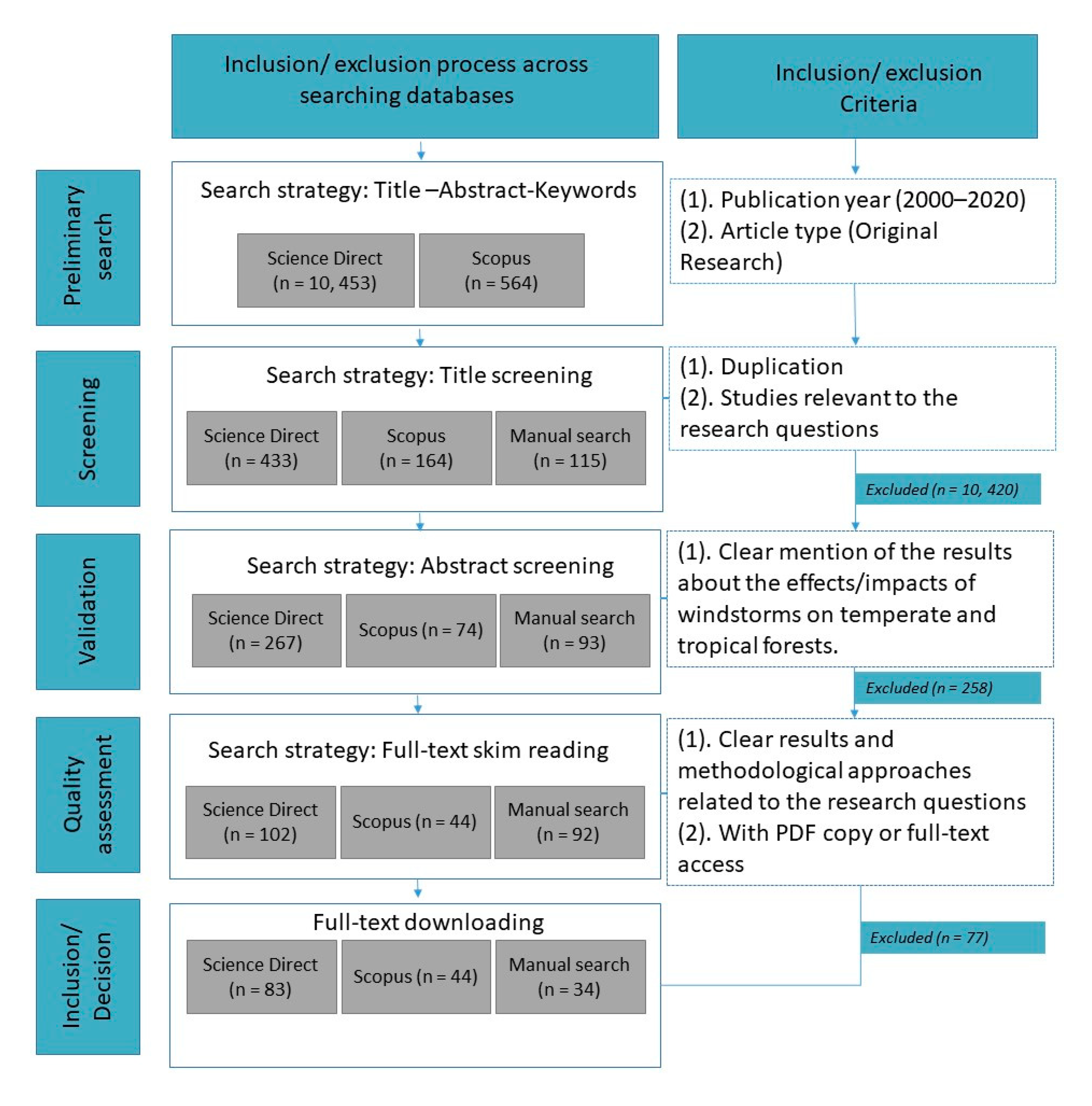
4.1. Data Collection
4.2. Article Selection and Appraisal
4.3. Data Categorization
4.4. Data Analysis and Presentation
4.5. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henkel, T.K.; Chambers, J.Q.; Baker, D.A. Delayed tree mortality and Chinese tallow (Triadica sebifera) population explosion in a Louisiana bottomland hardwood forest following Hurricane Katrina. Forest Ecol. Manag. 2016, 378, 222–232. [Google Scholar] [CrossRef] [Green Version]
- Vozmishcheva, A.S.; Bondarchuk, S.N.; Gromyko, M.N.; Kislov, D.E.; Pimenova, E.A.; Salo, M.A.; Korznikov, K.A. Strong Disturbance Impact of Tropical Cyclone Lionrock (2016) on Korean Pine-Broadleaved Forest in the Middle Sikhote-Alin Mountain Range, Russian Far East. Forests 2019, 10, 1017. [Google Scholar] [CrossRef] [Green Version]
- Frazer, G.W.; Wulder, M.A.; Niemann, K.O. Simulation and quantification of the fine-scale spatial pattern and heterogeneity of forest canopy structure: A lacunarity-based method designed for analysis of continuous canopy heights. Forest Ecol. Manag. 2005, 214, 65–90. [Google Scholar] [CrossRef]
- Weishampel, J.F.; Drake, J.B.; Cooper, A.; Blair, J.B.; Hofton, M. Forest canopy recovery from the 1938 hurricane and subsequent salvage damage measured with airborne LiDAR. Remote Sens. Environ. 2007, 109, 142–153. [Google Scholar] [CrossRef]
- Scalley, T.H.; Scatena, F.N.; Lugo, A.E.; Moya, S.; Ruiz, C.R.E. Changes in Structure, Composition, and Nutrients During 15 Yr of Hurricane-Induced Succession in a Subtropical Wet Forest in Puerto Rico. Biotropica 2010, 42, 455–463. [Google Scholar] [CrossRef]
- Webb, E.L.; Bult, M.V.D.; Fa’aumu, S.; Webb, R.C.; Tualaulelei, A.; Carrasco, L.R. Factors Affecting Tropical Tree Damage and Survival after Catastrophic Wind Disturbance. Biotropica 2013, 46, 32–41. [Google Scholar] [CrossRef]
- Ostertag, R.; Silver, W.L.; Lugo, A.E. Factors affecting mortality and resistance to damage following hurricanes in a rehabilitated subtropical moist forest. Biotropica 2005, 37, 16–24. [Google Scholar] [CrossRef]
- Uriarte, M.; Canham, C.D.; Thompson, J.; Zimmerman, J.K.; Murphy, L.; Sabat, A.M.; Fetcher, N.; Haines, B.L. Natural disturbance and human land use as determinants of tropical forest dynamics: Results from a forest simulator. Ecol. Monogr. 2009, 79, 423–443. [Google Scholar] [CrossRef]
- Unwin, G.L.; Applegate, G.B.; Stocker, G.C.; Nicholson, D.L. Initial effects of tropical cyclone “Winifred” on forests in north Queensland. Proc. Ecol. Soc. Aust. 1988, 15, 283–296. [Google Scholar]
- Vasquez-Yanes, C.; Orozco-Segovia, A. Seed germination of a tropical rain forest pioneer tree (Heliocarpus donnell-smithii) in response to diurnal fluctuation of temperature. Physiol. Plant. 1982, 56, 295–298. [Google Scholar] [CrossRef]
- Walker, L.R. Tree Damage and Recovery from Hurricane Hugo in Luquillo Experimental Forest, Puerto-Rico. Biotropica 1991, 23, 379–385. [Google Scholar] [CrossRef]
- Tzeng, H.-Y.; Wang, W.; Tseng, Y.-H.; Chiu, C.-A.; Kuo, C.-C.; Tsai, S.-T. Tree mortality in response to typhoon-induced floods and mudslides is determined by tree species, size, and position in a riparian Formosan gum forest in subtropical Taiwan. PLoS ONE 2018, 13, e0190832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, W.M.; Peet, R.K.; Decoster, J.K.; Urban, D.L. Tree damage risk factors associated with large, infrequent wind disturbances of Carolina forests. Forestry 2008, 81, 317–334. [Google Scholar] [CrossRef] [Green Version]
- Lugo, A.E. Visible and invisible effects of hurricanes on forest ecosystems: An international review. Austral Ecol. 2008, 33, 368–398. [Google Scholar] [CrossRef]
- Ibanez, T.; Keppel, G.; Menkes, C.; Gillespie, T.W.; Lengaigne, M.; Mangeas, M.; Rivas-Torres, G.; Birnbaum, P. Globally consistent impact of tropical cyclones on the structure of tropical and subtropical forests. J. Ecol. 2019, 107, 279–292. [Google Scholar] [CrossRef]
- Lin, K.C.; Hamburg, S.P.; Wang, L.X.; Duh, C.T.; Huang, C.M.; Chang, C.T.; Lin, T.C. Impacts of increasing typhoons on the structure and function of a subtropical forest: Reflections of a changing climate. Sci. Rep. 2017, 7, 4911. [Google Scholar] [CrossRef] [Green Version]
- Peereman, J.; Hogan, J.A.; Lin, T.C. Landscape Representation by a Permanent Forest Plot and Alternative Plot Designs in a Typhoon Hotspot, Fushan, Taiwan. Remote Sens. 2020, 12, 660. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.C.; Hogan, J.A.; Chang, C.T. Tropical Cyclone Ecology: A Scale-Link Perspective. Trends Ecol. Evol. 2020, 35, 594–604. [Google Scholar] [CrossRef] [Green Version]
- Bender, M.A.; Knutson, T.R.; Tuleya, R.E.; Sirutis, J.J.; Vecchi, G.A.; Garner, S.T.; Held, I.M. Modeled Impact of Anthropogenic Warming on the Frequency of Intense Atlantic Hurricanes. Science 2010, 327, 454–458. [Google Scholar] [CrossRef] [Green Version]
- Altman, J.; Dolezal, J.; Cerny, T.; Song, J.S. Forest response to increasing typhoon activity on the Korean peninsula: Evidence from oak tree-rings. Glob. Change Biol. 2013, 19, 498–504. [Google Scholar] [CrossRef]
- Fisk, J.P.; Hurtt, G.C.; Chambers, J.Q.; Zeng, H.; Dolan, K.A.; Negron-Juarez, R.I. The impacts of tropical cyclones on the net carbon balance of eastern US forests (1851–2000). Environ. Res. Lett. 2013, 8, 045017. [Google Scholar] [CrossRef] [Green Version]
- Kurz, W.A.; Stinson, G.; Rampley, G.J.; Dymond, C.C.; Neilson, E.T. Risk of natural disturbances makes future contribution of Canada’s forests to the global carbon cycle highly uncertain. Proc. Natl. Acad. Sci. USA 2008, 105, 1551–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uriarte, M.; Papaik, M. Hurricane impacts on dynamics, structure and carbon sequestration potential of forest ecosystems in Southern New England, USA. Tellus A 2007, 59, 19–28. [Google Scholar] [CrossRef]
- Dolan, K.A.; Hurtt, G.C.; Chambers, J.Q.; Dubayah, R.O.; Frolking, S.; Masek, J.G. Using ICESat’s Geoscience Laser Altimeter System (GLAS) to assess large-scale forest disturbance caused by hurricane Katrina. Remote Sens. Environ. 2011, 115, 86–96. [Google Scholar] [CrossRef]
- Xi, W.M.; Peet, R.K.; Urban, D.L. Changes in forest structure, species diversity and spatial pattern following hurricane disturbance in a Piedmont North Carolina forest, USA. J. Plant. Ecol. 2008, 1, 43–57. [Google Scholar] [CrossRef]
- Mayer, P.; Brang, P.; Dobbertin, M.; Hallenbarter, D.; Renaud, J.P.; Walthert, L.; Zimmermann, S. Forest storm damage is more frequent on acidic soils. Ann. Forest Sci. 2005, 62, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Peterson, C.J. Within-stand variation in windthrow in southern boreal forests of Minnesota: Is it predictable? Can. J. For. Res. 2004, 34, 365–375. [Google Scholar] [CrossRef]
- Butler, D.W.; Gleason, S.M.; Davidson, I.; Onoda, Y.; Westoby, M. Safety and streamlining of woody shoots in wind: An empirical study across 39 species in tropical Australia. New Phytol. 2012, 193, 137–149. [Google Scholar] [CrossRef]
- Piermattei, A.; von Arx, G.; Avanzi, C.; Fonti, P.; Gartner, H.; Piotti, A.; Urbinati, C.; Vendramin, G.G.; Buntgen, U.; Crivellaro, A. Functional Relationships of Wood Anatomical Traits in Norway Spruce. Front. Plant Sci. 2020, 11, 683. [Google Scholar] [CrossRef]
- Meyer, F.D.; Paulsen, J.; Korner, C. Windthrow damage in Picea abies is associated with physical and chemical stem wood properties. Trees-Struct. Funct. 2008, 22, 463–473. [Google Scholar] [CrossRef]
- Curran, T.J.; Brown, R.L.; Edwards, E.; Hopkins, K.; Kelley, C.; McCarthy, E.; Pounds, E.; Solan, R.; Wolf, J. Plant functional traits explain interspecific differences in immediate cyclone damage to trees of an endangered rainforest community in north Queensland. Austral Ecol. 2008, 33, 451–461. [Google Scholar] [CrossRef]
- Uriarte, M.; Thompson, J.; Zimmerman, J.K. Hurricane Maria tripled stem breaks and doubled tree mortality relative to other major storms. Nat. Commun. 2019, 10, 1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogan, J.A.; Zimmerman, J.K.; Thompson, J.; Uriarte, M.; Swenson, N.G.; Condit, R.; Hubbell, S.; Johnson, D.J.; Sun, I.F.; Chang-Yang, C.H.; et al. The Frequency of Cyclonic Wind Storms Shapes Tropical Forest Dynamism and Functional Trait Dispersion. Forests 2018, 9, 404. [Google Scholar] [CrossRef] [Green Version]
- Gelder, H.A.V.; Poorter, L.; Sterck, F.J. Wood mechanics, allometry, and life-history variation in a tropical rain forest tree community. New Phytol. 2006, 171, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Iida, Y.; Poorter, L.; Sterck, F.J.; Kassim, A.R.; Kubo, T.; Potts, M.D.; Kohyama, T.S. Wood density explains architectural differentiation across 145 co-occurring tropical tree species. Funct. Ecol. 2011, 26, 274–282. [Google Scholar] [CrossRef]
- Ribeiro, G.H.P.M.; Chambers, J.Q.; Peterson, C.J.; Trumbore, S.E.; Marra, D.M.; Wirth, C.; Cannon, J.B.; Negron-Juarez, R.I.; Lima, A.J.N.; de Paula, E.V.C.M.; et al. Mechanical vulnerability and resistance to snapping and uprooting for Central Amazon tree species. For. Ecol. Manag. 2016, 380, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Paz, H.; Vega-Ramos, F.; Arreola-Villa, F. Understanding hurricane resistance and resilience in tropical dry forest trees: A functional traits approach. For. Ecol. Manag. 2018, 426, 115–122. [Google Scholar] [CrossRef]
- Duryea, M.L.; Kampf, E.; Littell, R.C.; Rodriguez-Pedraza, C.D. Hurricanes and the urban forest: Effects on tropical and subtropical tree species. Arboric Urban For. 2007, 33, 98–112. [Google Scholar]
- Canham, C.D.; Papaik, M.J.; Latty, E.F. Interspecific variation in susceptibility to windthrow as a function of tree size and storm severity for northern temperate tree species. Can. J. For. Res. 2001, 31, 1–10. [Google Scholar] [CrossRef]
- Siliprandi, N.C.; Nogueira, E.M.; Toledo, J.J.; Fearnside, P.M.; Nascimento, H.E.M. Inter-site variation in allometry and wood density of Goupia glabra Aubl. in Amazonia. Braz. J. Biol. 2016, 76, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Niklas, K.J. Plant Biomechanics: An Engineering Approach to Plant Form and Function; University of Chicago: Illinois, IL, USA, 1992. [Google Scholar]
- Zimmerman, J.K.; Everham, E.M.; Waide, R.B.; Lodge, D.J.; Taylor, C.M.; Brokaw, N.V.L. Responses of Tree Species to Hurricane Winds in Subtropical Wet Forest in Puerto-Rico—Implications for Tropical Tree Life-Histories. J. Ecol. 1994, 82, 911–922. [Google Scholar] [CrossRef]
- Xi, W.M. Synergistic effects of tropical cyclones on forest ecosystems: A global synthesis. J. For. Res. 2015, 26, 1–21. [Google Scholar] [CrossRef]
- Shibuya, M.; Ishibashi, S. Stand-level windthrow patterns and long-term dynamics of surviving trees in natural secondary stands after a stand-replacing windthrow event. Forestry 2019, 92, 473–480. [Google Scholar] [CrossRef]
- Lomascolo, T.; Aide, T.M. Seed and seedling bank dynamics in secondary forests following hurricane Georges in Puerto Rico. Caribb. J. Sci. 2001, 37, 259–270. [Google Scholar]
- Pascarella, J.B.; Aide, T.M.; Zimmerman, J.K. Short-term response of secondary forests to hurricane disturbance in Puerto Rico, USA. For. Ecol. Manag. 2004, 199, 379–393. [Google Scholar] [CrossRef]
- Fontes, C.G.; Chambers, J.Q.; Higuchi, N. Revealing the causes and temporal distribution of tree mortality in Central Amazonia. For. Ecol. Manag. 2018, 424, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Metcalfe, D.J.; Bradford, M.G.; Ford, A.J. Cyclone damage to tropical rain forests: Species- and community-level impacts. Austral Ecol. 2008, 33, 432–441. [Google Scholar] [CrossRef]
- Sherman, R.E.; Fahey, T.J.; Martinez, P. Hurricane impacts on a mangrove forest in the Dominican Republic: Damage patterns and early recovery. Biotropica 2001, 33, 393–408. [Google Scholar] [CrossRef]
- Santos, S.L.D.; Válio, I.F. Litter accumulation and its effect on seedling recruitment in a Southeast Brazilian Tropical Forest. Braz. J. Bot. 2002, 25, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Abbas, S.; Nichol, J.E.; Fischer, G.A.; Wong, M.S.; Irteza, S.M. Impact assessment of a super-typhoon on Hong Kong’s secondary vegetation and recommendations for restoration of resilience in the forest succession. Agr. For. Meteorol. 2020, 280, 107784. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, D.; Li, Z.Q.; Wang, L.G. The Effect of Typhoons on the Diversity and Distribution Pattern of Aquatic Plants on Hainan Island, South China. Biotropica 2008, 40, 692–699. [Google Scholar] [CrossRef]
- Fath, B. Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Boose, E.R.; Serrano, M.I.; Foster, D.R. Landscape and regional impacts of hurricanes in Puerto Rico. Ecol. Monogr. 2004, 74, 335–352. [Google Scholar] [CrossRef]
- Verbylaite, R.; Pliura, A.; Lygis, V.; Suchockas, V.; Jankauskiene, J.; Labokas, J. Genetic Diversity and Its Spatial Distribution in Self-Regenerating Norway Spruce and Scots Pine Stands. Forests 2017, 8, 470. [Google Scholar] [CrossRef] [Green Version]
- Murphy, H.T.; Metcalfe, D.J.; Bradford, M.G.; Ford, A.F.; Galway, K.E.; Sydes, T.A.; Westcott, D.J. Recruitment dynamics of invasive species in rainforest habitats following Cyclone Larry. Austral Ecol. 2008, 33, 495–502. [Google Scholar] [CrossRef]
- Alfaro, R.I.; Fady, B.; Vendramin, G.G.; Dawson, I.K.; Fleming, R.A.; Saenz-Romero, C.; Lindig-Cisneros, R.A.; Murdock, T.; Vinceti, B.; Navarro, C.M.; et al. The role of forest genetic resources in responding to biotic and abiotic factors in the context of anthropogenic climate change. For. Ecol. Manag. 2014, 333, 76–87. [Google Scholar] [CrossRef]
- Leimu, R.; Mutikainen, P.; Koricheva, J.; Fischer, M. How general are positive relationships between plant population size, fitness and genetic variation? J. Ecol. 2006, 94, 942–952. [Google Scholar] [CrossRef]
- Monoy, C.C.; Tomlinson, K.W.; Iida, Y.; Swenson, N.G.; Slik, J.W.F. Temporal Changes in Tree Species and Trait Composition in a Cyclone-prone Pacific Dipterocarp Forest. Ecosystems 2016, 19, 1013–1022. [Google Scholar] [CrossRef]
- Swenson, N.G.; Stegen, J.C.; Davies, S.J.; Erickson, D.L.; Forero-Montana, J.; Hurlbert, A.H.; Kress, W.J.; Thompson, J.; Uriarte, M.; Wright, S.J.; et al. Temporal turnover in the composition of tropical tree communities: Functional determinism and phylogenetic stochasticity. Ecology 2012, 93, 490–499. [Google Scholar] [CrossRef]
- Marra, D.M.; Chambers, J.Q.; Higuchi, N.; Trumbore, S.E.; Ribeiro, G.H.P.M.; dos Santos, J.; Negron-Juarez, R.I.; Reu, B.; Wirth, C. Large-Scale Wind Disturbances Promote Tree Diversity in a Central Amazon Forest. PLoS ONE 2014, 9, 103711. [Google Scholar] [CrossRef]
- Miura, M.; Manabe, T.; Nishimura, N.; Yamamoto, S.I. Forest canopy and community dynamics in a temperate old-growth evergreen broad-leaved forest, south-western Japan: A 7-year study of a 4-ha plot. J. Ecol. 2001, 89, 841–849. [Google Scholar] [CrossRef] [Green Version]
- Palmer, M.W.; McAlister, S.D.; Arevalo, J.R.; DeCoster, J.K. Changes in the understory during 14 years following catastrophic windthrow in two Minnesota forests. J. Veg. Sci. 2000, 11, 841–854. [Google Scholar] [CrossRef] [Green Version]
- Birkinshaw, C.; Randrianjanahary, M. The Effects of Cyclone Hudah on the Forest of Masoala Peninsula, Madagascar. Madag. Conserv. Dev. 2007, 2. [Google Scholar] [CrossRef]
- Yao, A.-W.; Chiang, J.-M.; Mcewan, R.; Lin, T.-C. The effect of typhoon-related defoliation on the ecology of gap dynamics in a subtropical rain forest of Taiwan. J. Veg. Sci. 2014, 26, 145–154. [Google Scholar] [CrossRef]
- Kern, C.C.; Montgomery, R.A.; Reich, P.B.; Strong, T.F. Canopy gap size influences niche partitioning of the ground-layer plant community in a northern temperate forest. J. Plant Ecol. 2012, 6, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.C.; Hamburg, S.P.; Lin, K.C.; Wang, L.J.; Chang, C.T.; Hsia, Y.J.; Vadeboncoeur, M.A.; McMullen, C.M.M.; Liu, C.P. Typhoon Disturbance and Forest Dynamics: Lessons from a Northwest Pacific Subtropical Forest. Ecosystems 2011, 14, 127–143. [Google Scholar] [CrossRef]
- Namikawa, K.; Matsui, T.; Kobayashi, M.; Goto, R.; Kuramoto, S. Initial establishment and regeneration processes of an outlying isolated Fagus crenata Blume forest stand in the northernmost boundary of its range in Hokkaido, northern Japan. Plant Ecol. 2010, 207, 161–174. [Google Scholar] [CrossRef]
- Yang, H.; Liu, S.R.; Cao, K.F.; Wang, J.X.; Li, Y.D.; Xu, H. Characteristics of typhoon disturbed gaps in an old-growth tropical montane rainforest in Hainan Island, China. J. For. Res. 2017, 28, 1231–1239. [Google Scholar] [CrossRef]
- Ruiz, J.; Fandino, M.C. The Impact of Hurricane Beta on the Forests of Providencia Island, Colombia, Southwest Caribbean. Caldasia 2010, 32, 425–434. [Google Scholar]
- Imbert, D.; Portecop, J. Hurricane disturbance and forest resilience: Assessing structural vs. functional changes in a Caribbean dry forest. For. Ecol. Manag. 2008, 255, 3494–3501. [Google Scholar] [CrossRef]
- Shang, K.-K.; Zhang, Q.-P.; Da, L.-J.; Hara, K.; Yang, Y.-C.; Fujihara, M.; Tomita, M.; Zhao, Y. Effects of natural and artificial disturbance on landscape and forest structure in Tiantong National Forest Park, East China. Landsc. Ecol. Eng. 2011, 10, 163–172. [Google Scholar] [CrossRef]
- Chapman, E.L.; Chambers, J.Q.; Ribbeck, K.F.; Baker, D.B.; Tobler, M.A.; Zeng, H.C.; White, D.A. Hurricane Katrina impacts on forest trees of Louisiana’s Pearl River basin. For. Ecol. Manag. 2008, 256, 883–889. [Google Scholar] [CrossRef]
- Petrokas, R.; Baliuckas, V.; Manton, M. Successional Categorization of European Hemi-boreal Forest Tree Species. Plants 2020, 9, 1381. [Google Scholar] [CrossRef] [PubMed]
- Bellingham, P.J.; Tanner, E.V.J.; Healey, J.R. Hurricane disturbance accelerates invasion by the alien tree Pittosporum undulatum in Jamaican montane rain forests. J. Veg. Sci. 2005, 16, 675–684. [Google Scholar] [CrossRef]
- Salinas, N.; Malhi, Y.; Meir, P.; Silman, M.; Cuesta, R.R.; Huaman, J.; Salinas, D.; Farfan, F. The sensitivity of tropical leaf litter decomposition to temperature: Results from a large-scale leaf translocation experiment along an elevation gradient in Peruvian forests. New Phytol. 2010, 189, 967–977. [Google Scholar] [CrossRef]
- Park, B.B.; Rahman, A.; Han, S.H.; Youn, W.B.; Hyun, H.J.; Hernandez, J.; An, J.Y. Carbon and Nutrient Inputs by Litterfall in Evergreen and Deciduous Forests in Korea. Forests 2020, 11, 143. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Yrizar, A.; Jaramillo, V.J.; Maass, M.; Burquez, A.; Parker, G.; Alvarez-Yepiz, J.C.; Araiza, S.; Verduzco, A.; Sarukhan, J. Resilience of tropical dry forest productivity to two hurricanes of different intensity in western Mexico. For. Ecol. Manag. 2018, 426, 53–60. [Google Scholar] [CrossRef]
- Bloem, S.J.V.; Murphy, P.G.; Lugo, A.E.; Ostertag, R.; Costa, M.R.; Bernard, I.R.; Colon, S.M.; Mora, M.C. The Influence of Hurricane Winds on Caribbean Dry Forest Structure and Nutrient Pools1. Biotropica 2005, 37, 571–583. [Google Scholar] [CrossRef]
- Cordova, S.C.; Olk, D.C.; Dietzel, R.N.; Mueller, K.E.; Archontouilis, S.V.; Castellano, M.J. Plant litter quality affects the accumulation rate, composition, and stability of mineral-associated soil organic matter. Soil Biol. Biochem. 2018, 125, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Beard, K.H.; Vogt, K.A.; Vogt, D.J.; Scatena, F.N.; Covich, A.P.; Sigurdardottir, R.; Siccama, T.G.; Crowl, T.A. Structural and functional responses of a subtropical forest to 10 years of hurricanes and droughts. Ecol. Monogr. 2005, 75, 345–361. [Google Scholar] [CrossRef] [Green Version]
- Bonanomi, G.; Cesarano, G.; Lombardi, N.; Motti, R.; Scala, F.; Mazzoleni, S.; Incerti, G. Litter chemistry explains contrasting feeding preferences of bacteria, fungi, and higher plants. Sci. Rep. 2017, 7, 9208. [Google Scholar] [CrossRef] [Green Version]
- Eaton, W.D.; McGee, K.M.; Alderfer, K.; Jimenez, A.R.; Hajibabaei, M. Increase in abundance and decrease in richness of soil microbes following Hurricane Otto in three primary forest types in the Northern Zone of Costa Rica. PLoS ONE 2020, 15, e0231187. [Google Scholar] [CrossRef] [PubMed]
- Vanderwel, M.C.; Malcolm, J.R.; Smith, S.M. An integrated model for snag and downed woody debris decay class transitions. For. Ecol. Manag. 2006, 234, 48–59. [Google Scholar] [CrossRef]
- McNulty, S.G. Hurricane impacts on US forest carbon sequestration. Environ. Pollut. 2002, 116, S17–S24. [Google Scholar] [CrossRef]
- Laurance, W.F.; Curran, T.J. Impacts of wind disturbance on fragmented tropical forests: A review and synthesis. Austral Ecol. 2008, 33, 399–408. [Google Scholar] [CrossRef]
- Smith, I.A.; Hutyra, L.R.; Reinmann, A.B.; Marrs, J.K.; Thompson, J.R. Piecing together the fragments: Elucidating edge effects on forest carbon dynamics. Front. Ecol. Environ. 2018, 16, 213–221. [Google Scholar] [CrossRef]
- Fauset, S.; Johnson, M.O.; Gloor, M.; Baker, T.R.; Monteagudo, A.; Brienen, R.J.W.; Feldpausch, T.R.; Lopez-Gonzalez, G.; Malhi, Y.; ter Steege, H.; et al. Hyperdominance in Amazonian forest carbon cycling. Nat. Commun. 2015, 6, 6857. [Google Scholar] [CrossRef]
- Laurance, W.F.; Nascimento, H.E.; Laurance, S.G.; Andrade, A.C.; Fearnside, P.M.; Ribeiro, J.E.; Capretz, R.L. Rain forest fragmentation and the proliferation of successional trees. Ecology 2006, 87, 469–482. [Google Scholar] [CrossRef]
- Melito, M.; Metzger, J.P.; Oliveira, A.A. Landscape-level effects on aboveground biomass of tropical forests: A conceptual framework. Glob. Change Biol. 2017, 24, 597–607. [Google Scholar] [CrossRef]
- Michalski, F.; Nishi, I.; Peres, C.A. Disturbance-mediated drift in tree functional groups in Amazonian forest fragments. Biotropica 2007, 39, 691–701. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Fisher, J.I.; Zeng, H.C.; Chapman, E.L.; Baker, D.B.; Hurtt, G.C. Hurricane Katrina’s carbon footprint on U. S. Gulf Coast forests. Science 2007, 318, 1107. [Google Scholar] [CrossRef] [Green Version]
- Briber, B.M.; Hutyra, L.R.; Reinmann, A.B.; Raciti, S.M.; Dearborn, V.K.; Holden, C.E.; Dunn, A.L. Tree Productivity Enhanced with Conversion from Forest to Urban Land Covers. PLoS ONE 2015, 10, e0136237. [Google Scholar] [CrossRef] [PubMed]
- Reinmann, A.B.; Hutyra, L.R. Edge effects enhance carbon uptake and its vulnerability to climate change in temperate broadleaf forests. Proc. Natl. Acad. Sci. USA 2017, 114, 107–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remy, E.; Wuyts, K.; Boeckx, P.; Gundersen, P.; Verheyen, K. Edge effects in temperate forests subjected to high nitrogen deposition. Proc. Natl. Acad. Sci. USA 2017, 114, E7032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.Q.; Franklin, J.F.; Spies, T.A. Vegetation Responses to Edge Environments in Old-Growth Douglas-Fir Forests. Ecol. Appl. 1992, 2, 387–396. [Google Scholar] [CrossRef]
- Moran, C.; Catterall, C.P.; Green, R.J.; Olsen, M.F. Functional variation among frugivorous birds: Implications for rainforest seed dispersal in a fragmented subtropical landscape. Oecologia 2004, 141, 584–595. [Google Scholar] [CrossRef]
- Lee, M.F.; Lin, T.C.; Vadeboncoeur, M.A.; Hwong, J.L. Remote sensing assessment of forest damage in relation to the 1996 strong typhoon Herb at Lienhuachi Experimental Forest, Taiwan. For. Ecol. Manag. 2008, 255, 3297–3306. [Google Scholar] [CrossRef]
- Hu, T.G.; Smith, R.B. The Impact of Hurricane Maria on the Vegetation of Dominica and Puerto Rico Using Multispectral Remote Sensing. Remote Sens. 2018, 10, 827. [Google Scholar] [CrossRef] [Green Version]
- Battles, J.J.; Cleavitt, N.L.; Saah, D.S.; Poling, B.T.; Fahey, T.J. Ecological impact of a microburst windstorm in a northern hardwood forest. Can. J. For. Res. 2017, 47, 1695–1701. [Google Scholar] [CrossRef]
- Bhowmik, A.K.; Cabral, P. Cyclone Sidr Impacts on the Sundarbans Floristic Diversity. Earth Sci. Res. 2013, 2, 62. [Google Scholar] [CrossRef] [Green Version]
- Bianchette, T.A.; Liu, K.B.; Lam, N.S.N.; Kiage, L.M. Ecological Impacts of Hurricane Ivan on the Gulf Coast of Alabama: A Remote Sensing Study. J. Coast. Res. 2009, 2, 1622–1626. [Google Scholar]
- Prabhakara, K.; Hively, W.D.; McCarty, G.W. Evaluating the relationship between biomass, percent groundcover and remote sensing indices across six winter cover crop fields in Maryland, United States. Int. J. Appl. Earth Obs. 2015, 39, 88–102. [Google Scholar] [CrossRef] [Green Version]
- Rahetlah, V.B.; Andrianarisoa, B.; Salgado, P.; Tillard, T.; Razafindrazaka, H.; Le Mézo, L.; Ramalanjaona, V.L. Relationship between normalized difference vegetation index (NDVI) and forage biomass yield in the Vakinankaratra region, Madagascar. Lives Res. Rural Dev. 2014, 26, 11. [Google Scholar]
- Lewis, R.J.; Bannar-Martin, K.H. The Impact of Cyclone Fanele on a Tropical Dry Forest in Madagascar. Biotropica 2011, 44, 135–140. [Google Scholar] [CrossRef]
- Lee, T.-C.; Knutson, T.R.; Nakaegawa, T.; Ying, M.; Cha, E.J. Third assessment on impacts of climate change on tropical cyclones in the Typhoon Committee Region—Part I: Observed changes, detection and attribution. Trop. Cyclone Res. Rev. 2020, 9, 1–22. [Google Scholar] [CrossRef]
- Shimada, U.; Yamaguchi, M.; Nishimura, S. Is the number of tropical cyclone rapid intensification events in the western north pacific increasing? SOLA 2020, 16, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Cha, E.J.; Knutson, T.R.; Lee, T.; Ying, M.; Nakaegawa, T. Third assessment on impacts of climate change on tropical cyclones in the Typhoon Committee Region—Part II: Future projections. Trop. Cyclone Res. Rev. 2020, 9, 75–86. [Google Scholar] [CrossRef]
- Kang, N.Y.; Elsner, J.B. Influence of global warming on the rapid intensification of western North Pacific tropical cyclones. Environ. Res. Lett. 2019, 14, 044027. [Google Scholar] [CrossRef]
- Song, J.J.; Duan, Y.H.; Klotzbach, P.J. Increasing trend in rapid intensification magnitude of tropical cyclones over the western North Pacific. Environ. Res. Lett. 2020, 15, 084043. [Google Scholar] [CrossRef]
- Choy, C.W.; Chong, S.N.; Kong, D.X.; Cayanan, E.O. A Discussion of the Most Intense Tropical Cyclones in the Western North Pacific from 1978 to 2013. Trop. Cyclone Res. Rev. 2015, 4, 1–11. [Google Scholar] [CrossRef]
- Takagi, H.; Esteban, M. Statistics of tropical cyclone landfalls in the Philippines: Unusual characteristics of 2013 Typhoon Haiyan. Nat. Hazards 2016, 80, 211–222. [Google Scholar] [CrossRef]
- Lin, I.I.; Pun, I.F.; Lien, C.C. “Category-6” supertyphoon Haiyan in global warming hiatus: Contribution from subsurface ocean warming. Geophys. Res. Lett. 2014, 41, 8547–8553. [Google Scholar] [CrossRef]
- Takagi, H.; Esteban, M.; Shibayama, T.; Mikami, T.; Matsumaru, R.; Leon, M.D.; Thao, N.D.; Oyama, T.; Nakamura, R. Track analysis, simulation, and field survey of the 2013 Typhoon Haiyan storm surge. J. Flood Risk Manag. 2014, 10, 42–52. [Google Scholar] [CrossRef]
- Parolin, P.; Wittmann, F. Struggle in the flood: Tree responses to flooding stress in four tropical floodplain systems. Aob Plants 2010, 2010, plq003. [Google Scholar] [CrossRef] [Green Version]
- Shultz, D. How Tropical Cyclones Influence Photosynthesis. Eos 2016, 97. [Google Scholar] [CrossRef]
- Poorter, L.; Kitajima, K.; Mercado, P.; Chubina, J.; Melgar, I.; Prins, H.H.T. Resprouting as a persistence strategy of tropical forest trees: Relations with carbohydrate storage and shade tolerance. Ecology 2010, 91, 2613–2627. [Google Scholar] [CrossRef]
- Myers, J.A.; Kitajima, K. Carbohydrate storage enhances seedling shade and stress tolerance in a neotropical forest. J. Ecol. 2007, 95, 383–395. [Google Scholar] [CrossRef]
- Weemstra, M.; Kiorapostolou, N.; van Ruijven, J.; Mommer, L.; de Vries, J.; Sterck, F. The role of fine-root mass, specific root length and life span in tree performance: A whole-tree exploration. Funct. Ecol. 2020, 34, 575–585. [Google Scholar] [CrossRef]
- López-Marrero, T.; Heartsill-Scalley, T.; Rivera-López, C.F.; Escalera-García, I.A.; Echevarría-Ramos, M. Broadening Our Understanding of Hurricanes and Forests on the Caribbean Island of Puerto Rico: Where and What Should We Study Now? Forests 2019, 10, 710. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, E.; Castro, J.; Araujo, J.; Heineck, T. A Systematic Literature Review of iStar extensions. J. Syst. Softw. 2018, 137, 1–33. [Google Scholar] [CrossRef]
- Perevochtchikova, M.; De la Mora-De la Mora, G.; Flores, J.A.H.; Marin, W.; Flores, A.L.; Bueno, A.R.; Negrete, I.A.R. Systematic review of integrated studies on functional and thematic ecosystem services in Latin America, 1992–2017. Ecosyst. Serv. 2019, 36, 100900. [Google Scholar] [CrossRef]
- Shea, B.J.; Reeves, B.C.; Wells, G.; Thuku, M.; Hamel, C.; Moran, J.; Moher, D.; Tugwell, P.; Welch, V.; Kristjansson, E.; et al. AMSTAR 2: A critical appraisal tool for systematic reviews that include randomised or non-randomised studies of healthcare interventions, or both. Bmj-Brit. Med. J. 2017, 358, j4008. [Google Scholar] [CrossRef] [Green Version]
- Acharya, R.P.; Maraseni, T.; Cockfield, G. Global trend of forest ecosystem services valuation—An analysis of publications. Ecosyst. Serv. 2019, 39, 100979. [Google Scholar] [CrossRef]
- Hoque, M.A.; Phinn, S.; Roelfsema, C. A systematic review of tropical cyclone disaster management research using remote sensing and spatial analysis. Ocean Coast Manag. 2017, 146, 109–120. [Google Scholar] [CrossRef]
- Poudyal, B.H.; Maraseni, T.; Cockfield, G. Evolutionary dynamics of selective logging in the tropics: A systematic review of impact studies and their effectiveness in sustainable forest management. For. Ecol. Manag. 2018, 430, 166–175. [Google Scholar] [CrossRef]
- Diamond, J. Overview: Laboratory experiments, field experiments, and natural experiments. In Community Ecology; Diamond, J., Case, T.J., Eds.; Harper and Row: New York, NY, USA, 1986; pp. 1–22. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Search Terms and Strings | No. of Articles | ||
|---|---|---|---|
| Science Direct | Scopus | Total | |
| 1. Windstorms OR “typhoon” OR “tropical cyclone” OR “hurricane” AND “tropical forest” OR “subtropical forest” OR “temperate forest” | 5026 | 283 | 5309 |
| 2. Windstorms OR “typhoon” OR “tropical cyclone” OR “hurricane” AND “forest structure” OR “tree population” OR “tree genetic diversity” OR “tree diversity” OR “tree mortality” | 2757 | 231 | 2988 |
| 3. Windstorms OR “typhoon” OR “tropical cyclone” OR “hurricane” AND “tree biomass” OR “forest carbon sequestration” OR “forest nutrient cycling” OR “tree regeneration” OR “tree species composition” | 2636 | 49 | 2685 |
| 4. Windstorms OR “typhoon” OR “tropical cyclone” OR “hurricane” AND “gap dynamics” OR “tree demography” “tree primary productivity” OR “tree seedling growth” OR “seedling germination” OR “forest ecosystem structure” | 34 | 1 | 35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez, J.O.; Maldia, L.S.J.; Park, B.B. Research Trends and Methodological Approaches of the Impacts of Windstorms on Forests in Tropical, Subtropical, and Temperate Zones: Where Are We Now and How Should Research Move Forward? Plants 2020, 9, 1709. https://doi.org/10.3390/plants9121709
Hernandez JO, Maldia LSJ, Park BB. Research Trends and Methodological Approaches of the Impacts of Windstorms on Forests in Tropical, Subtropical, and Temperate Zones: Where Are We Now and How Should Research Move Forward? Plants. 2020; 9(12):1709. https://doi.org/10.3390/plants9121709
Chicago/Turabian StyleHernandez, Jonathan O., Lerma S.J. Maldia, and Byung Bae Park. 2020. "Research Trends and Methodological Approaches of the Impacts of Windstorms on Forests in Tropical, Subtropical, and Temperate Zones: Where Are We Now and How Should Research Move Forward?" Plants 9, no. 12: 1709. https://doi.org/10.3390/plants9121709
APA StyleHernandez, J. O., Maldia, L. S. J., & Park, B. B. (2020). Research Trends and Methodological Approaches of the Impacts of Windstorms on Forests in Tropical, Subtropical, and Temperate Zones: Where Are We Now and How Should Research Move Forward? Plants, 9(12), 1709. https://doi.org/10.3390/plants9121709