Germination and Growth of Spinach under Potassium Deficiency and Irrigation with High-Salinity Water

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenhouse Cultivation Details
2.2. Plant Mineral Analyses
2.3. Saline Solutions
2.4. Statistical Analyses
3. Results
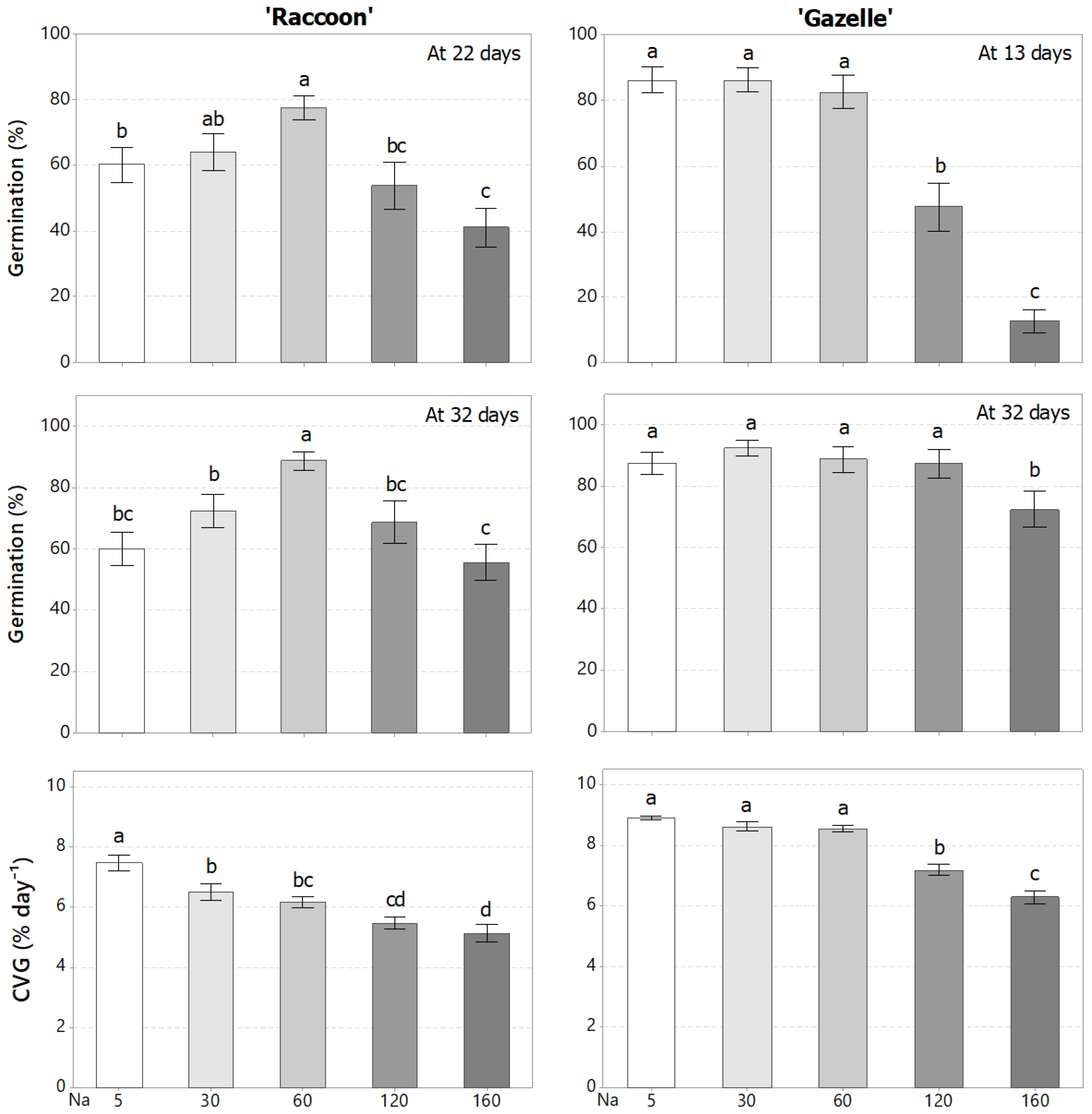
3.1. Effect of Combined Salinity and Potassium Doses on Germination and Seedling Establishment
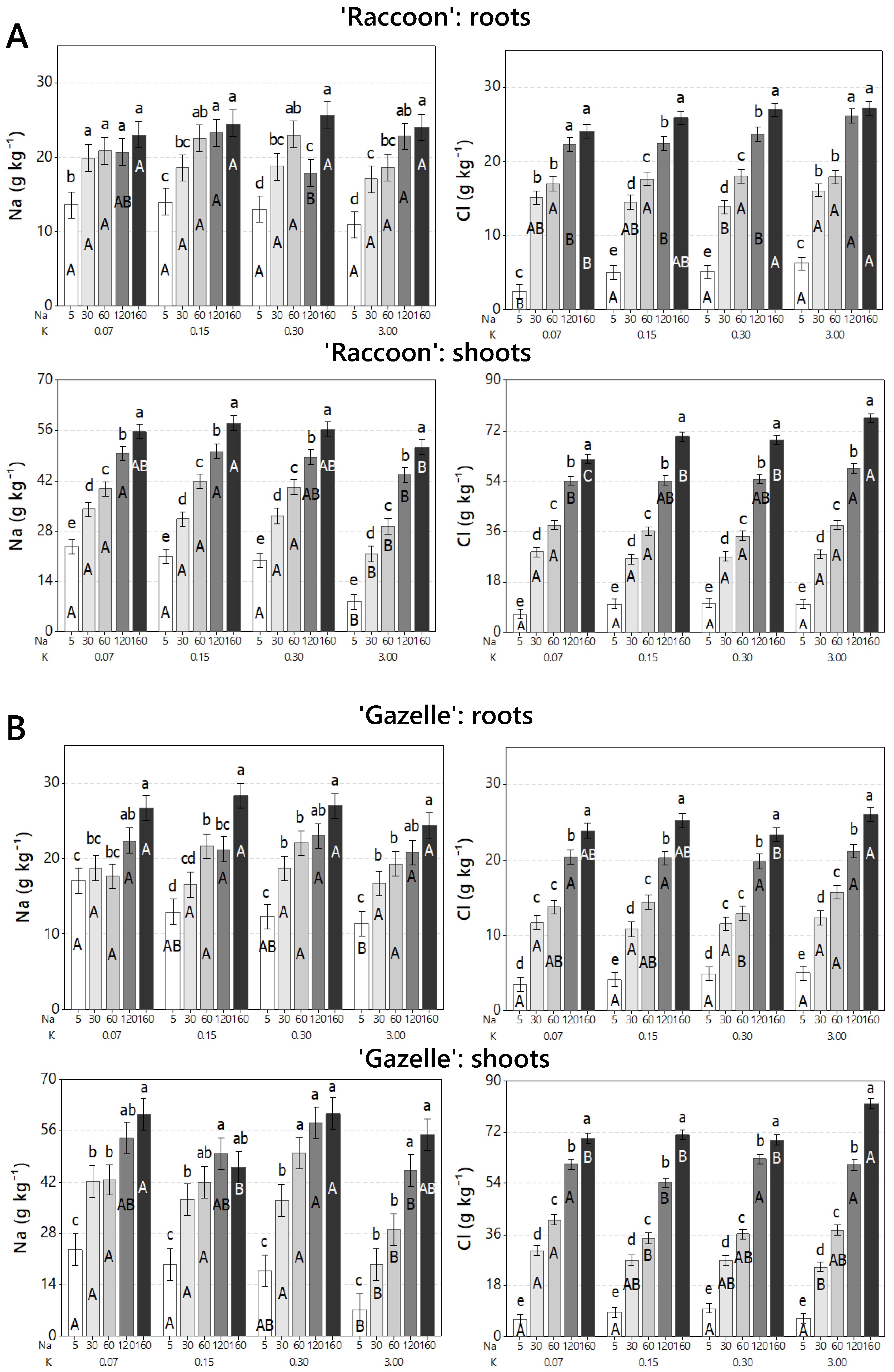
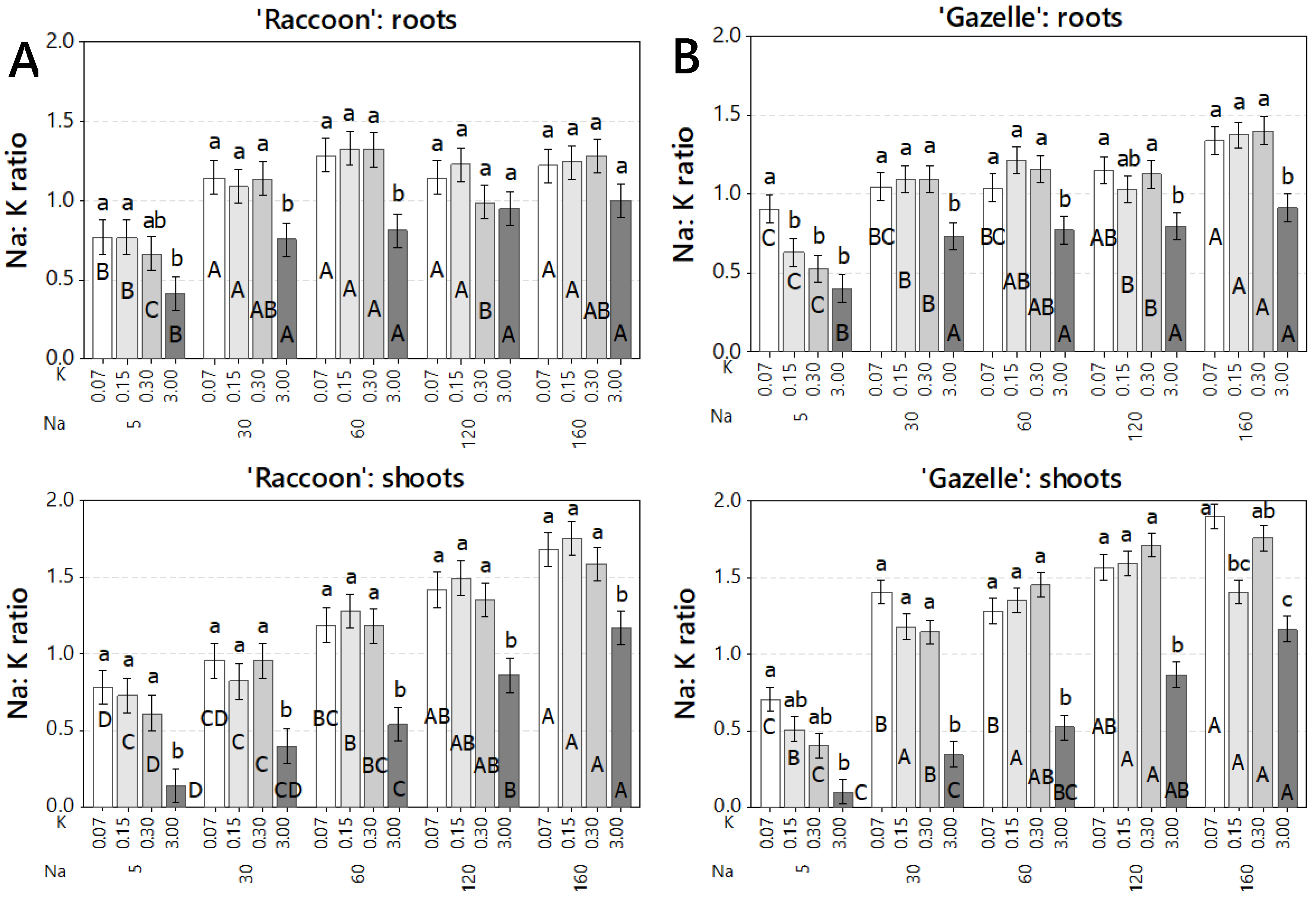
3.2. Effect of Combined Salinity and Potassium Doses on Tissue Na and Cl Accumulation
3.3. Effect of Combined Salinity and Potassium Doses on Root Mineral Composition
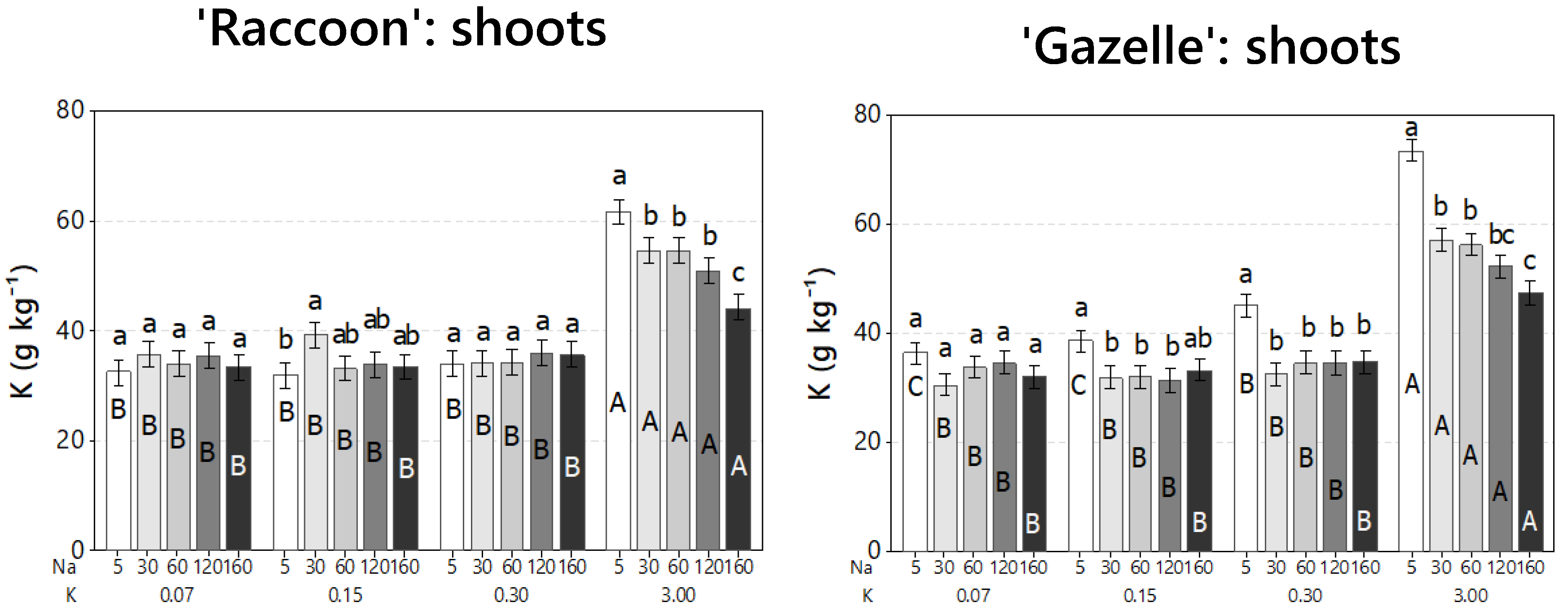
3.4. Effect of Combined Salinity and Potassium Doses on Shoot Mineral Composition
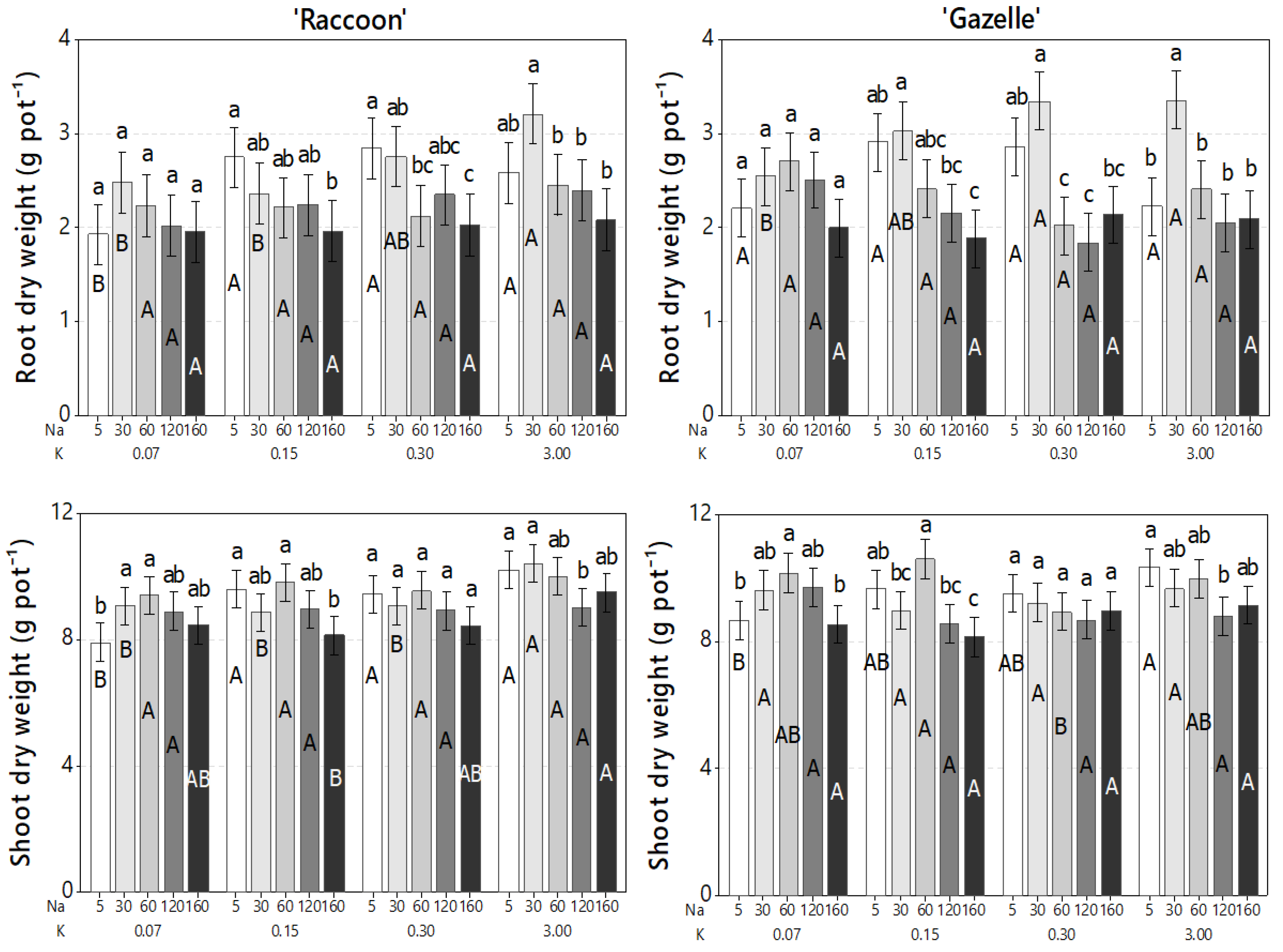
3.5. Effect of Combined Salinity and Potassium Doses on Plant Biomass Accumulation
4. Discussion
4.1. Effect of Combined Salinity and Potassium Doses on Germination and Seedling Establishment
4.2. Effect of Combined Salinity and Potassium Doses on Tissue Na and Cl Accumulation
4.3. Effect of Combined Salinity and Potassium Doses on Root Mineral Composition
4.4. Effect of Combined Salinity and Potassium Doses on Shoot Mineral Composition
4.5. Effect of Combined Salinity and Potassium Doses on Plant Biomass Accumulation
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ashraf, M. Breeding for salinity tolerance in plants. Crit. Rev. Plant Sci. 1994, 13, 17–42. [Google Scholar] [CrossRef]
- Levitt, J. Responses of Plants to Environmental Stresses, Water, Radiation, Salt and Other Stresses; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Dias, N.S.; Ferreira, J.F.S.; Liu, X.; Suarez, D.L. Jerusalem artichoke (Helianthus tuberosus L.) maintains high inulin, tuber yield, and antioxidant capacity under moderately-saline irrigation waters. Ind. Crops Prod. 2016, 94, 1009–1024. [Google Scholar] [CrossRef]
- Ferreira, J.F.S.; Sandhu, D.; Liu, X.; Halvorson, J.J. Spinach (Spinacea oleracea L.) response to salinity: Nutritional value, physiological parameters, antioxidant capacity, and gene expression. Agriculture 2018, 8, 163. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.F.S.; Cornacchione, M.; Liu, X.; Suarez, D. Nutrient Composition, Forage Parameters, and Antioxidant Capacity of Alfalfa (Medicago sativa, L.) in Response to Saline Irrigation Water. Agriculture 2015, 5, 577–597. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.F.S.; da Silva, J.B.; Liu, X.; Sandhu, D. Spinach plants favor the absorption of K+ over Na+ regardless of salinity, and may benefit from Na+ when K+ is deficient in the soil. Plants 2020, 9, 507. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.F.S.; Liu, X.; Suarez, D.L. Fruit yield and survival of five commercial strawberry cultivars under field cultivation and salinity stress. Sci. Hortic. 2019, 243, 401–410. [Google Scholar] [CrossRef]
- Suarez, D.L.; Grieve, C.M. Growth, Yield, and Ion Relations of Strawberry in Response To Irrigation With Chloride-Dominated Waters. J. Plant Nutr. 2013, 36, 1963–1981. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Jagendorf, A.; Zhu, J.K. Understanding and improving salt tolerance in plants. Crop Sci. 2005, 45, 437–448. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Fan, H.; Zhao, Y.; Jia, Y.; Du, X.; Wang, B. Effect of salinity on germination, seedling emergence, seedling growth and ion accumulation of a euhalophyte Suaeda salsa in an intertidal zone and on saline inland. Aquat. Bot. 2008, 88, 331–337. [Google Scholar] [CrossRef]
- Zhou, D.; Xiao, M. Specific ion effects on the seed germination of sunflower. J. Plant Nutr. 2010, 33, 255–266. [Google Scholar] [CrossRef]
- Gomes-Filho, E.; Lima, C.R.F.M.; Costa, J.H.; Da Silva, A.C.M.; Da Guia Silva Lima, M.; De Lacerda, C.F.; Prisco, J.T. Cowpea ribonuclease: Properties and effect of NaCl-salinity on its activation during seed germination and seedling establishment. Plant Cell Rep. 2008, 27, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Yupsanis, T.; Moustakas, M.; Eleftheriou, P.; Damianidou, K. Protein phosphorylation-dephosphorylation in alfalfa seeds germinating under salt stress. J. Plant Physiol. 1994, 143, 234–240. [Google Scholar] [CrossRef]
- Dantas, B.F.; De Sá Ribeiro, L.; Aragão, C.A. Germination, initial growth and cotyledon protein content of bean cultivars under salinity stress. Rev. Bras. Sementes 2007, 29, 106–110. [Google Scholar] [CrossRef]
- Khan, M.A.; Rizvi, Y. Effect of salinity, temperature, and growth regulators on the germination and early seedling growth of Atriplex griffithii var. stocksii. Can. J. Bot. 1994, 72, 475–479. [Google Scholar] [CrossRef]
- Promila, K.; Kumar, S. Vigna radiata seed germination under salinity. Biol. Plant. 2000, 43, 423–426. [Google Scholar] [CrossRef]
- Tawaha, A.M.; Othman, Y.; Al-Karaki, G.; Al-Tawaha, A.R.; Al-Horani, A. Variation in germination and ion uptake in barley genotypes under salinity conditions. World J. Agric. Sci. 2006, 2, 11–15. [Google Scholar]
- Turhan, A.; Kuşcu, H.; Şeniz, V. Effects of different salt concentrations (NaCl) on germination of some spinach cultivars. Uludağ Üniv. Ziraat Fak. Derg. 2011, 25, 65–77. [Google Scholar] [CrossRef]
- Borsai, O.; Al-Hassan, M.; Boscaiu, M.; Sestras, R.E.; Vicente, O. Effects of salt and drought stress on seed germination and seedling growth in Portulaca. Rom. Biotechnol. Lett. 2018, 23, 13340–13349. [Google Scholar]
- Sandhu, D.; Cornacchione, M.V.; Ferreira, J.F.S.; Suarez, D.L. Variable salinity responses of 12 alfalfa genotypes and comparative expression analyses of salt-response genes. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, D.; Pudussery, M.V.; Ferreira, J.F.S.; Liu, X.; Pallete, A.; Grover, K.K.; Hummer, K. Variable salinity responses and comparative gene expression in woodland strawberry genotypes. Sci. Hortic. 2019, 254, 61–69. [Google Scholar] [CrossRef]
- Zrig, A.; Ferreira, J.F.S.; Serrano, M.; Valero, D.; Tounekti, T.; Khemira, H. Polyamines and other secondary metabolites of green-leaf and red-leaf almond rootstocks trigger in response to salinity. Pak. J. Bot. 2018, 50, 1273–1279. [Google Scholar]
- Abdul Jaleel, C.; Gopi, R.; Sankar, B.; Manivannan, P.; Kishorekumar, A.; Sridharan, R.; Panneerselvam, R. Studies on germination, seedling vigour, lipid peroxidation and proline metabolism in Catharanthus roseus seedlings under salt stress. S. Afr. J. Bot. 2007, 73, 190–195. [Google Scholar] [CrossRef] [Green Version]
- Kaymakanova, M. Effect of salinity on germination and seed physiology in bean (Phaseolus vulgaris L.). Biotechnol. Biotechnol. Equip. 2009, 23, 326–329. [Google Scholar] [CrossRef] [Green Version]
- Prado, F.E.; Boero, C.; Gallardo, M.; González, J.A. Effect of NaCl on germination, growth, and soluble sugar content in Chenopodium quinoa Willd. seeds. Bot. Bull. Acad. Sin. 2000, 41, 27–34. [Google Scholar] [CrossRef]
- Brakez, M.; Harrouni, M.C.; Tachbibi, N.; Daoud, S. Comparative effect of NaCl and seawater on germination of quinoa seed (Chenopodium quinoa willd). Emir. J. Food Agric. 2014, 26, 1091–1096. [Google Scholar] [CrossRef]
- Al-Harbi, A.R.; Wahb-Allah, M.A.; Abu-Muriefah, S.S. Salinity and nitrogen level affects germination, emergence, and seedling growth of tomato. Int. J. Veg. Sci. 2008, 14, 380–392. [Google Scholar] [CrossRef]
- Jamil, M.; Lee, D.B.A.E.; Jung, K.Y.; Ashraf, M.; Chun, S.; Rha, E.U.I.S. Effect of salt (Nacl) stress on germination and early seedling growth of four vegetables species. J. Cent. Eur. Agric. 2006, 7, 273–282. [Google Scholar] [CrossRef]
- Janagard, M.S.; Tobeh, A.; Esmailpour, B. Evaluation of salinity tolerance of three canola cultivars at germination and early seedling growth stage. J. Food Agric. Environ. 2008, 6, 272–275. [Google Scholar]
- Abdel-Haleem, A.; El-Shaieny, H. Seed germination percentage and early seedling establishment of five (Vigna unguiculata L. (Walp) genotypes under salt stress. Eur. J. Exp. Biol. 2015, 5, 22–32. [Google Scholar]
- Alemzadeh, N.; Khaleghi, A.S. Germination and emergence response of some onion cultivars of Southern Iran to salinity stress. Seed Sci. Biotechnol. 2009, 3, 21–23. [Google Scholar]
- Kaya, M.D.; Ipek, A.; Öztürk, A. Effects of different soil salinity levels on germination and seedling growth of safflower (Carthamus tinctorius L.). Turk. J. Agric. For. 2003, 27, 221–227. [Google Scholar] [CrossRef]
- Wu, G.Q.; Jiao, Q.; Shui, Q.Z. Effect of salinity on seed germination, seedling growth, and inorganic and organic solutes accumulation in sunflower (Helianthus annuus L.). Plant Soil Environ. 2015, 61, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Akinci, I.E.; Akinci, S.; Yilmaz, K.; Dikici, H. Response of eggplant varieties (Solanum melongena) to salinity in germination and seedling stages. N. Z. J. Crop Hortic. Sci. 2004, 32, 193–200. [Google Scholar] [CrossRef]
- Kahouli, B.; Borgi, Z.; Hannachi, C. Effect of sodium chloride on the germination of the seeds of a collection of carrot accessions (Daucus carota L.) cultivated in the region of Sidi Bouzid. J. Stress Physiol. Biochem. 2014, 10, 28–36. [Google Scholar]
- Subbarao, G.V.; Wheeler, R.M.; Stutte, G.W.; Levine, L.H. How-far can sodium substitute for potassium in red beet? J. Plant Nutr. 1999, 22, 1745–1761. [Google Scholar] [CrossRef]
- Nemadodzi, L.E.; Araya, H.; Nkomo, M.; Ngezimana, W.; Mudau, N.F. Nitrogen, phosphorus, and potassium effects on the physiology and biomass yield of baby spinach (Spinacia oleracea L.). J. Plant Nutr. 2017, 40, 2033–2044. [Google Scholar] [CrossRef]
- Ors, S.; Suarez, D.L. Salt tolerance of spinach as related to seasonal climate. Hortic. Sci. 2016, 43, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Koike, S.T.; Cahn, M.; Fennimore, S.; Lestrange, M.; Natwick, E.; Smith, R.F.; Takele, E. Vegetable Production Series in Spinach Production in California; Publication 7212; University of California Agricultural and Natural Resources: Davis, CA, USA, 2011; 6p. [Google Scholar]
- Karege, F.; Penel, C.; Greppin, H. Floral induction in spinach leaves by light, temperature and gibberellic acid: Use of the photocontrol of basic peroxidase activity as biochemical marker. Z. Pflanzenphysiol. 1982, 107, 357–365. [Google Scholar] [CrossRef]
- Kader, M.A. A Comparison of seed germination calculation formulae and the associated interpretation of resulting data. J. Proc. R. Soc. N. S. W. 2005, 138, 65–75. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; California Agricultural Experiment Station: Berkeley, CA, USA, 1950; Volume Circular 347, 32p. [Google Scholar]
- Ribeiro Junior, J.I.; Melo, A.L.P. Guia Prático Para Utilização do Saeg; Folha Artes Graficas Ltd.: Viçosa, Brazil, 2008. [Google Scholar]
- Lacerda, C.F.; Ferreira, J.F.S.; Liu, X.; Suarez, D.L. Evapotranspiration as a criterion to estimate nitrogen requirement of maize under salt stress. J. Agron. Crop Sci. 2016, 202, 192–202. [Google Scholar] [CrossRef]
- Morgan, K.T. (Ed.) Nutrient Management of Vegetable and Row Crops Handbook (SP500); University of Florida: Gainesville, FL, USA, 2015. [Google Scholar]
- Schröppel-Meier, G.; Kaiser, W.M. Ion homeostasis in chloroplasts under salinity and mineral deficiency. Plant Physiol. 1988, 87, 828–832. [Google Scholar] [CrossRef] [PubMed]
- Sheikhi, J.; Ronaghi, A. Growth and macro and micronutrients concentration in spinach (Spinacia oleracea L.) as influenced by salinity and nitrogen rates. Int. Res. J. Appl. Basic Sci. 2012, 3, 770–777. [Google Scholar]
- Chakraborty, K.; Bose, J.; Shabala, L.; Shabala, S. Difference in root K+ retention ability and reduced sensitivity of K+-permeable channels to reactive oxygen species confer differential salt tolerance in three Brassica species. J. Exp. Bot. 2016, 67, 4611–4625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, W.; Ball, M.; Anderson, J. Growth and photosynthetic responses of spinach to salinity: Implications of K+ nutrition for salt tolerance. Funct. Plant Biol. 1990, 17, 563. [Google Scholar] [CrossRef]
- Raddatz, N.; Morales de los Ríos, L.; Lindahl, M.; Quintero, F.J.; Pardo, J.M. Coordinated transport of nitrate, potassium, and sodium. Front. Plant Sci. 2020, 11, 247. [Google Scholar] [CrossRef] [Green Version]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ nutrition in plants. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Grieve, C.; Grattan, S.; Maas, E. Plant salt tolerance. In Agricultural Salinity Assessment and Management; Wallender, W.W., Tanji, K.K., Eds.; ASCE Press: Reston, VA, USA, 2012; pp. 405–459. [Google Scholar]
- Maas, E.V.; Grattan, S.R. Crop Yields as Affected by Salinity. In Agricultural Drainage; Skaggs, R.W., Schilfgaarde, J.V., Eds.; ASA/CSSA/SSSA: Madison, WI, USA, 1999; pp. 55–108. [Google Scholar]
- Maas, E.V.; Hoffman, G.J. Crop salt tolerance-current assessment. J. Irrig. Drain. Div. 1977, 103, 115–134. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | ARH (%) | Temp (°C) | PPFD * (µmol m−2 s−1) | Day Length ** (h:min:s) |
|---|---|---|---|---|
| October 19/2018 (min/max) | 5.0/66.0 | 22.2/31.8 | 433.0/624.4 | 11:12:36 |
| November/2018 (min/max) | 3.0/73.4 | 19.8/30.1 | 224.4/517.6 | 10:04:37 |
| December/2018 (min/max) | 10.0/71.0 | 15.6/28.0 | 222.8/423.0 | 09:56:06 |
| January/2019 (min/max) | 8.0/69.0 | 14.6/27.8 | 200.0/458.6 | 10:31:53 |
| February/2019 (min/max) | 18.0/61.3 | 13.1/31.1 | 427.0/634.4 | 11:25:28 |
| March 11/2019 (min/max) | 30.0/66.0 | 17.0/30.7 | 351.6/771.2 | 11:48:34 |
| Treatment | K+ | Na+ | Cl− | H2PO4− | Ca2+ | Mg2+ | SO42− | NO3− | TECiw | MECiw | pH |
|---|---|---|---|---|---|---|---|---|---|---|---|
| (mmolc L−1) | (dS m−1) | ||||||||||
| T1 | 0.07 | 3.5 | 0.07 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 1.0 | 1.3 | 6.14 |
| T2 | 0.07 | 30.5 | 27.07 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 4.0 | 4.1 | 6.05 |
| T3 | 0.07 | 60.5 | 57.07 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 7.2 | 7.2 | 6.15 |
| T4 | 0.07 | 120.5 | 117.07 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 13.2 | 12.7 | 6.07 |
| T5 | 0.07 | 160.5 | 157.07 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 17.1 | 16.8 | 6.09 |
| T6 | 0.15 | 3.5 | 0.15 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 1.1 | 1.2 | 6.22 |
| T7 | 0.15 | 30.5 | 27.15 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 4.0 | 4.0 | 6.17 |
| T8 | 0.15 | 60.5 | 57.15 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 7.2 | 7.2 | 6.15 |
| T9 | 0.15 | 120.5 | 117.15 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 13.2 | 13.2 | 6.18 |
| T10 | 0.15 | 160.5 | 157.15 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 17.1 | 16.8 | 6.12 |
| T11 | 0.3 | 3.5 | 0.30 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 1.1 | 1.3 | 6.19 |
| T12 | 0.3 | 30.5 | 27.30 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 4.1 | 4.1 | 6.17 |
| T13 | 0.3 | 60.5 | 57.30 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 7.2 | 7.4 | 6.21 |
| T14 | 0.3 | 120.5 | 117.30 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 13.2 | 13.2 | 6.16 |
| T15 | 0.3 | 160.5 | 157.30 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 17.1 | 16.9 | 6.09 |
| T16 | 3.0 | 0.5 | 0.00 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 1.1 | 1.2 | 6.25 |
| T17 | 3.0 | 27.5 | 27.00 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 4.1 | 4.8 | 6.17 |
| T18 | 3.0 | 57.5 | 57.00 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 7.3 | 7.2 | 6.18 |
| T19 | 3.0 | 117.5 | 117.00 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 13.2 | 13.2 | 6.18 |
| T20 | 3.0 | 157.5 | 157.00 | 0.5 | 5.0 | 2.0 | 2.0 | 8.0 | 17.1 | 16.7 | 6.13 |
| Nutrient | Roots | Shoots | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| NaCl Doses (mmolc L−1) | ||||||||||
| 5.0 | 30.0 | 60.0 | 120.0 | 160.0 | 5.0 | 30.0 | 60.0 | 120.0 | 160.0 | |
| ‘Raccoon’ | ||||||||||
| N (%) | 2.42A | 2.10B | 2.19B | 2.17B | 2.35A | 4.82A | 4.50C | 4.61BC | 4.65ABC | 4.70AB |
| P (g kg−1) | 4.96B | 4.86B | 5.41AB | 5.38B | 5.95A | 4.88AB | 4.74B | 4.88B | 4.92AB | 5.17A |
| Ca (g kg−1) | 9.01A | 7.33B | 6.27C | 4.28D | 4.49D | 15.21A | 10.09B | 7.56C | 7.11C | 7.03C |
| Mg (g kg−1) | 11.50A | 9.74B | 8.65C | 6.73D | 7.59D | 10.95A | 9.78B | 8.84C | 7.88D | 7.55D |
| S (g kg−1) | 5.36A | 3.78B | 3.49B | 2.11C | 2.39C | 4.98A | 4.32B | 4.06C | 3.44D | 3.24D |
| ‘Gazelle’ | ||||||||||
| N (%) | 2.27AB | 2.02D | 2.09CD | 2.23BC | 2.42A | 3.94A | 3.60A | 3.47A | 3.61A | 3.97A |
| P (g kg−1) | 5.32CD | 5.08D | 5.78BC | 6.17AB | 6.62A | 4.57BC | 4.31C | 4.58B | 4.60B | 5.04A |
| Ca (g kg−1) | 8.52A | 6.68B | 5.89C | 4.52D | 4.20D | 10.90A | 7.64B | 6.30C | 5.45D | 5.04D |
| Mg (g kg−1) | 10.62A | 8.85B | 8.29B | 7.48C | 7.57C | 9.71A | 8.32B | 8.10B | 7.32C | 6.94C |
| S (g kg−1) | 4.82A | 3.59B | 3.33B | 2.60C | 2.31C | 4.40A | 4.00B | 3.79B | 3.29C | 3.11C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uçgun, K.; Ferreira, J.F.S.; Liu, X.; da Silva Filho, J.B.; Suarez, D.L.; Lacerda, C.F.d.; Sandhu, D. Germination and Growth of Spinach under Potassium Deficiency and Irrigation with High-Salinity Water. Plants 2020, 9, 1739. https://doi.org/10.3390/plants9121739
Uçgun K, Ferreira JFS, Liu X, da Silva Filho JB, Suarez DL, Lacerda CFd, Sandhu D. Germination and Growth of Spinach under Potassium Deficiency and Irrigation with High-Salinity Water. Plants. 2020; 9(12):1739. https://doi.org/10.3390/plants9121739
Chicago/Turabian StyleUçgun, Kadir, Jorge F. S. Ferreira, Xuan Liu, Jaime Barros da Silva Filho, Donald L. Suarez, Claudivan F. de Lacerda, and Devinder Sandhu. 2020. "Germination and Growth of Spinach under Potassium Deficiency and Irrigation with High-Salinity Water" Plants 9, no. 12: 1739. https://doi.org/10.3390/plants9121739
APA StyleUçgun, K., Ferreira, J. F. S., Liu, X., da Silva Filho, J. B., Suarez, D. L., Lacerda, C. F. d., & Sandhu, D. (2020). Germination and Growth of Spinach under Potassium Deficiency and Irrigation with High-Salinity Water. Plants, 9(12), 1739. https://doi.org/10.3390/plants9121739