Effect of Rhododendron arboreum Leaf Extract on the Antioxidant Defense System against Chromium (VI) Stress in Vigna radiata Plants



Abstract
:1. Introduction
2. Materials and Methods
2.1. Raising of Plant Material
2.2. Rhododendron arboreum Methanol Leaf Extract(MEL)
2.3. Cr(VI)Metal
2.4. Treatments
2.5. Phenolic Pigments:
2.5.1. Xanthophyll Content
2.5.2. Anthocyanin Content
2.6. Protein Content
2.7. Enzymatic Antioxidants
2.7.1. Preparation of Plant Extracts
2.7.2. Guaiacol Peroxidase (POD, EC. 1.11.1.7)
2.7.3. Catalase (CAT, EC 1.11.1.6)
2.7.4. Superoxide Dismutase (SOD, EC 1.15.1.1)
2.7.5. Ascorbate Peroxidase (APOX, EC. 1.11.1.11)
2.7.6. Glutathione Reductase (GR, EC 1.6.4.2)
2.7.7. Dehydroascorbate Reductase (DHAR, EC. 1.8.5.1)
2.7.8. Polyphenol Oxidase (PPO, EC 1.10.3.1)
2.7.9. Glutathione-S-Transferase (GST, EC 2.5.1.13)
2.7.10. Glutathione Peroxidase (GPOX, EC 1.11.1.7)
2.8. Non-Enzymatic Antioxidants
2.8.1. Ascorbic Acid Content
2.8.2. Tocopherol Content
2.8.3. Glutathione Content
2.8.4. Glutathione Imaging
2.9. Polyphenol Estimation
2.10. Amino Acid Profiling
2.11. Organic Acid Profiling
2.12. Statistical Analysis
3. Results
3.1. Phenolic Pigments
3.2. Protein Content
3.3. Enzymatic Antioxidants
3.4. Non-Enzymatic Antioxidants
3.5. Glutathione Imaging
3.6. Polyphenol Content
3.7. Amino Acid Content
3.8. Organic Acids
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hasanuzzaman, M.; Fujita, M. Heavy metals in the environment: Current status, toxic effects on plants and possible phytoremediation. In Phytotechnologies: Remediation of Environmental Contaminants; Anjum, N.A., Pereira, M.A., Ahmad, I., Duarte, A.C., Umar, S., Khan, N.A., Eds.; Taylor and Francis/CRC Press: Boca Raton, FL, USA, 2012; pp. 7–73. [Google Scholar]
- Jomovaa, K.; Valkob, M. Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Dheeba, B.; Sampathkumar, P. Ameliorating effect of fertilizers on biochemical characteristics of Vigna radiata treated with hexavalent chromium. Biosci. Biotechnol. Res. Asia 2014, 11, 301–307. [Google Scholar] [CrossRef]
- Oliveira, H. Chromium as an environmental pollutant: Insights on induced plant toxicity. J. Bot. 2012. [Google Scholar] [CrossRef]
- Panda, S.K. Chromium-mediated oxidative stress and ultrastructural changes in root cells of developing rice seedlings. J. Plant Physiol. 2007, 164, 1419–1428. [Google Scholar] [CrossRef]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Niwa, T.; Doi, U.; Kato, Y.; Osawa, T. Antioxidative Properties of Phenolic Antioxidants Isolated from Corn Steep Liquor. J. Agric. Food Chem. 2001, 49, 177–182. [Google Scholar] [CrossRef]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef] [Green Version]
- Andresen, M.; Cedergreen, N. Plant growth is stimulated by tea-seed extract: A new natural growth regulator? Hortscience 2010, 45, 1848–1853. [Google Scholar] [CrossRef] [Green Version]
- Gautam, V.; Kohli, S.; Arora, S.; Bhardwaj, R.; Kazi, M.; Ahmad, A.; Raish, M.; Ganaie, M.; Ahmad, P. Antioxidant and Antimutagenic Activities of Different Fractions from the Leaves of Rhododendron arboreum Sm. and Their GC-MS Profiling. Molecules 2018, 23, 2239. [Google Scholar] [CrossRef] [Green Version]
- Painuli, S.; Rai, N.; Kumar, N. Gas chromatography and mass spectrometry analysis of methanolic extract of leaves of Rhododendron arboreum. Asian J. Pharm. Clin. Res. 2016, 9, 66–69. [Google Scholar]
- Roy, J.D.; Handique, A.K.; Barua, C.C.; Talukdar, A.; Ahmed, F.A.; Barua, I.C. Evaluation of phytoconstituents and assessment of adaptogenic activity in vivo in various extracts of Rhododendron arboreum(leaves). J. Pharm. Biol. Res. 2014, 2, 49–56. [Google Scholar] [CrossRef]
- Kiruba, S.; Mahesh, M.; Nisha, S.R.; Paul, Z.M.; Jeeva, S. Phytochemical analysis of the flower extracts of Rhododendron arboreum Sm. ssp. nilagiricum (Zenker) Tagg. Asian Pac. J. Trop. Biomed. 2011, 1, S284–S286. [Google Scholar] [CrossRef]
- Rath, B.P.; Hota, S.; Subhadarshini, S.; Dash, D.; Das, P.K. Consequence of chromium-tainted soil on physical and biochemical responses of Vigna radiata L. J. Appl. Biol. Biotechnol. 2019, 7, 35–41. [Google Scholar]
- Cobbett, C.S. Heavy metals and plants-model systems and hyper accumulators. New Phytol. 2003, 159, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, J. Determination of total xanthophyll and marigold oleoresin. J. Ass. Off. Anal. Chem. 1990, 2, 970–975. [Google Scholar]
- Mancinelli, A.L. Photoregulation of Anthocyanin Synthesis. Plant Physiol. 1984, 75, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Pütter, J. Peroxidases. In Methods of Enzymatic Analysis; Elsevier: Amsterdam, The Netherlands, 1974; pp. 685–690. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; pp. 121–126. [Google Scholar] [CrossRef]
- Kono, Y. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen-Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach-Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Purification and characterization of the flavoenzyme glutathione reductase from rat liver. J. Biol. Chem. 1975, 250, 5475–5480. [Google Scholar] [PubMed]
- Dalton, D.A.; Russell, S.A.; Hanus, F.J.; Pascoe, G.A.; Evans, H.J. Enzymatic reactions of ascorbate and glutathione that prevent peroxide damage in soybean root nodules. Proc. Natl. Acad. Sci. USA 1986, 83, 3811–3815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, K.; Khan, P. Age-related changes in catalase and peroxidase activities in the excised leaves of Eleusine coracana Gaertn. cv PR 202 during senescence. Exp. Gerontol. 1982, 18, 409–417. [Google Scholar] [CrossRef]
- Habig, W.H.; Jakoby, W.B. Assays for differentiation of glutathione S-Transferases. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1981; pp. 398–405. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. Assays of glutathione peroxidase. In Methods in enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 114–120. [Google Scholar]
- Roe, J.H.; Kuether, C.A. The determination of ascorbic acid in whole blood and urine through the 2, 4-dinitrophenylhydrazine derivavative of dehydroascorbic acid. J. Biol. Chem. 1943, 147, 399–407. [Google Scholar]
- Martinek, R.G. Method for the determination of vitamin E (total tocopherols) in serum. Clin. Chem. 1964, 10, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Hartmann, T.N.; Fricker, M.D.; Rennenberg, H.; Meyer, A.J. Cell-specific measurement of cytosolic glutathione in poplar leaves. Plant Cell Environ. 2003, 26, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Iriti, M.; Rossoni, M.; Borgo, M.; Ferrara, L.; Faoro, F. Induction of Resistance to Gray Mold with Benzothiadiazole Modifies Amino Acid Profile and Increases Proanthocyanidins in Grape: Primary versus Secondary Metabolism. J. Agric. Food Chem. 2005, 53, 9133–9139. [Google Scholar] [CrossRef]
- Chen, M.-C.; Wang, M.-K.; Chiu, C.-Y.; Huang, P.-M.; King, H.-B. Determination of low molecular weight dicarboxylic acids and organic functional groups in rhizosphere and bulk soils of Tsuga and Yushania in a temperate rain forest. Plant Soil 2001, 231, 37–44. [Google Scholar] [CrossRef]
- Bailey, N.T. Statistical Methods in Biology; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- El-Soud, W.A.; Hegab, M.M.; AbdElgawad, H.; Zinta, G.; Asard, H. Ability of ellagic acid to alleviate osmotic stress on chickpea seedlings. Plant Physiol. Biochem. 2013, 71, 173–183. [Google Scholar] [CrossRef]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Interaction of 24-epibrassinolide and salicylic acid regulates pigment contents, antioxidative defense responses, and gene expression in Brassica juncea L. seedlings under Pb stress. Environ. Sci. Pollut. Res. 2018, 25, 15159–15173. [Google Scholar] [CrossRef] [PubMed]
- Posmyk, M.M.; Kontek, R.; Janas, K.M. Antioxidant enzymes activity and phenolic compounds content in red cabbage seedlings exposed to copper stress. Ecotoxicol. Environ. Saf. 2009, 72, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Kovinich, N.; Kayanja, G.; Chanoca, A.; Otegui, M.S.; Grotewold, E. Abiotic stresses induce different localizations of anthocyanins in Arabidopsis. Plant Signal Behav. 2015, 10, e1027850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Mahmood, K.; Rothstein, S.J. ROS Induces Anthocyanin Production Via Late Biosynthetic Genes and Anthocyanin Deficiency Confers the Hypersensitivity to ROS-Generating Stresses in Arabidopsis. Plant Cell Physiol. 2017, 58, 1364–1377. [Google Scholar] [CrossRef]
- Baek, S.-A.; Han, T.; Ahn, S.-K.; Kang, H.; Cho, M.R.; Lee, S.-C.; Im, K.-H. Effects of Heavy Metals on Plant Growths and Pigment Contents in Arabidopsis thaliana. Plant Pathol. J. 2012, 28, 446–452. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Xie, X.; Wang, G. Effects of the heavy metal cadmium on photosynthetic activity and the xanthophyll cycle in Phaeodactylum tricornutum. J. Oceanol. Limnol. 2018, 36, 2194–2201. [Google Scholar] [CrossRef]
- Ahmad, S.; Kamran, M.; Ding, R.; Meng, X.; Wang, H.; Ahmad, I.; Fahad, S.; Han, Q. Exogenous melatonin confers drought stress by promoting plant growth, photosynthetic capacity and antioxidant defense system of maize seedlings. PeerJ 2019, 7, e7793. [Google Scholar] [CrossRef] [Green Version]
- Rai, V.; Vajpayee, P.; Singh, S.N.; Mehrotra, S. Effect of chromium accumulation on photosynthetic pigments, oxidative stress defense system, nitrate reduction, proline level and eugenol content of Ocimum tenuiflorum L. Plant Sci. 2004, 167, 1159–1169. [Google Scholar] [CrossRef]
- Hameed, A.; Gulzar, S.; Aziz, I.; Hussain, T.; Gul, B.; Khan, M.A. Effects of salinity and ascorbic acid on growth, water status and antioxidant system in a perennial halophyte. AoB PLANTS 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Dey, S.; Sidor, A.; O’Rourke, B. Compartment-specific Control of Reactive Oxygen Species Scavenging by Antioxidant Pathway Enzymes. J. Biol. Chem. 2016, 291, 11185–11197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, V.C.; Eymery, F.; Genty, B.; Rey, P.; Havaux, M. Vitamin E is essential for the tolerance of Arabidopsis thaliana to metal-induced oxidative stress. Plant Cell Environ. 2008, 31, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Piyatida, P.; Teixeira-da-Silva, J.A.; Fujita, M. Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J. Bot. 2012, 2012, 872875. [Google Scholar] [CrossRef]
- Gallie, D.R. L-Ascorbic Acid: A Multifunctional Molecule Supporting Plant Growth and Development. Scientifica 2013, 2013, 795964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azooz, M.M.; Hassanein, A.M.; Faheed, F.A. Riboflavin (vitamin B2) treatments counteract the adverse effects of salinity on growth and some relevant physiological responses of Hibiscus sabdariffa L. seedlings. Bull. Fac. Sci. Assuit Univ. 2002, 31, 395–403. [Google Scholar]
- Barakat, H. Interactive effects of salinity and certain vitamin on gene expression and cell division. Int. J. Agric. Biol. 2003, 3, 219–225. [Google Scholar]
- Cvelkorska, M.; Rampitsch, C.; Bykova, N.; Xing, T. Genomic analysis of MAP kinase cascades in Arabidopsis defense responses Plant Mol. Biol. Report. 2005, 23, 331–343. [Google Scholar]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Mahmud, J.A.; Alharby, H.F.; Fujita, M. Exogenous glutathione attenuates lead-induced oxidative stress in wheat by improving antioxidant defense and physiological mechanisms. J. Plant Interact. 2018, 13, 203–212. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Xi, Z.-m.; Zhang, Z.-w.; Huo, S.-s.; Luan, L.-y.; Gao, X.; Ma, L.-n.; Fang, Y.-l. Regulating the secondary metabolism in grape berry using exogenous 24-epibrassinolide for enhanced phenolics content and antioxidant capacity. Food Chem. 2013, 141, 3056–3065. [Google Scholar] [CrossRef] [PubMed]
- Agada, O.O. Abiotic Stress, Antioxidants and Crop Productivity: The Mitigating Role of Exogenous Substances. Greener J. Agric. Sci. 2016, 6, 79–86. [Google Scholar] [CrossRef]
- Alhasnawi, A.N.; Kadhimi, A.A.; Isahak, A.; Mohamad, A.; Yusoff, W.M.W.; Zain, C.R.C.M. Exogenous Application of Ascorbic Acid Ameliorates Detrimental Effects of Salt Stress in Rice (MRQ74 and MR269) Seedlings. Asian J. Crop Sci. 2015, 7, 186–196. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.; Maeder, V.; Funk, F.; Frey, B.; Sticher, H.; Frossard, E. Release of phenols from Lupinus albus L. roots exposed to Cu and their possible role in Cu detoxification. Plant Soil 2003, 252, 301–312. [Google Scholar] [CrossRef]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Combined effect of 24-epibrassinolide and salicylic acid mitigates lead (Pb) toxicity by modulating various metabolites in Brassica juncea L. seedlings. Protoplasma 2018, 255, 11–24. [Google Scholar] [CrossRef]
- Ma, J.F. Role of Organic Acids in Detoxification of Aluminum in Higher Plants. Plant Cell Physiol. 2000, 41, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Callahan, D.L.; Baker, A.J.M.; Kolev, S.D.; Wedd, A.G. Metal ion ligands in hyperaccumulating plants. J. Biol. Inorg. Chem. 2006, 11, 2–12. [Google Scholar] [CrossRef]
- Yusuf, M.; Fariduddin, Q.; Ahmad, A. 24-Epibrassinolide modulates growth, nodulation, antioxidant system, and osmolyte in tolerant and sensitive varieties of Vigna radiata under different levels of nickel: A shotgun approach. Plant Physiol. Biochem. 2012, 57, 143–153. [Google Scholar] [CrossRef]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics, and ionomics. Front. Plant Sci. 2015, 6, 1143. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Yang, X.E.; Jin, X.F.; Feng, Y.; Islam, E. Molecular mechanisms and genetic basis of heavy metal tolerance/hyperaccumulation in plants. J. Integr. Plant Biol. 2005, 47, 1025–1035. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S. Molecular mechanisms of plant metal tolerance and homeostasis. Planta 2001, 212, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Oven, M.; Grill, E.; Golan-Goldhirsh, A.; Kutchan, T.M.; Zenk, M.H. Increase of free cysteine and citric acid in plant cells exposed to cobalt ions. Phytochemistry 2002, 60, 467–474. [Google Scholar] [CrossRef]
- Sinclair, S.A.; Krämer, U. The zinc homeostasis network of land plants. Biochim. Et Biophys. Acta Mol. Cell Res. 2012, 1823, 1553–1567. [Google Scholar] [CrossRef]
- Zhang, W.H.; Ryan, P.R.; Tyerman, S.D. Malate permeable channels and cation channels activated by aluminum in the apical cells of wheat roots. Plant Physiol. 2001, 125, 1459–1472. [Google Scholar] [CrossRef] [Green Version]
- Salazar, F.S.; Pandey, S.; Narro, L. Diallel analysis of acid-soil tolerant and intolerant tropical maize populations. Crop Sci. 1997, 37, 1457–1462. [Google Scholar] [CrossRef]
- Ascacio-Valdés, J.A.; Buenrostro-Figueroa, J.J.; Aguilera-Carbo, A.; Prado-Barragán, A.; Rodríguez-Herrera, R.; Aguilar, C.N. Ellagitannins: Biosynthesis, biodegradation and biological properties. J. Med. Plants Res. 2011, 5, 4696–4703. [Google Scholar]
- Khan, A.; Nazar, S.; Lang, I.; Nawaz, H.; Hussain, M.A. Effect of ellagic acid on growth and physiology of canola (Brassica napus L.) under saline conditions. J. Plant Interact. 2017, 12, 520–525. [Google Scholar] [CrossRef] [Green Version]


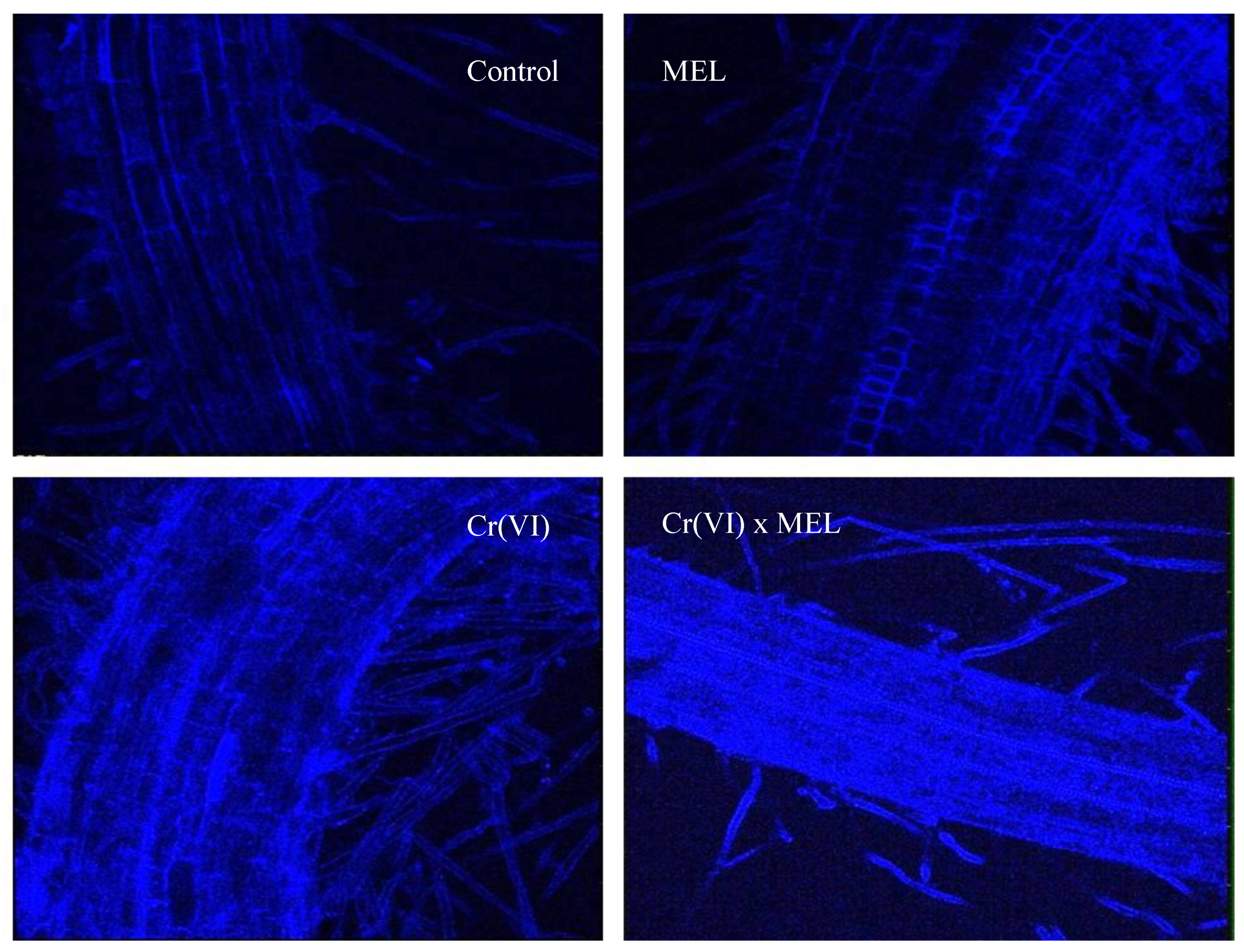{kind=link}
| Sr. No. | Name of the Treatment | MEL (ppm) | Cr(VI) (μM) |
|---|---|---|---|
| 1. | Control | 0 | 0 |
| 2. | MEL-1 | 125 | 0 |
| 3. | MEL-2 | 250 | 0 |
| 4. | MEL-3 | 500 | 0 |
| 5. | Cr(VI) | 0 | 250 |
| 6. | Cr(VI) + MEL-1 | 125 | 250 |
| 7. | Cr(VI) + MEL-2 | 250 | 250 |
| 8. | Cr(VI) + MEL-3 | 500 | 250 |
| Concentrations | Anthocyanin (µg g−1 FW) | Xanthophyll (µg g−1 FW) | |||
| Cr(VI) (µM) | MEL (ppm) | ||||
| 0 | 0 | 1.20 ± 0.58 | 16.10 ± 0.97 | ||
| 0 | 125 | 2.78 ± 1.11 | 18.85 ± 1.32 | ||
| 0 | 250 | 5.39 ± 1.12 | 22.24 ± 0.63 | ||
| 0 | 500 | 11.89 ± 1.37 | 25.21 ± 1.94 | ||
| 250 | 0 | 13.75 ± 0.42 | 28.81 ± 0.97 | ||
| 250 | 125 | 17.47 ± 3.23 | 34.95 ± 2.54 | ||
| 250 | 250 | 24.34 ± 3.20 | 42.37 ± 3.19 | ||
| 250 | 500 | 36.61 ± 3.07 | 48.94 ± 2.77 | ||
| F-ratio Treatment (1,16) | 429.96* | 492.25* | |||
| F-ratio Dose (3,16) | 74.46* | 60.38* | |||
| F-ratio Treatment × Dose (3,16) | 9.86* | 8.55* | |||
| HSD | 5.92 | 5.67 | |||
| Parameter | MLR Equation | β-Regression Coefficient | Multiple Correlation | ||
| Cr(VI) | (MEL) | (Cr(VI) × MEL) | |||
| Anthocyanin | Y = 0.53 + 0.04 Cr(VI) + 0.02 (MEL) + 0.0001 (Cr(VI) × MEL) | 0.54 | 0.36 | 0.38 | 0.99* |
| Xanthophyll | Y = 16.61 + 0.05 Cr(VI) + 0.01 (MEL) + 9 × 10–5 Cr(VI) × MEL) | 0.61 | 0.31 | 0.34 | 0.99* |
| Concentrations | Protein Content (µg/g FW) | SOD (UA mg g−1 Protein) | POD (UA mg g−1 Protein) | ||
| Cr(VI) (µM) | MEL (ppm) | ||||
| 0 | 0 | 18.70 ± 0.45 | 1.37 ± 0.11 | 132.08 ± 2.59 | |
| 0 | 125 | 14.23 ± 0.45 | 1.70 ± 0.47 | 132.35 ± 2.09 | |
| 0 | 250 | 15.90 ± 0.30 | 1.98 ± 1.24 | 133.38 ± 4.68 | |
| 0 | 500 | 16.50 ± 0.90 | 2.09 ± 0.05 | 146.67 ± 1.27 | |
| 250 | 0 | 9.60 ± 0.30 | 6.08 ± 0.66 | 160.58 ± 8.76 | |
| 250 | 125 | 12.60 ± 0.90 | 7.71 ± 0.08 | 184.77 ± 9.37 | |
| 250 | 250 | 15.73 ± 0.45 | 8.51 ± 0.17 | 196.39 ± 1.86 | |
| 250 | 500 | 16.83 ± 0.25 | 9.22 ± 0.09 | 241.91 ± 36.27 | |
| F-ratio Treatment (1,16) | 135.06* | 779.27* | 113.04* | ||
| F-ratio Dose (3,16) | 43.05* | 14.63* | 13.10* | ||
| F-ratio Treatment × Dose (3,16) | 93.06* | 5.56* | 6.05* | ||
| HSD | 1.57 | 1.51 | 38.97 | ||
| Parameter | MLR Equation | β-Regression Coefficient | Multiple Correlation | ||
| Cr(VI) | (MEL) | (Cr(VI) × MEL) | |||
| Protein Content | Y= 16.79 + −0.02 Cr(VI) + −0.002 (MEL) + 7 × 10−6 (Cr(VI) × MEL) | −1.17 | −0.14 | 1.05 | 0.86* |
| SOD | Y= 1.48 + 0.02 Cr(VI) + 0.001 (MEL) +2 × 10−6 (Cr(VI) × MEL) | 0.80 | 0.08 | 0.24 | 0.99* |
| POD | Y= 129.53 + 0.12 Cr(VI) + 0.03 (MEL) + 0.0005 (Cr(VI) × MEL) | 0.43 | 0.15 | 0.59 | 0.99* |
| Concentrations | CAT (UA mg g−1 Protein) | APOX (UA mg g−1 Protein) | GR (UA mg g−1 Protein) | ||
| Cr(VI) (µM) | MEL (ppm) | ||||
| 0 | 0 | 7.27 ± 1.19 | 14.28 ± 1.78 | 27.01 ± 3.62 | |
| 0 | 125 | 7.56 ± 0.65 | 14.76 ± 4.32 | 25.96 ± 3.10 | |
| 0 | 250 | 7.82 ± 1.86 | 15.02 ± 3.47 | 28.20 ± 2.26 | |
| 0 | 500 | 8.40 ± 1.50 | 16.61 ± 2.82 | 28.44 ± 1.78 | |
| 250 | 0 | 16.40 ±1.92 | 41.66 ± 6.27 | 47.16 ± 3.71 | |
| 250 | 125 | 17.54 ± 2.19 | 53.04 ± 2.61 | 57.16 ± 1.24 | |
| 250 | 250 | 18.58 ± 2.65 | 61.30 ± 5.74 | 61.44 ± 3.56 | |
| 250 | 500 | 23.22 ± 2.06 | 80.94 ± 2.72 | 74.78 ± 3.92 | |
| F-ratio Treatment (1,16) | 217.87* | 725.75* | 689.60* | ||
| F-ratio Dose (3,16) | 5.24* | 28.78* | 24.07* | ||
| F-ratio Treatment × Dose (3,16) | 2.76 | 22.64* | 18.55* | ||
| HSD | 5.24 | 11.33 | 8.63 | ||
| Parameter | MLR Equation | β-Regression Coefficient | Multiple Correlation | ||
| Cr(VI) | (MEL) | (Cr(VI) × MEL) | |||
| CAT | Y= 7.27 + 0.03 Cr(VI) + 0.002 (MEL) + 5 × 10−6 (Cr(VI) × MEL) | 0.73 | 0.07 | 0.33 | 0.99* |
| APOX | Y= 14.15 + 0.11 Cr(VI) + 0.004 (MEL) + 0.0003 (Cr(VI) × MEL) | 0.57 | 0.03 | 0.51 | 0.99* |
| GR | Y= 26.54 + 0.08 Cr(VI) + 0.003 (MEL) + 0.0002 (Cr(VI) × MEL) | 0.61 | 0.04 | 0.47 | 0.99* |
| Concentrations | DHAR (UA mg g−1 Protein) | PPO (UA mg g−1 Protein) | GST (UA mg g−1 Protein) | GPOX (UA mg g−1 Protein) | ||
| Cr(VI) (µM) | MEL (ppm) | |||||
| 0 | 0 | 26.95 ± 2.57 | 2.54 ± 0.42 | 16.80 ± 1.47 | 16.41 ± 0.34 | |
| 0 | 125 | 25.71 ± 2.85 | 2.75 ± 0.27 | 16.91 ± 0.18 | 17.75 ± 0.39 | |
| 0 | 250 | 27.80 ± 2.65 | 3.13 ± 0.13 | 17.90 ± 0.82 | 17.82 ± 0.13 | |
| 0 | 500 | 30.57 ± 0.75 | 3.74 ± 0.37 | 18.56 ± 0.29 | 17.98 ± 0.34 | |
| 250 | 0 | 54.28 ± 2.85 | 8.96 ± 0.86 | 34.64 ± 4.46 | 26.09 ± 0.67 | |
| 250 | 125 | 69.62 ± 1.08 | 10.83 ± 0.43 | 45.82 ± 2.23 | 27.65 ± 0.79 | |
| 250 | 250 | 78.92 ± 4.32 | 12.47 ± 0.43 | 56.23 ± 3.94 | 26.49 ± 0.21 | |
| 250 | 500 | 92.65 ± 2.67 | 13.58 ± 0.37 | 64.51 ± 1.65 | 28.67 ± 1.05 | |
| F-ratio Treatment (1,16) | 1770.58* | 2035.90* | 1115.51* | 1708.56* | ||
| F-ratio Dose (3,16) | 65.78* | 45.55* | 49.10* | 13.93* | ||
| F-ratio Treatment × Dose (3,16) | 44.24* | 16.64* | 38.32* | 3.09* | ||
| HSD | 7.59 | 1.29 | 6.79 | 1.63 | ||
| Parameter | MLR Equation | β-Regression Coefficient | Multiple Correlation | |||
| Cr(VI) | (MEL) | (Cr(VI) x MEL) | ||||
| DHAR | Y= 25.9 + 0.12 Cr(VI) + 0.008 (MEL) + 0.0003 (Cr(VI) × MEL) | 0.63 | 0.06 | 0.44 | 0.99* | |
| PPO | Y= 2.5 + 0.02Cr(VI) + 0.002 (MEL) +3 × 10−6 Cr(VI) × MEL) | 0.79 | 0.10 | 0.25 | 0.99* | |
| GST | Y= 16.71 + 0.08 Cr(VI) + 0.003 (MEL) + 0.0002 (Cr(VI) × MEL) | 0.56 | 0.03 | 0.51 | 0.99* | |
| GPOX | Y= 16.92 + 0.03 Cr(VI) + 0.002 (MEL) + 7 × 10−7 (Cr(VI) × MEL) | 0.94 | 0.09 | 0.05 | 0.99* | |
| Concentrations | Ascorbic Acid (µg g−1 FW) | Tocopherol (µg g−1 FW) | Glutathione (µg g−1 FW) | |||
| Cr(VI) (µM) | MEL (ppm) | |||||
| 0 | 0 | 11.97 ± 0.36 | 21.88 ± 1.90 | 299.79 ± 7.68 | ||
| 0 | 125 | 15.80 ± 0.72 | 30.42 ± 1.02 | 333.10 ± 8.87 | ||
| 0 | 250 | 19.70 ± 1.88 | 32.22 ± 0.62 | 384.35 ± 20.33 | ||
| 0 | 500 | 26.45 ± 0.49 | 40.76 ± 4.12 | 443.28 ± 16.01 | ||
| 250 | 0 | 22.60 ± 0.41 | 47.88 ± 0.69 | 481.71 ± 11.74 | ||
| 250 | 125 | 28.45 ± 1.27 | 62.52 ± 0.04 | 407.41 ± 20.33 | ||
| 250 | 250 | 39.30 ± 0.65 | 82.07 ± 4.85 | 520.15 ± 29.10 | ||
| 250 | 500 | 49.05 ± 2.23 | 98.99 ± 7.33 | 635.45 ± 19.34 | ||
| F-ratio Treatment (1,16) | 1097.11* | 828.52* | 396.82* | |||
| F-ratio Dose (3,16) | 326.84* | 107.15* | 107.27* | |||
| F-ratio Treatment × Dose (3,16) | 32.78* | 27.12* | 13.43* | |||
| HSD | 3.42 | 10.001 | 50.8 | |||
| Parameter | MLR Equation | β-Regression Coefficient | Multiple Correlation | |||
| Cr(VI) | (MEL) | (Cr(VI) × MEL) | ||||
| Ascorbic Acid | Y= 12.15 + 0.04 Cr(VI) + 0.02 (MEL) + 0.0001 (Cr(VI) × MEL) | 0.47 | 0.46 | 0.37 | 0.99* | |
| Tocopherol | Y= 23.59 + 0.1 Cr(VI) + 0.03 (MEL) + 0.0003 (Cr(VI) × MEL) | 0.52 | 0.25 | 0.45 | 0.99* | |
| Glutathione | Y= 301.33 + 0.51 Cr(VI) + 0.29 (MEL) + 0.0003 (Cr(VI) × MEL) | 0.63 | 0.53 | 0.14 | 0.94* | |
| Concentrations | Gallic Acid (µg g−1 FW) | ChlorogenicAcid (µg g−1 FW) | Caffeic Acid (µg g−1 FW) | Catechin (µg g−1 FW) | Coumaric Acid (µg g−1 FW) | ||
| Cr(VI) (µM) | MEL (ppm) | ||||||
| 0 | 0 | 9.60 ± 1.40 | 1.74 ± 0.05 | 0.10 ± 0.002 | 263.80 ± 16.15 | 0.18 ± 0.004 | |
| 0 | 125 | 9.87 ± 0.60 | 1.64 ± 0.01 | 0.15 ± 0.008 | 268.03 ± 49.93 | 0.16 ± 0.006 | |
| 0 | 250 | 10.46 ± 1.06 | 1.74 ± 0.03 | 0.16±0.003 | 282.89 ± 55.04 | 0.18 ± 0.006 | |
| 0 | 500 | 13.57 ± 0.91 | 1.81 ± 0.03 | 0.18 ± 0.004 | 287.84 ± 9.55 | 0.21 ± 0.006 | |
| 250 | 0 | 16.05 ± 0.43 | 4.60 ± 0.11 | 0.94 ± 0.009 | 331.08 ± 53.67 | 0.44 ± 0.020 | |
| 250 | 125 | 13.22 ± 0.71 | 3.21 ± 0.64 | 0.84 ± 0.01 | 422.96 ± 10.10 | 0.35 ± 0.010 | |
| 250 | 250 | 15.18 ± 0.61 | 4.22 ± 0.25 | 0.96 ± 0.04 | 430.51 ± 20.83 | 0.59 ± 0.020 | |
| 250 | 500 | 17.51 ± 0.50 | 5.26 ± 0.12 | 1.24 ± 0.16 | 447.50 ± 35.40 | 0.74 ± 0.050 | |
| F-ratio Treatment (1,16) | 185.09* | 618.21* | 124.54* | 79.75* | 1207.79* | ||
| F-ratio Dose (3,16) | 24.66* | 19.73* | 16.50* | 4.35* | 87.41* | ||
| F-ratio Treatment × Dose (3,16) | 3.93* | 14.41* | 10.08* | 2.17 | 59.07* | ||
| HSD | 2.35 | 0.72 | 0.16 | 102.71 | 0.06 | ||
| Parameter | MLR Equation | β-Regression Coefficient | Multiple Correlation | ||||
| Cr(VI) | (MEL) | (Cr(VI) × MEL) | |||||
| Gallic Acid | Y= 9.08 + 0.02 Cr(VI) + 0.008 (MEL) + 1 × 10−6 (Cr(VI) × MEL) | 0.96 | 0.54 | −0.2 | 0.93* | ||
| Chlorogenic Acid | Y= 1.69 + 0.008 Cr(VI) + 0.0002 (MEL) + 8 × 10−7 (Cr(VI) × MEL) | 0.76 | 0.02 | 0.24 | 0.94* | ||
| Caffeic Acid | Y= 0.12 + 0.002 Cr(VI) + 0.0001 (MEL) + 2× 10−7 (Cr(VI) × MEL) | 0.83 | 0.06 | 0.21 | 0.99* | ||
| Catechin | Y= 264.5 + 0.39 Cr(VI) + 0.05 (MEL) + 0.0006 (Cr(VI) × MEL) | 0.67 | 0.12 | 0.34 | 0.96* | ||
| Coumaric Acid | Y= 0.17 + 0.0008 Cr(VI) + 7 × 10−6 (MEL) + 3 × 10−7 (Cr(VI) × MEL) | 0.51 | 0.05 | 0.53 | 0.96* | ||
| Concentrations | Aspartic Acid (µg g−1 FW) | Glutamine (µg g−1 FW) | β-Alanine (µg g−1 FW) | Lysine (µg g−1 FW) | ||
| Cr(VI) (µM) | MEL (ppm) | |||||
| 0 | 0 | 79.00 ± 0.85 | 40.61 ± 0.35 | 168.83 ± 0.86 | 35.11 ± 0.48 | |
| 0 | 125 | 45.71 ± 2.43 | 37.30 ± 0.46 | 90.79 ± 0.34 | 17.81 ± 0.62 | |
| 0 | 250 | 63.18 ± 0.39 | 38.45 ± 0.50 | 101.34 ± 0.51 | 18.85 ± 0.85 | |
| 0 | 500 | 74.18 ± 0.68 | 39.72 ± 0.98 | 114.37 ± 0.48 | 20.73 ± 0.81 | |
| 250 | 0 | 32.16 ± 0.60 | 8.82 ± 1.12 | 35.96 ± 0.98 | 18.54 ± 1.44 | |
| 250 | 125 | 73.39 ± 0.57 | 30.08 ± 0.71 | 115.62 ± 0.22 | 23.70 ± 1.03 | |
| 250 | 250 | 75.90 ± 0.13 | 36.40 ± 0.51 | 124.54 ± 1.40 | 29.54 ± 0.78 | |
| 250 | 500 | 78.04 ± 0.51 | 39.60 ± 0.57 | 164.09 ± 0.67 | 30.84 ± 1.14 | |
| F-ratio Treatment (1,16) | 2.43 | 1295.79* | 769.44* | 43.96* | ||
| F-ratio Dose (3,16) | 509.11* | 529.81* | 2956.89* | 48.24* | ||
| F-ratio Treatment × Dose (3,16) | 1520.74* | 655.19* | 17456.18* | 285.57* | ||
| HSD | 2.87 | 1.98 | 2.19 | 2.64 | ||
| Parameter | MLR equation | β-Regression Coefficient | Multiple correlation | |||
| Cr(VI) | (MEL) | (Cr(VI) × MEL) | ||||
| Aspartic Acid | Y= 63.48 + −0.06 Cr(VI) + 0.009 (MEL) + 0.0003 (Cr(VI) × MEL) | −0.17 | 0.1 | 0.7 | 0.62 | |
| Glutamine | Y= 39.03 + −0.08 Cr(VI) + 5 × 10−6 (MEL) +0.0002 (Cr(VI) × MEL) | −1.12 | −0.0009 | 0.94 | 0.88* | |
| β-Alanine | Y= 135.01 + −0.3 Cr(VI) + −0.074 (MEL) + 0.0012 (Cr(VI) × MEL) | −0.95 | −0.34 | 1.31 | 0.8* | |
| Lysine | Y= 27.81 + −0.03 Cr(VI) + −0.02 (MEL) + 0.0002 (Cr(VI) × MEL) | −0.6 | −0.64 | 1.26 | 0.71* | |
| Concentrations | Glutamic Acid (µg g−1 FW) | Asparagine (µg g−1 FW) | Serine (µg g−1 FW) | Isoleucine (µg g−1 FW) | ||
| Cr(VI) (µM) | MEL (ppm) | |||||
| 0 | 0 | 29.83 ± 0.76 | 1134.65 ± 48.39 | 10.01 ± 0.38 | 174.95 ± 1.75 | |
| 0 | 125 | 15.84 ± 0.79 | 443.42 ± 9.80 | 1.23 ± 0.09 | 65.96 ± 0.77 | |
| 0 | 250 | 18.80 ± 0.87 | 652.13 ± 14.26 | 1.94 ± 0.13 | 68.31 ± 0.60 | |
| 0 | 500 | 22.84 ± 0.70 | 686.02 ± 12.23 | 3.68 ± 0.56 | 86.11 ± 1.10 | |
| 250 | 0 | 8.65 ± 1.31 | 355.18 ± 34.14 | 0.58 ± 0.15 | 13.75 ± 3.03 | |
| 250 | 125 | 25.35 ± 0.71 | 824.64 ± 26.16 | 6.15 ± 0.06 | 87.03 ± 1.25 | |
| 250 | 250 | 27.43 ± 0.52 | 875.57 ± 16.85 | 6.98 ± 0.12 | 104.83 ± 0.99 | |
| 250 | 500 | 28.47 ± 0.50 | 974.98 ± 22.29 | 7.84 ± 0.89 | 146.25 ± 16.09 | |
| F-ratio Treatment (1,16) | 3.83 | 7.20* | 49.47* | 20.52* | ||
| F-ratio Dose (3,16) | 73.33* | 58.91* | 30.23* | 49.36* | ||
| F-ratio Treatment × Dose (3,16) | 489.49* | 651.33* | 451.69* | 447.64* | ||
| HSD | 2.29 | 73.65 | 1.153 | 16.62 | ||
| Parameter | MLR equation | β-Regression Coefficient | Multiple correlation | |||
| Cr(VI) | (MEL) | (Cr(VI) × MEL) | ||||
| Glutamic Acid | Y= 23.43 + −0.03 Cr(VI) + −0.007 (MEL) + 0.0002 (Cr(VI) × MEL) | −0.61 | −0.19 | 1.02 | 0.66 | |
| Asparagine | Y= 851.39 + −1.31 Cr(VI) + −0.55 (MEL) + 0.0065 (Cr(VI) × MEL) | −0.66 | −0.42 | 1.13 | 0.64 | |
| Serine | Y= 6.15 + −0.01 Cr(VI) + −0.009 (MEL) + 9 × 10−6 (Cr(VI) × MEL) | −0.54 | −0.5 | 1.13 | 0.64 | |
| Isoleucine | Y= 127.8 + −0.37 Cr(VI) + −0.13 (MEL) + 0.001 (Cr(VI) × MEL) | −0.99 | −0.52 | 1.37 | 0.78* | |
| Concentrations | Glycine (µg g−1 FW) | Threonine (µg g−1 FW) | Citrulline (µg g−1 FW) | Arginine (µg g−1 FW) | GABA (µg g−1 FW) | ||
| Cr(VI)(µM) | MEL (ppm) | ||||||
| 0 | 0 | 9.47 ± 0.56 | 6.39 ± 0.68 | 22.55 ± 1.18 | 647.02 ± 12.48 | 5.009 ± 0.26 | |
| 0 | 125 | 4.64 ± 0.40 | 2.72 ± 0.22 | 10.97 ± 0.81 | 97.11 ± 5.87 | 1.89 ± 0.04 | |
| 0 | 250 | 5.48 ± 0.46 | 3.38 ± 0.72 | 12.60 ± 0.50 | 164.57 ± 6.72 | 2.47 ± 0.40 | |
| 0 | 500 | 6.07 ± 0.76 | 3.81 ± 0.06 | 15.61 ± 1.07 | 181.85 ± 6.55 | 3.06 ± 0.67 | |
| 250 | 0 | 3.58 ± 0.32 | 1.37 ± 0.42 | 4.91 ± 0.75 | 78.43 ± 7.27 | 1.18 ± 0.37 | |
| 250 | 125 | 6.77 ± 0.37 | 3.72 ± 0.09 | 15.93 ± 0.82 | 311.96 ± 49.42 | 2.89 ± 0.12 | |
| 250 | 250 | 7.25 ± 0.29 | 4.22 ± 0.27 | 19.47 ± 0.62 | 466.28 ± 18.01 | 3.49 ± 0.43 | |
| 250 | 500 | 8.14 ± 0.96 | 4.86 ± 0.76 | 20.94 ± 0.52 | 575.68 ± 11.60 | 4.16 ± 0.83 | |
| F-ratio Treatment (1,16) | 0.006 | 7.06* | 0.12 | 108.4* | 0.84 | ||
| F-ratio Dose (3,16) | 6.24* | 5.26* | 45.23* | 91.77* | 6.97* | ||
| F-ratio Treatment × Dose (3,16) | 73.37* | 56.22* | 305.2* | 725.5* | 41.19* | ||
| HSD | 1.59 | 1.38 | 2.32 | 56.87 | 1.31 | ||
| Parameter | MLR Equation | β-Regression Coefficient | Multiple Correlation | ||||
| Cr(VI) | (MEL) | (Cr(VI) × MEL) | |||||
| Glycine | Y= 7.42 + −0.01 Cr(VI) + −0.005 (MEL) + 5 × 10−6 (Cr(VI) × MEL) | −0.77 | −0.47 | 1.2 | 0.67 | ||
| Threonine | Y= 4.84 + −0.01 Cr(VI) + −0.003 (MEL) + 4 × 10−6 (Cr(VI) × MEL) | −0.96 | −0.46 | 1.19 | 0.7 | ||
| Citrulline | Y= 17.32 + −0.03 Cr(VI) + −0.009 (MEL) + 0.0001 (Cr(VI) × MEL) | −0.76 | −0.29 | 1.17 | 0.72* | ||
| Arginine | Y= 423.61 + −1.09 Cr(VI) + −0.69 (MEL) + 0.006 (Cr(VI) × MEL) | −0.65 | −0.61 | 1.34 | 0.76* | ||
| GABA | Y= 3.64 + −0.008 Cr(VI) + −0.002 (MEL) + 3 × 10−6 (Cr(VI) × MEL) | −0.83 | −0.39 | 1.18 | 0.69 | ||
| Concentrations | Cystine (µg g−1 FW) | Ornithine (µg g−1 FW) | Proline (µg g−1 FW) | Methionine (µg g−1 FW) | Total Amino Acids (µg g−1 FW) | ||
| Cr(VI) (µM) | MEL (ppm) | ||||||
| 0 | 0 | 28.42 ± 2.48 | 9.91 ± 1.03 | 44.49 ± 9.005 | 51.98 ± 0.79 | 2504.69 ± 54.9 | |
| 0 | 125 | 8.56 ± 2.02 | 5.82 ± 0.92 | 14.39 ± 2.14 | 26.21 ± 0.55 | 893.15 ± 24.40 | |
| 0 | 250 | 11.94 ± 1.48 | 6.58 ± 0.44 | 16.94 ± 0.90 | 33.15 ± 1.47 | 1223.58 ± 13.29 | |
| 0 | 500 | 14.78 ± 0.85 | 7.38 ± 0.51 | 19.38 ± 0.59 | 43.61 ± 0.67 | 1347.07 ± 14.95 | |
| 250 | 0 | 4.54 ± 0.54 | 3.30 ± 0.47 | 9.99 ± 0.92 | 9.10 ± 0.68 | 591.47 ± 47.18 | |
| 250 | 125 | 19.49 ± 0.64 | 7.47 ± 0.40 | 22.49 ± 1.65 | 45.85 ± 1.04 | 1626.33 ± 73.72 | |
| 250 | 250 | 23.03 ± 0.43 | 8.42 ± 0.19 | 26.66 ± 2.99 | 47.92 ± 1.009 | 1892.25 ± 29.66 | |
| 250 | 500 | 27.77 ± 1.03 | 9.59 ± 1.17 | 32.17 ± 3.34 | 50.54 ± 0.52 | 2208.89 ± 49.85 | |
| F-ratio Treatment (1,16) | 24.46* | 0.59 | 0.41 | 1.08 | 24.5* | ||
| F-ratio Dose (3,16) | 28.71* | 9.01* | 6.87* | 366.005* | 144.16* | ||
| F-ratio Treatment × Dose (3,16) | 250.58* | 52.11* | 54.57* | 1549.31* | 1424.96* | ||
| HSD | 3.89 | 2.04 | 10.52 | 2.53 | 122.65 | ||
| Parameter | MLR Equation | β-Regression Coefficient | Multiple correlation | ||||
| Cr(VI) | (MEL) | (Cr(VI) × MEL) | |||||
| Cystine | Y= 19.91 + −0.04 Cr(VI) + −0.01 (MEL) + 0.0002 (Cr(VI) × MEL) | −0.62 | −0.4 | 1.23 | 0.74* | ||
| Ornithine | Y= 8.11 + −0.01 Cr(VI) + −0.003 (MEL) + 6 × 10−6 (Cr(VI) × MEL) | −0.84 | −0.29 | 1.22 | 0.76* | ||
| Proline | Y= 31.96 + −0.07 Cr(VI) + −0.03 (MEL) + 0.0003 (Cr(VI) × MEL) | −0.87 | −0.66 | 1.29 | 0.7* | ||
| Methionine | Y= 39.58 + −0.06 Cr(VI) + −0.004 (MEL) + 0.0003 (Cr(VI) × MEL) | −0.58 | −0.05 | 0.89 | 0.65 | ||
| Total Amino Acids | Y= 1835.4 + −3.57 Cr(VI) + −1.56 (MEL) + 0.01 (Cr(VI) × MEL) | −0.73 | −0.47 | 1.25 | 0.71* | ||
| Concentrations | FumaricAcid Content (µg g−1 DW) | Malic Acid Content (µg g−1 DW) | Citric Acid Content (µg g−1 DW) | Succinic Acid Content (µg g−1 DW) | ||
| Cr(VI)(µM) | MEL (ppm) | |||||
| 0 | 0 | 0.38 ± 0.009 | 1.47 ± 0.15 | 2.32 ± 0.02 | 0.870 ± 0.05 | |
| 0 | 125 | 0.393 ± 0.005 | 1.45 ± 0.41 | 2.32 ± 0.15 | 0.783 ± 0.01 | |
| 0 | 250 | 0.395 ± 0.005 | 1.46 ± 0.19 | 2.40 ± 0.01 | 0.824 ± 0.01 | |
| 0 | 500 | 0.410 ± 0.008 | 1.47 ± 0.11 | 2.40 ± 0.01 | 0.866 ± 0.08 | |
| 250 | 0 | 0.415 ± 0.010 | 2.15 ± 0.21 | 2.94 ± 0.34 | 0.876 ± 0.03 | |
| 250 | 125 | 0.394 ± 0.008 | 2.19 ± 0.23 | 3.64 ± 0.10 | 0.867 ± 0.04 | |
| 250 | 250 | 0.409 ± 0.007 | 2.48 ± 0.02 | 3.67 ± 0.05 | 0.884 ± 0.04 | |
| 250 | 500 | 0.416 ± 0.008 | 2.50 ± 0.15 | 3.68 ± 0.08 | 0.899 ± 0.01 | |
| F-ratio Treatment (1,16) | 10.75* | 95.55* | 370.78* | 11.30* | ||
| F-ratio Dose (3,16) | 1.97 | 1.11 | 11.18* | 1.07 | ||
| F-ratio Treatment × Dose (3,16) | 10.73* | 1.05 | 8.20* | 3.71* | ||
| HSD | 0.02 | 0.61 | 0.4 | 0.09 | ||
| Parameter | MLR Equation | β-Regression Coefficient | Multiple Correlation | |||
| Cr(VI) | (MEL) | (Cr(VI) × MEL) | ||||
| Fumaric acid | Y= 0.38 + 0.0001 Cr(VI) + 5 × 10−6 (MEL) + 3 × 10−8 (Cr(VI) × MEL) | 1.21 | 0.78 | −1.16 | 0.8* | |
| Malic acid | Y= 1.46 + 0.002 Cr(VI) + 1 × 10−6 0.000001 (MEL) + 3 × 10−7 (Cr(VI) × MEL) | 0.78 | 0.006 | 0.28 | 0.99* | |
| Citric acid | Y= 2.23 + 0.003 Cr(VI) + 0.0002 (MEL) + 4 × 10−7 (Cr(VI) × MEL) | 0.74 | 0.05 | 0.28 | 0.96* | |
| Succinic acid | Y= 0.82 + 0.0002 Cr(VI) + 4 × 10−6 0.000004 (MEL) + 3 × 10−8 (Cr(VI) × MEL) | 0.88 | 0.21 | −0.34 | 0.68 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gautam, V.; Sharma, P.; Bakshi, P.; Arora, S.; Bhardwaj, R.; Paray, B.A.; Alyemeni, M.N.; Ahmad, P. Effect of Rhododendron arboreum Leaf Extract on the Antioxidant Defense System against Chromium (VI) Stress in Vigna radiata Plants. Plants 2020, 9, 164. https://doi.org/10.3390/plants9020164
Gautam V, Sharma P, Bakshi P, Arora S, Bhardwaj R, Paray BA, Alyemeni MN, Ahmad P. Effect of Rhododendron arboreum Leaf Extract on the Antioxidant Defense System against Chromium (VI) Stress in Vigna radiata Plants. Plants. 2020; 9(2):164. https://doi.org/10.3390/plants9020164
Chicago/Turabian StyleGautam, Vandana, Pooja Sharma, Palak Bakshi, Saroj Arora, Renu Bhardwaj, Bilal Ahmad Paray, Mohammed Nasser Alyemeni, and Parvaiz Ahmad. 2020. "Effect of Rhododendron arboreum Leaf Extract on the Antioxidant Defense System against Chromium (VI) Stress in Vigna radiata Plants" Plants 9, no. 2: 164. https://doi.org/10.3390/plants9020164
APA StyleGautam, V., Sharma, P., Bakshi, P., Arora, S., Bhardwaj, R., Paray, B. A., Alyemeni, M. N., & Ahmad, P. (2020). Effect of Rhododendron arboreum Leaf Extract on the Antioxidant Defense System against Chromium (VI) Stress in Vigna radiata Plants. Plants, 9(2), 164. https://doi.org/10.3390/plants9020164