Induction of Radiata Pine Somatic Embryogenesis at High Temperatures Provokes a Long-Term Decrease in DNA Methylation/Hydroxymethylation and Differential Expression of Stress-Related Genes

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Global DNA Methylation/Hydroxymethylation Analysis
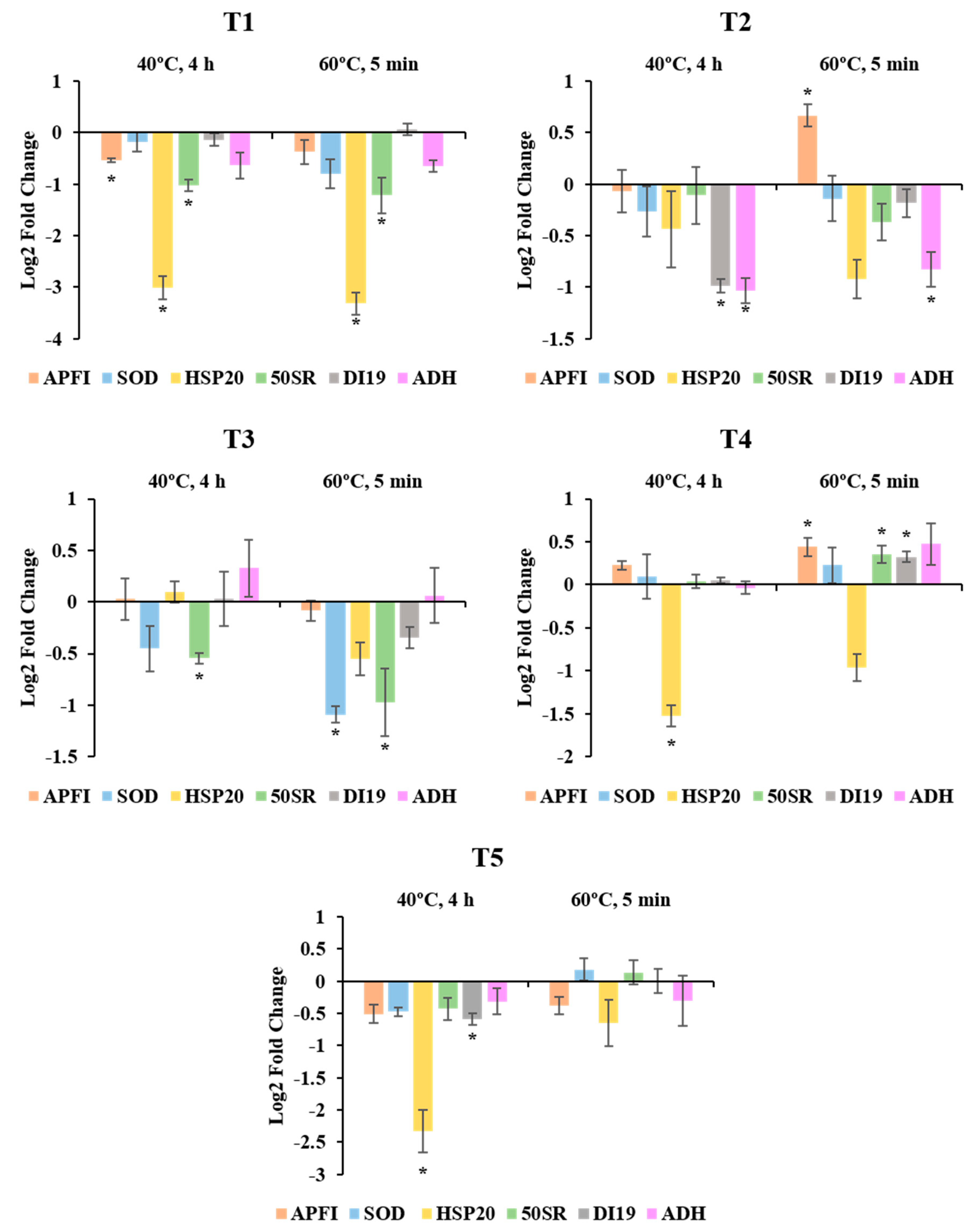
2.2. Expression Pattern of Stress-Related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Heat Stress Experiment
4.2. Global DNA Methylation/Hydroxymethylation Analysis
4.3. RNA Extraction
4.4. Expression Pattern of Stress-Related Genes
- 50S RIBOSOMAL_L6_CHP (50SR): gene encoding a component of the ribosome, part of the translation machinery and responsible for the synthesis of proteins. Previously described as being temperature-responsive and important during early stress events and acclimation in radiata pine [59]. It has also been linked with the response to cytokinins [61], whose profiles were altered by high temperatures in previous studies [25,26].
- SUPEROXIDE DISMUTASE [Cu-Zn] (SOD): encoding an enzyme that catalyses the partitioning of the superoxide (O2−) radical. Heat provokes the synthesis of toxic reactive oxygen species (ROS) and plants require specific enzymes to detoxify these compounds. Previous studies have shown that oxidative stress is the main effect of high temperatures during early stages of radiata pine SE [24].
- TRANSCRIPTION FACTOR APFI (APFI): gene encoding a transcription factor that has been described as an excellent thermo-tolerance biomarker in radiata pine [59].
- HSP20 FAMILY PROTEIN (HSP20): gene coding for a protein member of the small heat shock protein family. These family of proteins have been widely documented as taking active part during heat response and memory acquisition in a great variety of plant species, including radiata pine [59].
- ADH_SF_ZN_TYPE (ADH): encodes an oxidoreductase that catalyses the oxidation of primary alcohols. Other studies have reported enhanced synthesis of this enzyme during heat stress in radiata pine and linked its accumulation with the production of secondary metabolites like flavonoids [59]. The presence of these last compounds was remarked in previous studies after heat exposure in EMs [24].
4.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Iwasaki, M.; Paszkowski, J. Epigenetic memory in plants. EMBO J. 2014, 18, 1987–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, S.; Bertín, A.; Turner, A.; Sepúlveda, F.; Jopia, P.; Parra, M.J.; Castillo, R.; Hasbún, R. Differences in DNA methylation, DNA structure and embryogenesis-related gene expression between embryogenic and non embryogenic lines of Pinus radiata D. don. Plant Cell Tissue Organ. Cult. 2017, 130, 521–529. [Google Scholar] [CrossRef]
- Lämke, J.; Bäurle, I. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 2017, 18, 124. [Google Scholar] [CrossRef] [PubMed]
- Yakovlev, I.A.; Fossdal, C.G. In silico analysis of small RNAs suggest roles for novel and conserved miRNAs in the formation of epigenetic memory in somatic embryos of Norway spruce. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.; Ribeyre, Z.; Vigneaud, J.; Sow, M.D.; Fichot, R.; Messier, C.; Pinto, G.; Nolet, P.; Maury, S. Advances and promises of epigenetics for forest trees. Forests 2020, 11, 976. [Google Scholar] [CrossRef]
- Kumar, S.; Kumari, R.; Sharma, V. Roles, and establishment, maintenance and erasing of the epigenetic cytosine methylation marks in plants. J. Genet. 2013, 92, 629–666. [Google Scholar] [CrossRef]
- Dubin, M.J.; Zhang, P.; Meng, D.; Remigereau, M.S.; Osborne, E.J.; Paolo Casale, F.; Drewe, P.; Kahles, A.; Jean, G.; Vilhjálmsson, B.; et al. DNA methylation in Arabidopsis has a genetic basis and shows evidence of local adaptation. Elife 2015, 4. [Google Scholar] [CrossRef]
- Boyko, A.; Kovalchuk, I. Epigenetic control of plant stress response. Environ. Mol. Mutagen. 2008, 49, 61–72. [Google Scholar] [CrossRef]
- Perrone, A.; Martinelli, F. Plant stress biology in epigenomic era. Plant Sci. 2020, 294, 110376. [Google Scholar] [CrossRef]
- Tittel-Elmer, M.; Bucher, E.; Broger, L.; Mathieu, O.; Paszkowski, J.; Vaillant, I. Stress-induced activation of heterochromatic transcription. PLoS Genet. 2010, 6, e1001175. [Google Scholar] [CrossRef]
- Li, J.; Huang, Q.; Sun, M.; Zhang, T.; Li, H.; Chen, B.; Xu, K.; Gao, G.; Li, F.; Yan, G.; et al. Global DNA methylation variations after short-term heat shock treatment in cultured microspores of Brassica napus cv. Topas. Sci. Rep. 2016, 6, 38401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sow, M.D.; Allona, I.; Ambroise, C.; Conde, D.; Fichot, R.; Gribkova, S.; Jorge, V.; Le-Provost, G.; Pâques, L.; Plomion, C.; et al. Epigenetics in forest trees: State of the art and potential implications for breeding and management in a context of climate change. In Advances in Botanical Research. Plant Epigenetics Coming of Age for Breeding Applications; Gallusci, P., Bucher, E., Mirouze, M., Eds.; Academic Press, Elsevier: Amsterdam, The Netherlands, 2018; Volume 88, pp. 387–453. [Google Scholar] [CrossRef]
- Blödner, C.; Skroppa, T.; Johnsen, Ø.; Polle, A. Freezing tolerance in two Norway spruce (Picea abies [L.] Karst.) progenies is physiologically correlated with drought tolerance. J. Plant Physiol. 2005, 162, 549–558. [Google Scholar] [CrossRef]
- Bouché, F.; Woods, D.P.; Amasino, R.M. Winter memory throughout the plant kingdom: Different paths to flowering. Plant Physiol. 2017, 173, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrath, U.; Beckers, G.J.; Langenbach, C.J.; Jaskiewicz, M.R. Priming for enhanced defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Barua, P.; Chakraborty, N.; Nandi, A.K. Systemic acquired resistance specific proteome of Arabidopsis thaliana. Plant Cell Rep. 2020, 39, 1549–1563. [Google Scholar] [CrossRef] [PubMed]
- Morcillo, M.; Sales, E.; Ponce, L.; Guillén, A.; Segura, J.; Arrillaga, I. Effect of elicitors on holm oak somatic embryo development and efficacy inducing tolerance to Phytophthora cinnamomi. Sci. Rep. 2020, 10, 15166. [Google Scholar] [CrossRef]
- Kvaalen, H.; Johnsen, O. Timing of bud set in Picea abies is regulated by a memory of temperature during zygotic and somatic embryogenesis. New Phytol. 2008, 177, 49–59. [Google Scholar] [CrossRef]
- Le Gac, A.L.; Lafon-Placette, C.; Chauveau, D.; Segura, V.; Delaunay, A.; Fichot, R.; Marron, N.; Le Jan, I.; Berthelot, A.; Bodineau, G.; et al. Winter-dormant shoot apical meristem in poplar trees shows environmental epigenetic memory. J. Exp. Bot. 2018, 14, 4821–4837. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.Q.; Ali, I.; Tang, J.; Yang, W.C. New insights into 5hmC DNA modification: Generation, distribution and function. Front. Genet. 2017, 8, 100. [Google Scholar] [CrossRef]
- Moricová, P.; Ondřej, V.; Navrátilová, B.; Luhová, L. Changes of DNA methylation and hydroxymethylation in plant protoplast cultures. Acta Biochim. Pol. 2013, 60, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.L.; Song, S.H.; Wu, Y.S.; Li, Y.L.; Chen, T.T.; Huang, Z.Y.; Liu, S.; Dunwell, T.L.; Pfeifer, G.P.; Dunwell, J.M.; et al. Genome-wide mapping of 5-hydroxymethylcytosine in three rice cultivars reveals its preferential localization in transcriptionally silent transposable element genes. J. Exp. Bot. 2015, 66, 6651–6663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakovlev, I.A.; Gackowski, D.; Abakir, A.; Viejo, M.; Ruzov, A.; Olinski, R.; Starczak, M.; Fossdal, C.G.; Krutovsky, K.V. Mass spectrometry reveals the presence of specific set of epigenetic DNA modifications in the Norway spruce genome. Sci. Rep. 2019, 9, 19314. [Google Scholar] [CrossRef] [PubMed]
- Castander-Olarieta, A.; Montalbán, I.A.; De Medeiros Oliveira, E.; Dell’Aversana, E.; D’Amelia, L.; Carillo, P.; Steiner, N.; Fraga, H.P.D.F.; Guerra, M.P.; Goicoa, T.; et al. Effect of thermal stress on tissue ultrastructure and metabolite profiles during initiation of radiata pine somatic embryogenesis. Front. Plant. Sci. 2019, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Castander-Olarieta, A.; Moncaleán, P.; Pereira, C.; Pěnčík, A.; Petřík, I.; Pavlović, I.; Novák, O.; Strnad, M.; Goicoa, T.; Ugarte, M.D.; et al. Cytokinins are involved in drought tolerance of Pinus radiata plants originating from embryonal masses induced at high temperatures. Tree Physiol. 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Castander-Olarieta, A.; Pereira, C.; Montalbán, I.A.; Pěnčík, A.; Petřík, I.; Pavlović, I.; Novák, O.; Strnad, M.; Moncaleán, P. Quantification of endogenous aromatic cytokinins in Pinus radiata embryonal masses after application of heat stress during initiation of somatic embryogenesis. Trees 2020. [Google Scholar] [CrossRef]
- Pereira, C.; Castander-Olarieta, A.; Montalbán, I.A.; Pěnčík, A.; Petřík, I.; Pavlović, I.; De Medeiros Oliveira, E.; Fraga, H.P.D.F.; Guerra, M.P.; Novák, O.; et al. Embryonal masses induced at high temperatures in Aleppo pine: Cytokinin profile and cytological characterization. Forests 2020, 11, 807. [Google Scholar] [CrossRef]
- Fernandez, A.F.; Valledor, L.; Vallejo, F.; Cañal, M.J.; Fraga, M.F. Quantification of global DNA methylation levels by mass spectrometry. In DNA Methylation Protocols. Methods in Molecular Biology; Tost, J., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1708, pp. 49–58. [Google Scholar] [CrossRef]
- Santamaría, M.E.; Hasbún, R.; Valera, M.J.; Meijón, M.; Valledor, L.; Rodríguez, J.L.; Toorop, P.E.; Cañal, M.J.; Rodríguez, R. Acetylated H4 histone and genomic DNA methylation patterns during bud set and bud burst in Castanea sativa. J. Plant Physiol. 2009, 166, 1360–1369. [Google Scholar] [CrossRef]
- Yakovlev, I.A.; Carneros, E.; Lee, Y.; Olsen, J.E.; Fossdal, C.G. Transcriptional profiling of epigenetic regulators in somatic embryos during temperature induced formation of an epigenetic memory in Norway spruce. Planta 2016, 243, 1237–1249. [Google Scholar] [CrossRef]
- García-Mendiguren, O.; Montalbán, I.A.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P. Are we able to modulate the response of somatic embryos of pines to drought stress? Act. Hortic. 2017, 1155, 77–84. [Google Scholar] [CrossRef]
- Noceda, C.; Salaj, T.; Pérez, M.; Viejo, M.; Cañal, M.J.; Salaj, J.; Rodríguez, R. DNA demethylation and decrease on free polyamines is associated with the embryogenic capacity of Pinus nigra Arn. cell culture. Trees 2009, 23, 1285. [Google Scholar] [CrossRef]
- De-la-Peña, C.; Nic-Can, G.I.; Galaz-Ávalos, R.M.; Avilez-Montalvo, R.; Loyola-Vargas, V.M. The role of chromatin modifications in somatic embryogenesis in plants. Front. Plant Sci. 2015, 6, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miguel, C.; Marum, L. An epigenetic view of plant cells cultured in vitro: Somaclonal variation and beyond. J. Exp. Bot. 2011, 62, 3713–3725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corredoira, E.; Cano, V.; Bárány, I.; Solís, M.T.; Rodríguez, H.; Vieitez, A.M.; Risueño, M.C.; Testillano, P.S. Initiation of leaf somatic embryogenesis involves high pectin esterification, auxin accumulation and DNA demethylation in Quercus alba. J. Plant Physiol. 2017, 213, 42–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraga, H.P.F.; Vieira, L.N.; Caprestano, C.A.; Steinmacher, D.A.; Micke, G.A.; Spudeit, D.A.; Pescador, R.; Guerra, M.P. 5-Azacytidine combined with 2,4-D improves somatic embryogenesis of Acca sellowiana (O. Berg) Burret by means of changes in global DNA methylation levels. Plant Cell Rep. 2012, 31, 2165–2176. [Google Scholar] [CrossRef]
- Teyssier, C.; Maury, S.; Beaufour, M.; Grondin, C.; Delaunay, A.; Le Metté, C.; Ader, K.; Cadene, M.; Label, P.; Lelu-Walter, M.A. In search of markers for somatic embryo maturation in hybrid larch (Larix × eurolepis): Global DNA methylation and proteomic analyses. Physiol. Plant 2014, 150, 271–291. [Google Scholar] [CrossRef]
- Solís, M.T.; Rodríguez-Serrano, M.; Meijón, M.; Cañal, M.J.; Cifuentes, A.; Risueño, M.C.; Testillano, P.S. DNA methylation dynamics and MET1a-like gene expression changes during stress-induced pollen reprogramming to embryogenesis. J. Exp. Bot. 2012, 63, 6431–6444. [Google Scholar] [CrossRef]
- Che, P.; Love, T.M.; Frame, B.R.; Wang, K.; Carriquiry, A.L.; Howell, S.H. Gene expression patterns during somatic embryo eevelopment and germination in maize Hi II callus cultures. Plant Mol. Biol. 2006, 62, 1–14. [Google Scholar] [CrossRef]
- Fraga, M.F.; Rodríguez, R.; Cañal, M.J. Genomic DNA methylation–demethylation during aging and reinvigoration of Pinus radiata. Tree Physiol. 2002, 22, 813–816. [Google Scholar] [CrossRef] [Green Version]
- Fraga, M.F.; Cañal, M.J.; Rodríguez, R. Phase-change related epigenetic and physiological changes in Pinus radiata D. Don. Planta 2002, 215, 672–678. [Google Scholar] [CrossRef]
- Berdasco, M.; Alcázar, R.; García-Ortiz, M.V.; Ballestar, E.; Fernández, A.F.; Roldán-Arjona, T. Promoter DNA hypermethylation and gene repression in undifferentiated Arabidopsis cells. PLoS ONE 2008, 3, e3306. [Google Scholar] [CrossRef] [Green Version]
- Boyko, A.; Blevins, T.; Yao, Y.; Golubov, A.; Bilichak, A.; Ilnytskyy, Y.; Hollander, J.; Meins Jr, F.; Kovalchuk, I. Transgenerational adaptation of Arabidopsis to stress requires DNA methylation and the function of dicer-like proteins. PLoS ONE 2010, 5, e9514. [Google Scholar] [CrossRef]
- Johnsen, Ø.; Fossdal, C.G.; Nagy, N.; Mølmann, J.; Dæhlen, O.G.; Skrøppa, T. Climatic adaptation in Picea abies progenies is affected by the temperature during zygotic embryogenesis and seed maturation. Plant Cell Environ. 2005, 28, 1090–1102. [Google Scholar] [CrossRef]
- Alakärppä, E.; Salo, H.M.; Valledor, L.; Cañal, M.J.; Häggman, H.; Vuosku, J. Natural variation of DNA methylation and gene expression may determine local adaptations of Scots pine populations. J. Exp. Bot. 2018, 69, 5293–5305. [Google Scholar] [CrossRef] [PubMed]
- Min, L.; Li, Y.; Hu, Q.; Zhu, L.; Gao, W.; Wu, Y.; Ding, Y.; Liu, S.; Yang, X.; Zhang, X. Sugar and auxin signaling pathways respond to high-temperature stress during anther development as revealed by transcript profiling analysis in cotton. Plant Physiol. 2014, 164, 1293–1308. [Google Scholar] [CrossRef] [Green Version]
- Browne, L.; Mead, A.; Horn, C.; Chang, K.; Celikkol, Z.A.; Henriquez, C.L.; Ma, F.; Beraut, E.; Meyer, R.S.; Sork, V.L. Experimental DNA demethylation associates with changes in growth and gene expression of oak tree seedlings. G3 2020, 10, 1019–1028. [Google Scholar] [CrossRef] [Green Version]
- Sow, M.D.; Le Gac, A.L.; Fichot, R.; Lanciano, S.; Delaunay, A.; Le Jan, I.; Lesage-Descauses, M.C.; Citerne, S.; Caius, J.; Brunaud, V.; et al. Hypomethylated poplars show higher tolerance to water deficit and highlight a dual role for DNA methylation in shoot meristem: Regulation of stress response and genome integrity. bioRxiv 2020, 045328. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.; Shin, H.; Eichman, B.F.; Huh, J.H. Excision of 5-hydroxymethylcytosine by DEMETER family DNA glycosylases. Biochem. Biophys. Res. Commun. 2014, 446, 1067–1072. [Google Scholar] [CrossRef] [Green Version]
- Nystedt, B.; Street, N.; Wetterbom, A.; Zuccolo, A.; Lin, Y.C.; Scofield, D.G.; Vezzi, F.; Delhomme, M.; Giacomello, S.; Alexeyenko, A.; et al. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef] [Green Version]
- Valinluck, V.; Tsai, H.H.; Rogstad, D.K.; Burdzy, A.; Bird, A.; Sowers, L.C. Oxidative damage to methyl-CpG sequences inhibits the binding of the methyl-CpG binding domain (MBD) of methyl-CpG binding protein 2 (MeCP2). Nucleic Acids Res. 2004, 32, 4100–4108. [Google Scholar] [CrossRef] [Green Version]
- Nestor, C.E.; Ottaviano, R.; Reddington, J.; Sproul, D.; Reinhardt, D.; Dunican, D.; Katz, E.; Dixon, J.M.; Harrison, D.J.; Meehan, R.R. Tissue type is a major modifier of the 5-hydroxymethylcytosine content of human genes. Genome Res. 2012, 3, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Bokszczanin, K.L.; Fragkostefanakis, S. Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Front. Plant Sci. 2013, 4, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Taïbi, K.; del Campo, A.D.; Aguado, A.; Mulet, J.M. The effect of genotype by environment interaction, phenotypic plasticity and adaptation on Pinus halepensis reforestation establishment under expected climate drifts. Ecol. Eng. 2015, 84, 218–228. [Google Scholar] [CrossRef]
- Sedaghatmehr, M.; Thirumalaikumar, V.P.; Kamranfar, I.; Marmagne, A.; Masclaux-Daubresse, C.; Balazadeh, S. A regulatory role of autophagy for resetting the memory of heat stress in plants. Plant Cell Environ. 2019, 42, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Thirumalaikumar, V.P.; Karina Schulz, M.G.; Masclaux-Daubresse, C.; Sampathkumar, A.; Skirycz, A.; Vierstra, R.D.; Balazadeh, S. Selective autophagy regulates heat stress memory in Arabidopsis by NBR1-mediated targeting of HSP90 and ROF1. Autophagy 2020. [Google Scholar] [CrossRef] [PubMed]
- Ujino-Ihara, T. Transcriptome analysis of heat stressed seedlings with or without pre-heat treatment in Cryptomeria japonica. Mol. Genet. Genom. 2020, 295, 1163–1172. [Google Scholar] [CrossRef]
- Wang, X.; Li, Z.; Liu, B.; Zhou, H.; Elmongy, M.S.; Xia, Y. Combined proteome and transcriptome analysis of heat-primed azalea reveals new insights into plant heat acclimation memory. Front. Plant Sci. 2020, 11, 1278. [Google Scholar] [CrossRef]
- Escandón, M.; Valledor, L.; Pascual, J.; Pinto, G.; Cañal, M.J.; Meijón, M. System-wide analysis of short-term response to high temperature in Pinus radiata. J. Exp. Bot. 2017, 68, 3629–3641. [Google Scholar] [CrossRef]
- Businge, E.; Egertsdotter, U. A possible biochemical basis for fructose-induced inhibition of embryo development in Norway spruce (Picea abies). Tree Physiol. 2014, 34, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Černý, M.; Kuklová, A.; Hoehenwarter, W.; Fragner, L.; Novák, O.; Rotková, G.; Jedelský, P.L.; Žáková, K.; Šmehilová, M.; Strnad, M.; et al. Proteome and metabolome profiling of cytokinin action in Arabidopsis identifying both distinct and similar responses to cytokinin down- and up-regulation. J. Exp. Bot. 2013, 64, 4193–4206. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.X.; Li, Y.; Li, D.D.; Xu, W.L.; Zheng, Y.; Li, X.B. Arabidopsis drought-induced protein Di19-3 participates in plant response to drought and high salinity stresses. Plant Mol. Biol. 2014, 6, 609–625. [Google Scholar] [CrossRef] [PubMed]
- Milla, M.A.R.; Townsend, J.; Chang, I.; Cushman, J.C. The Arabidopsis AtDi19 gene family encodes a novel type of Cys2/His2 zinc-finger protein implicated in ABA-independent dehydration, high-salinity stress and light signaling pathways. Plant Mol. Biol. 2006, 61, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Montalbán, I.A.; de Diego, N.; Moncaleán, P. Enhancing initiation and proliferation in radiata pine (Pinus radiata D. Don) somatic embryogenesis through seed family screening, zygotic embryo staging and media adjustments. Acta Physiol. Plant 2012, 34, 451–460. [Google Scholar] [CrossRef]
- Walter, C.; Find, J.I.; Grace, L.J. Somatic embryogenesis and genetic transformation in Pinus radiata. In Protocol for Somatic Embryogenesis in Woody Plants; Jain, S.M., Gupta, P.K., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 11–24. [Google Scholar]
- Montalbán, I.A.; Moncaleán, P. Pinus radiata (D. Don) somatic embryogenesis. In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants; Jain, S., Gupta, P., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–11. [Google Scholar] [CrossRef]
- Quoirin, M.; Lepoivre, P. Études des milieux adaptés aux cultures in vitro de Prunus. Acta Hort. 1977, 78, 437–442. [Google Scholar] [CrossRef]
- Aitken-Christie, J.; Singh, A.P.; Davies, H. Multiplication of meristematic tissue: A new tissue culture system for radiata pine. In Genetic Manipulation of Woody Plants; Hanover, J.W., Keathley, D.E., Eds.; Plenum: New York, NY, USA, 1988; pp. 413–432. [Google Scholar] [CrossRef]
- Valledor, L.; Escandón, M.; Meijón, M.; Nukarinen, E.; Cañal, M.J.; Weckwerth, W. A universal protocol for the combined isolation of metabolites, DNA, long RNAs, small RNAs, and proteins from plants and microorganisms. Plant J. 2014, 79, 173–180. [Google Scholar] [CrossRef]
- Alvarez, C.; Valledor, L.; Sáez, P.; Hasbún, R.; Sánchez-Olate, M.; Cañal, M.J.; Ríos, D. Changes in gene expression in needles and stems of Pinus radiata rootstock plants of different ontogenic age. Am. J. Plant Sci. 2016, 7, 1205–1216. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 4, 402–408. [Google Scholar] [CrossRef]


{kind=link}
| T2 | T5 | ||||
|---|---|---|---|---|---|
| Treatment | 5mC % | Treatment | 5mC % | 5hmC % | 5hmC/5mC × 100 |
| 23 ° C, 8 weeks | 39.28 ± 1.06 a | 23 ° C, 8 weeks | 40.45 ± 0.53 a | 0.0175 ± 0.0024 a | 0.044 ± 0.0071 a |
| 40 ° C, 4 h | 40.85 ± 0.36 a | 40 ° C, 4 h | 40.39 ± 0.22 a | 0.0103 ± 0.0005 b | 0.026 ± 0.0012 b |
| 60 ° C, 5 min | 37.82 ± 1.06 a | 60 ° C, 5 min | 37.64 ± 0.33 b | 0.0101 ± 0.0003 b | 0.027 ± 0.0009 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castander-Olarieta, A.; Pereira, C.; Sales, E.; Meijón, M.; Arrillaga, I.; Cañal, M.J.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P.; Montalbán, I.A. Induction of Radiata Pine Somatic Embryogenesis at High Temperatures Provokes a Long-Term Decrease in DNA Methylation/Hydroxymethylation and Differential Expression of Stress-Related Genes. Plants 2020, 9, 1762. https://doi.org/10.3390/plants9121762
Castander-Olarieta A, Pereira C, Sales E, Meijón M, Arrillaga I, Cañal MJ, Goicoa T, Ugarte MD, Moncaleán P, Montalbán IA. Induction of Radiata Pine Somatic Embryogenesis at High Temperatures Provokes a Long-Term Decrease in DNA Methylation/Hydroxymethylation and Differential Expression of Stress-Related Genes. Plants. 2020; 9(12):1762. https://doi.org/10.3390/plants9121762
Chicago/Turabian StyleCastander-Olarieta, Ander, Cátia Pereira, Ester Sales, Mónica Meijón, Isabel Arrillaga, María Jesús Cañal, Tomás Goicoa, María Dolores Ugarte, Paloma Moncaleán, and Itziar A. Montalbán. 2020. "Induction of Radiata Pine Somatic Embryogenesis at High Temperatures Provokes a Long-Term Decrease in DNA Methylation/Hydroxymethylation and Differential Expression of Stress-Related Genes" Plants 9, no. 12: 1762. https://doi.org/10.3390/plants9121762
APA StyleCastander-Olarieta, A., Pereira, C., Sales, E., Meijón, M., Arrillaga, I., Cañal, M. J., Goicoa, T., Ugarte, M. D., Moncaleán, P., & Montalbán, I. A. (2020). Induction of Radiata Pine Somatic Embryogenesis at High Temperatures Provokes a Long-Term Decrease in DNA Methylation/Hydroxymethylation and Differential Expression of Stress-Related Genes. Plants, 9(12), 1762. https://doi.org/10.3390/plants9121762