Physiological, Biochemical, and Metabolic Responses to Short and Prolonged Saline Stress in Two Cultivated Cardoon Genotypes

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Morphological Traits
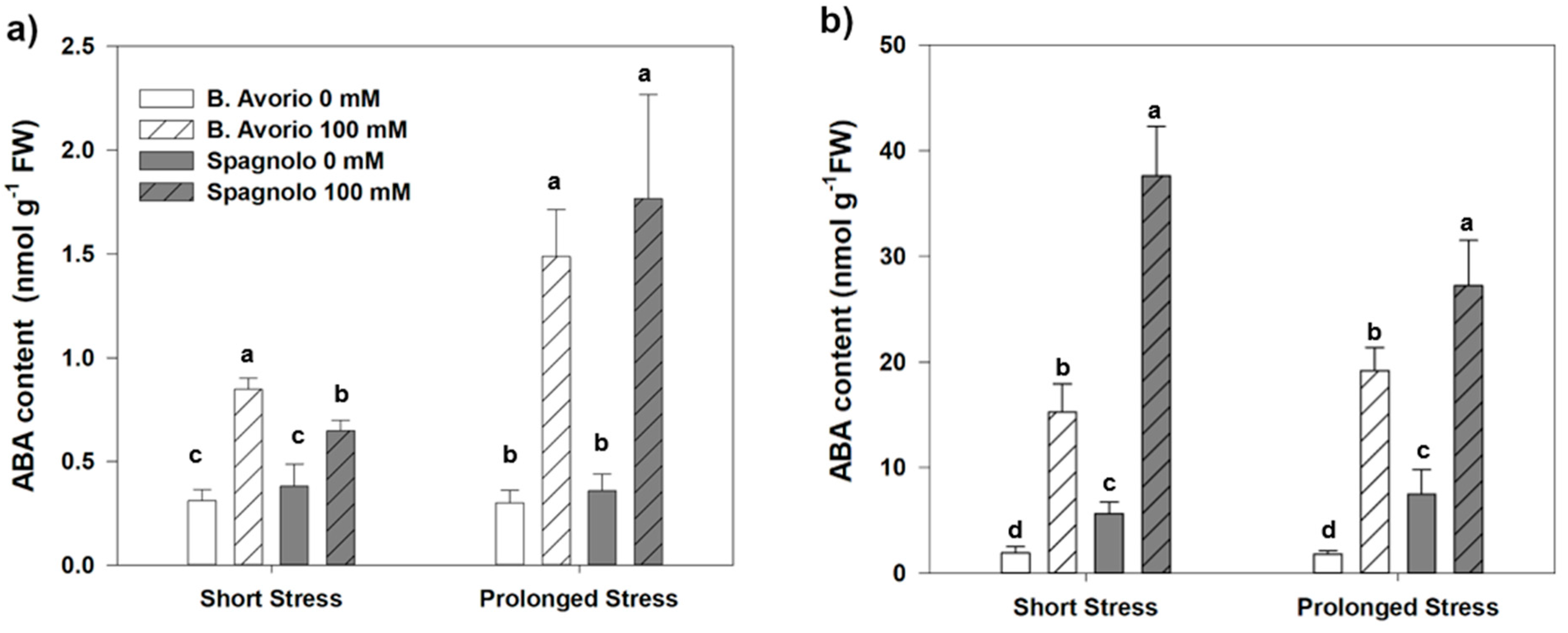
2.2. Effect of Salt Stress on ABA Concentration
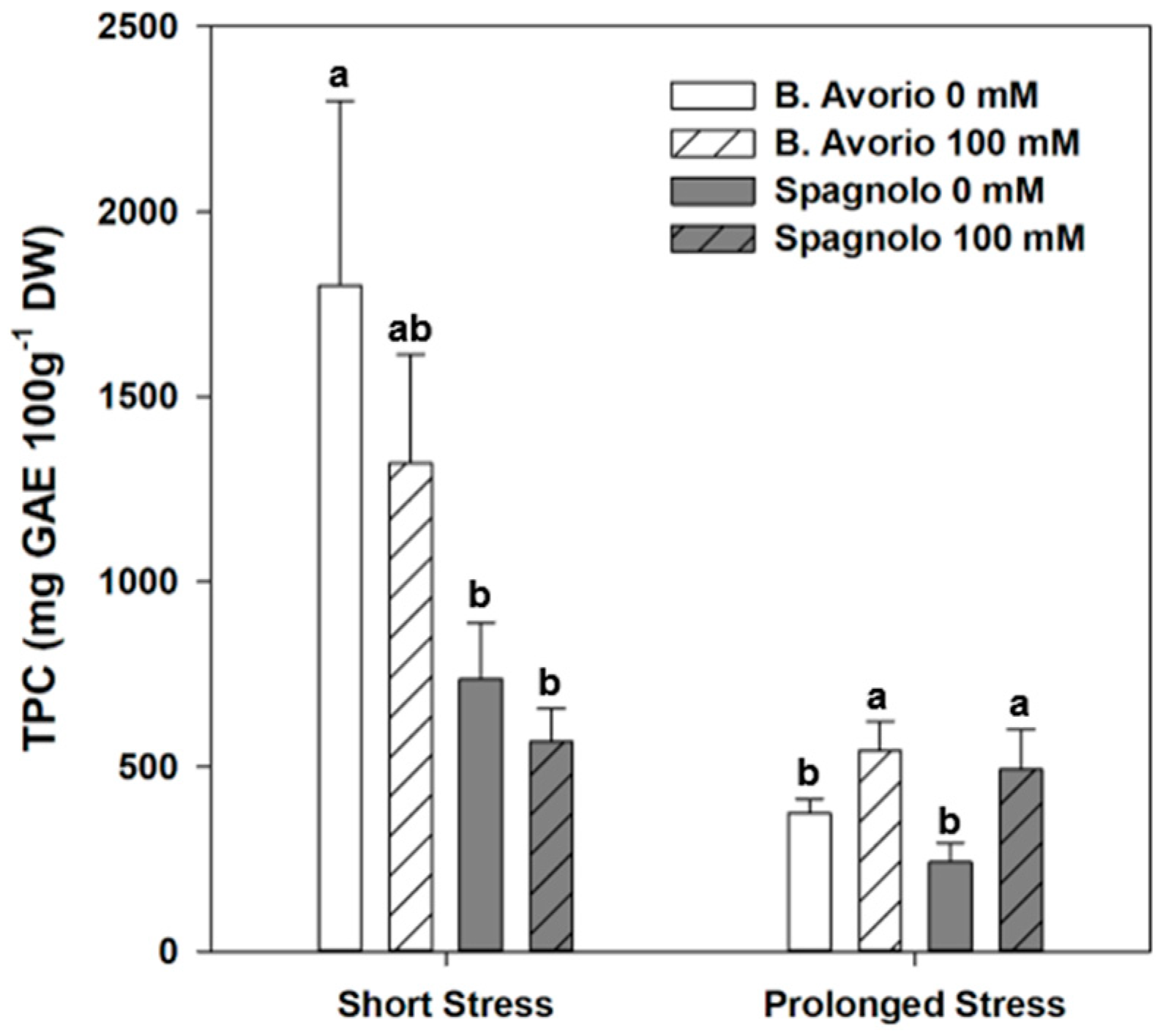
2.3. Metabolites Production under Salt Stress
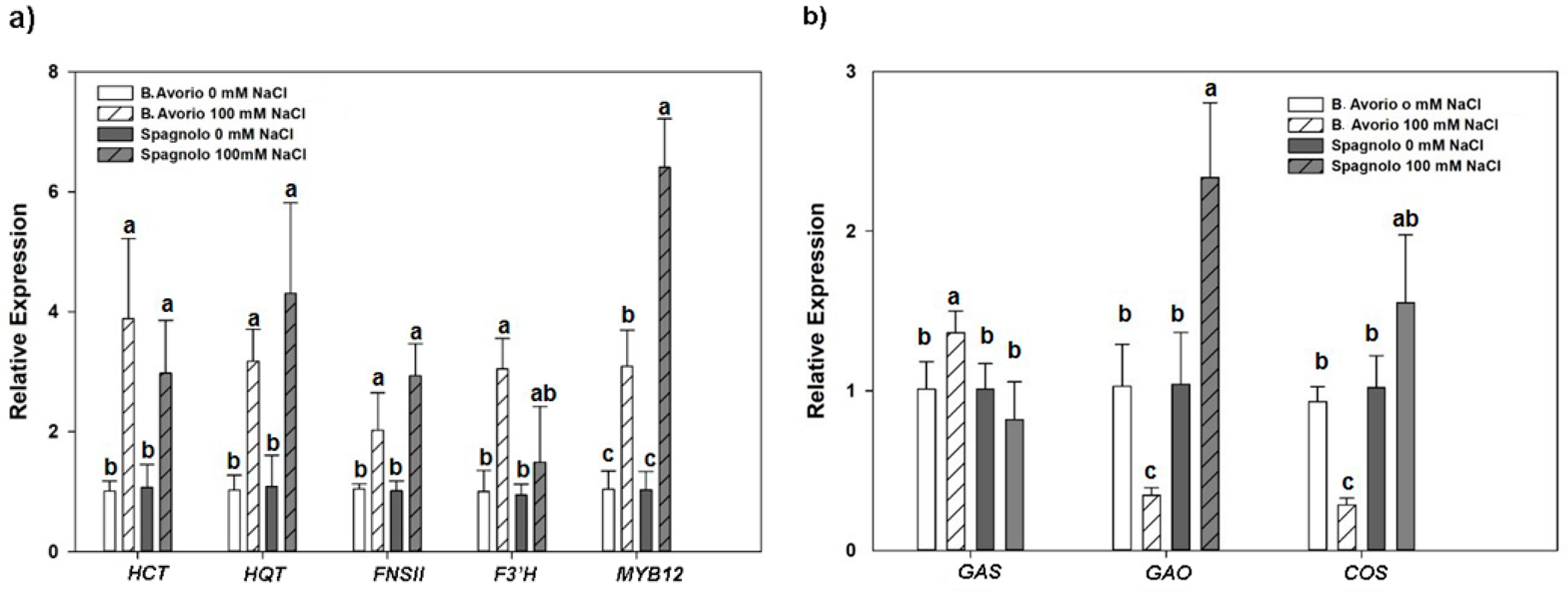
2.4. Transcriptional Changes in Phenylpropanoids and Sesquiterpenes Biosynthetic Genes
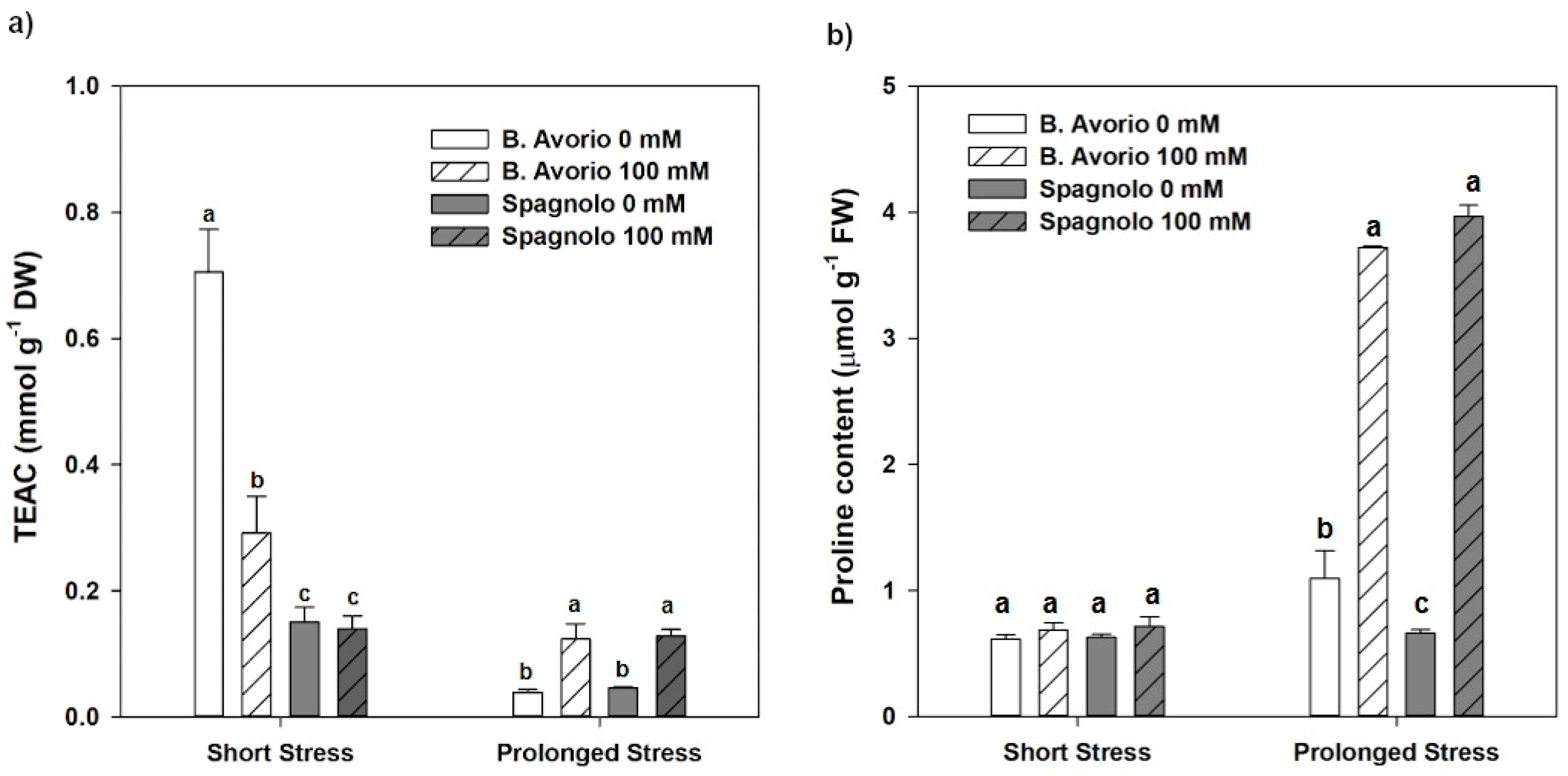
2.5. Physiological Traits: Non-Enzymatic Antioxidant Machinery
2.6. Enzymatic Antioxidant Machinery
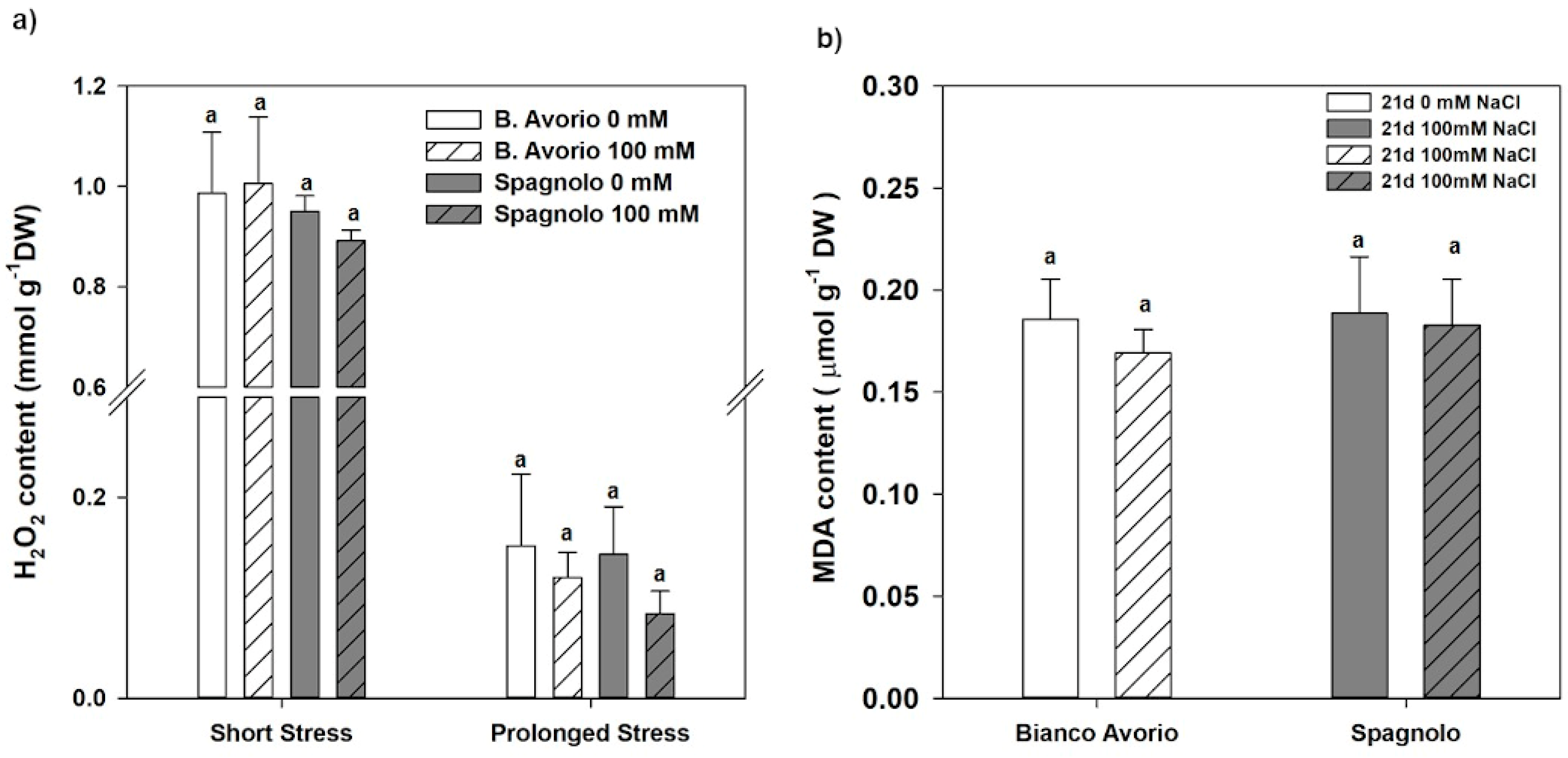
2.7. Oxidative Stress Detection
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Growth Parameters
4.3. ABA Determination
4.4. Analysis of Secondary Metabolites
4.4.1. Extraction
4.4.2. Determination of Total Phenolics Content (TPC)
4.4.3. Secondary Metabolite Profiling by UHPLC-DAD-HRMS
4.4.4. Quantitative Analysis of Target Polyphenols and Cynaropicrin
4.5. RNA Extraction and qRT-PCR
4.6. Non-Enzymatic Antioxidants: ABTS and Proline
4.7. Antioxidant Enzymes
4.8. MDA and H2O2 Assays
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.J.; Drechsel, P.; Noble, A.D.D. Economics of salt-induced land degradation and restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, A.; Takeda, S.; Nick, P. Life and death under salt stress: Same players, different timing? J. Exp. Bot. 2014, 65, 2963–2979. [Google Scholar] [CrossRef] [PubMed]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.-K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High Salinity Induces Different Oxidative Stress and Antioxidant Responses in Maize Seedlings Organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef] [Green Version]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Fini, A.; Guidi, L.; Ferrini, F.; Brunetti, C.; Di Ferdinando, M.; Biricolti, S.; Pollastri, S.; Calamai, L.; Tattini, M. Drought stress has contrasting effects on antioxidant enzymes activity and phenylpropanoid biosynthesis in Fraxinus ornus leaves: An excess light stress affair? J. Plant Physiol. 2012, 169, 929–939. [Google Scholar] [CrossRef]
- Tattini, M.; Loreto, F.; Fini, A.; Guidi, L.; Brunetti, C.; Velikova, V.; Gori, A.; Ferrini, F. Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed Platanus × acerifolia plants during Mediterranean summers. New Phytol. 2015, 207, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Tattini, M. Multiple functional roles of flavonoids in photoprotection. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, C.; Guidi, L.; Sebastiani, F.; Tattini, M. Isoprenoids and phenylpropanoids are key components of the antioxidant defense system of plants facing severe excess light stress. Environ. Exp. Bot. 2015, 119, 54–62. [Google Scholar] [CrossRef]
- Arena, C.; Figlioli, F.; Sorrentino, M.C.; Izzo, L.G.; Capozzi, F.; Giordano, S.; Spagnuolo, V. Ultrastructural, protein and photosynthetic alterations induced by Pb and Cd in Cynara cardunculus L., and its potential for phytoremediation. Ecotoxicol. Environ. Saf. 2017, 145, 83–89. [Google Scholar] [CrossRef]
- Sorrentino, M.C.; Capozzi, F.; Amitrano, C.; Giordano, S.; Arena, C.; Spagnuolo, V. Performance of three cardoon cultivars in an industrial heavy metal-contaminated soil: Effects on morphology, cytology and photosynthesis. J. Hazard. Mater. 2018, 351, 131–137. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Svecova, E.; Rea, E.; Lucini, L. Effects of saline stress on mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon genotypes grown in floating system. J. Sci. Food Agric. 2013, 93, 1119–1127. [Google Scholar] [CrossRef]
- Papazoglou, E.G. Responses of Cynara cardunculus L. to single and combined cadmium and nickel treatment conditions. Ecotoxicol. Environ. Saf. 2011, 74, 195–202. [Google Scholar] [CrossRef]
- Pappalardo, H.D.; Toscano, V.; Puglia, G.D.; Genovese, C.; Raccuia, S.A. Cynara cardunculus L. as a Multipurpose Crop for Plant Secondary Metabolites Production in Marginal Stressed Lands. Front. Plant Sci. 2020, 11, 240. [Google Scholar] [CrossRef]
- Leonardi, C.; Pappalardo, H.; Genovese, C.; Puglia, G.; Bua, G.D.; Dugo, G.; Raccuia, S.A. Mechanisms of phytoextraction in Cynara cardunculus L. Growing under cadmium and arsenic stress. Acta Hortic. 2016, 1147, 139–144. [Google Scholar] [CrossRef]
- Leonardi, C.; Toscano, V.; Pappalardo, H.; Genovese, C.; Raccuia, S.A. The potential of Cynara cardunculus L. for phytoremediation of heavy metal in contaminated soils. Acta Hortic. 2016. [Google Scholar] [CrossRef]
- Raccuia, S.A.; Melilli, M.G. Biomass and grain oil yields in Cynara cardunculus L. genotypes grown in a Mediterranean environment. Field Crop. Res. 2007, 101, 187–197. [Google Scholar] [CrossRef]
- Gominho, J.; Curt, M.D.; Lourenço, A.; Fernández, J.; Pereira, H. Cynara cardunculus L. as a biomass and multi-purpose crop: A review of 30 years of research. Biomass Bioenergy 2018, 109, 257–275. [Google Scholar] [CrossRef]
- Zidorn, C. Sesquiterpene lactones and their precursors as chemosystematic markers in the tribe Cichorieae of the Asteraceae. Phytochemistry 2008, 69, 2270–2296. [Google Scholar] [CrossRef]
- Eljounaidi, K.; Comino, C.; Moglia, A.; Cankar, K.; Genre, A.; Hehn, A.; Bourgaud, F.; Beekwilder, J.; Lanteri, S. Accumulation of cynaropicrin in globe artichoke and localization of enzymes involved in its biosynthesis. Plant Sci. 2015, 239, 128–136. [Google Scholar] [CrossRef]
- Menin, B.; Comino, C.; Portis, E.; Moglia, A.; Cankar, K.; Bouwmeester, H.J.; Lanteri, S.; Beekwilder, J. Genetic mapping and characterization of the globe artichoke (+)-germacrene A synthase gene, encoding the first dedicated enzyme for biosynthesis of the bitter sesquiterpene lactone cynaropicrin. Plant Sci. 2012, 190, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Scavo, A.; Pandino, G.; Restuccia, A.; Mauromicale, G. Leaf extracts of cultivated cardoon as potential bioherbicide. Sci. Hortic. 2020, 261, 109024. [Google Scholar] [CrossRef]
- Chadwick, M.; Trewin, H.; Gawthrop, F.; Wagstaff, C. Sesquiterpenoids lactones: Benefits to plants and people. Int. J. Mol. Sci. 2013, 14, 12780–12805. [Google Scholar] [CrossRef] [Green Version]
- Dufour, J.; Arsuaga, J.; Moreno, J.; Torrealba, H.; Camacho, J. Comparative life cycle assessment of biodiesel production from cardoon (Cynara cardunculus) and rapeseed oil obtained under Spanish conditions. Energy Fuels 2013, 27, 5280–5286. [Google Scholar] [CrossRef]
- Oliet, M.; Gilarranz, M.A.; Domínguez, J.C.; Alonso, M.V.; Rodríguez, F. Ethanol-based pulping from Cynara cardunculus L. J. Chem. Technol. Biotechnol. 2005, 80, 746–753. [Google Scholar] [CrossRef]
- Benlloch-González, M.; Fournier, J.M.; Ramos, J.; Benlloch, M. Strategies underlying salt tolerance in halophytes are present in Cynara cardunculus. Plant Sci. 2005, 168, 653–659. [Google Scholar] [CrossRef]
- Angelini, L.G.; Ceccarini, L.; Nassi o Di Nasso, N.; Bonari, E. Long-term evaluation of biomass production and quality of two cardoon (Cynara cardunculus L.) cultivars for energy use. Biomass Bioenergy 2009, 33, 810–816. [Google Scholar] [CrossRef]
- Raccuia, S.A.; Piscioneri, I.; Sharma, N.; Melilli, M.G. Genetic variability in Cynara cardunculus L. domestic and wild types for grain oil production and fatty acids composition. Biomass Bioenergy 2011, 35, 3167–3173. [Google Scholar] [CrossRef]
- Piscioneri, I.; Sharma, N.; Baviello, G.; Orlandini, S. Promising industrial energy crop, Cynara cardunculus: A potential source for biomass production and alternative energy. Energy Convers. Manag. 2000, 41, 1091–1105. [Google Scholar] [CrossRef]
- Pagano, I.; Piccinelli, A.L.; Celano, R.; Campone, L.; Gazzerro, P.; De Falco, E.; Rastrelli, L. Chemical profile and cellular antioxidant activity of artichoke by-products. Food Funct. 2016, 7, 4841–4850. [Google Scholar] [CrossRef]
- Clifford, M.N.; Knight, S.; Kuhnert, N. Discriminating between the Six Isomers of Dicaffeoylquinic Acid by LC-MS n. J. Agric. Food Chem. 2005, 53, 3821–3832. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Läuchli, A.; Grattan, S.R. Plant Responses to Saline and Sodic Conditions. In Agricultural Salinity Assessment and Management; American Society of Civil Engineers: Reston, VA, USA, 2011; pp. 169–205. ISBN 9780784476482. [Google Scholar]
- Carillo, P.; Grazia, M.; Pontecorvo, G.; Fuggi, A.; Woodrow, P. Salinity Stress and Salt Tolerance. In Abiotic Stress in Plants—Mechanisms and Adaptations; Shanker, A., Venkateswarlu, B., Eds.; IntechOpen: London, UK, 2011; Available online: https://www.intechopen.com/books/abiotic-stress-in-plants-mechanisms-and-adaptations/salinity-stress-and-salt-tolerance (accessed on 31 March 2020). [CrossRef] [Green Version]
- Docimo, T.; De Stefano, R.; De Palma, M.; Cappetta, E.; Villano, C.; Aversano, R.; Tucci, M. Transcriptional, metabolic and DNA methylation changes underpinning the response of Arundo donax ecotypes to NaCl excess. Planta 2020, 251, 34. [Google Scholar] [CrossRef]
- Agathokleous, E.; Belz, R.G.; Kitao, M.; Koike, T.; Calabrese, E.J. Does the root to shoot ratio show a hormetic response to stress? An ecological and environmental perspective. J. For. Res. 2019, 30, 1569–1580. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Motos, J.; Ortuño, M.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.; Hernandez, J. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Freschet, G.T.; Violle, C.; Bourget, M.Y.; Scherer-Lorenzen, M.; Fort, F. Allocation, morphology, physiology, architecture: The multiple facets of plant above- and below-ground responses to resource stress. New Phytol. 2018, 219, 1338–1352. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Andújar, C.; Martínez-Pérez, A.; Ferrández-Ayela, A.; Albacete, A.; Martínez-Melgarejo, P.A.; Dodd, I.C.; Thompson, A.J.; Pérez-Pérez, J.M.; Pérez-Alfocea, F. Impact of overexpression of 9-cis-epoxycarotenoid dioxygenase on growth and gene expression under salinity stress. Plant Sci. 2019, 110268. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walia, H.; Wilson, C.; Condamine, P.; Liu, X.; Ismail, A.M.; Zeng, L.; Wanamaker, S.I.; Mandal, J.; Xu, J.; Cui, X.; et al. Comparative Transcriptional Profiling of Two Contrasting Rice Genotypes under Salinity Stress during the Vegetative Growth Stage. Plant Physiol. 2005, 139, 822–835. [Google Scholar] [CrossRef] [Green Version]
- Lillo, C.; Lea, U.; Ruoff, P. Nutrient depletion as a key factor for manipulating gene expression and product formation in different branches of the flavonoid pathway. Plant. Cell Environ. 2008, 31, 587–601. [Google Scholar] [CrossRef]
- Hernández, I.; Alegre, L.; Van Breusegem, F.; Munné-Bosch, S. How relevant are flavonoids as antioxidants in plants? Trends Plant Sci. 2009, 14, 125–132. [Google Scholar] [CrossRef]
- Di Ferdinando, M.; Brunetti, C.; Fini, A.; Tattini, M. Flavonoids as antioxidants in plants under abiotic stresses. In Abiotic Stress Responses in Plants: Metabolism, Productivity and Sustainability; Ahmad, P., Prasad, M., Eds.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Miret, J.A.; Munné-Bosch, S.; Dijkwel, P.P. ABA signalling manipulation suppresses senescence of a leafy vegetable stored at room temperature. Plant Biotechnol. J. 2018, 16, 530–544. [Google Scholar] [CrossRef] [Green Version]
- Scavo, A.; Rial, C.; Varela, R.M.; Molinillo, J.M.G.; Mauromicale, G.; Macias, F.A. Influence of Genotype and Harvest Time on the Cynara cardunculus L. Sesquiterpene Lactone Profile. J. Agric. Food Chem. 2019, 67, 6487–6496. [Google Scholar] [CrossRef]
- Rial, C.; Novaes, P.; Varela, R.M.; José, J.M.; Macias, F.A. Phytotoxicity of cardoon (Cynara cardunculus) allelochemicals on standard target species and weeds. J. Agric. Food Chem. 2014, 62, 6699–6706. [Google Scholar] [CrossRef]
- Blanco, E.; Sabetta, W.; Danzi, D.; Negro, D.; Passeri, V.; De Lisi, A.; Paolocci, F.; Sonnante, G. Isolation and Characterization of the Flavonol Regulator CcMYB12 from the Globe Artichoke [Cynara cardunculus var. scolymus (L.) Fiori]. Front. Plant Sci. 2018, 9, 941. [Google Scholar] [CrossRef] [Green Version]
- Soshinkova, T.N.; Radyukina, N.L.; Korolkova, D.V.; Nosov, A.V. Proline and functioning of the antioxidant system in Thellungiella salsuginea plants and cultured cells subjected to oxidative stress. Russ. J. Plant Physiol. 2013, 60, 41–54. [Google Scholar] [CrossRef]
- Schwanz, P.; Polle, A. Differential stress responses of antioxidative systems to drought in pendunculate oak (Quercus robur) and maritime pine (Pinus pinaster) grown under high CO2 concentrations. J. Exp. Bot. 2001, 52, 133–143. [Google Scholar] [PubMed]
- Jabeen, N.; Ahmad, R. The activity of antioxidant enzymes in response to salt stress in safflower (Carthamus tinctorius L.) and sunflower (Helianthus annuus L.) seedlings raised from seed treated with chitosan. J. Sci. Food Agric. 2013, 93, 1699–1705. [Google Scholar] [CrossRef] [PubMed]
- De Stefano, R.; Cappetta, E.; Guida, G.; Mistretta, C.; Caruso, G.; Giorio, P.; Albrizio, R.; Tucci, M. Screening of giant reed (Arundo donax L.) ecotypes for biomass production under salt stress. Plant Biosyst. — Int. J. Deal. Asp. Plant Biol. 2017, 3504, 1–7. [Google Scholar] [CrossRef]
- Salahuddin, S.; Rewald, B.; Razaq, M.; Lixue, Y.; Li, J.; Khan, F.; Jie, Z. Root order-based traits of Manchurian walnut & larch and their plasticity under interspecific competition. Sci. Rep. 2018, 8, 1–14. [Google Scholar]
- Celano, R.; Piccinelli, A.L.; Pagano, I.; Roscigno, G.; Campone, L.; De Falco, E.; Russo, M.; Rastrelli, L. Oil distillation wastewaters from aromatic herbs as new natural source of antioxidant compounds. Food Res. Int. 2017, 99, 298–307. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pfaffl, M.W. Quantification strategies in real-time PCR. A-Z Quant. PCR 2004, 1, 87–112. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Parameters | NaCl mM | “Bianco Avorio” | “Spagnolo” |
|---|---|---|---|
| last leaf width mm | 0 | 47.44 ± 6.61 a | 42.45 ± 6.56 a |
| 100 | 43.17 ± 4.16 a | 46.49 ± 5.00 a | |
| shoot FW g plant−1 | 0 | 22.35 ± 6.74 a | 22.28 ± 6.74 a |
| 100 | 15.02 ± 4.41 b | 15.00 ± 5.11 b | |
| shoot DW g plant−1 | 0 | 1.37 ± 0.47 a | 1.27 ± 0.44 a |
| 100 | 1.07 ± 0.32 a | 1.15 ± 0.49 a | |
| root FW g plant−1 | 0 | 3.19 ± 1.52 b | 3.52 ± 1.24 b |
| 100 | 4.34 ± 1.26 ab | 5.50 ± 1.82 a | |
| root DW g plant−1 | 0 | 0.26 ± 0.18 b | 0.27 ± 0.15 b |
| 100 | 0.39 ± 0.16 ab | 0.46 ± 0.29 a | |
| root length cm | 0 | 22.2 ± 6.58 b | 25.59 ± 7.86 b |
| 100 | 27.31 ± 8.36 ab | 37.81±14.55 a | |
| R:S | 0 | 0.22 ± 0.19 b | 0.23 ± 0.11 b |
| 100 | 0.34 ± 0.13 a | 0.39 ± 0.14 a | |
| SRL cm g−1 | 0 | 82.5 ± 33.3 a | 86.5 ± 36.4 a |
| 100 | 86.5 ± 41.1 a | 97.5 ± 41.3 a |
| Time (day) | “Bianco Avorio” | “Spagnolo” | ||
|---|---|---|---|---|
| 0 mM NaCl | 100 mM NaCl | 0 mM | 100 mM NaCl | |
| 0 | 22.45 ± 5.62 a | 25.16 ± 6.25 a | 23.13 ± 3.62 a | 23.66 ± 4.99 a |
| 2 | 20.47 ± 4.61 a | 20.75 ± 4.70 a | 23.39 ± 2.72 a | 24.73 ± 3.87 a |
| 21 | 26.88 ± 6.70 a | 32.42 ± 5.95 a | 29.77 ± 7.55 a | 26.28 ± 6.82 a |
| Time (Day) | Genotypes | NaCl (mM) | CQA Tot | 5-CQA | diCQAs | Lgly | L |
|---|---|---|---|---|---|---|---|
| mg/100 g DW | |||||||
| 2 | Bianco Avorio | 0 | 2861.0 ± 753 a | 893.0 ± 68.8 a | 1968.0 ± 794.0 a | 540.3 ± 163.0 a | 8.2 ± 3.0 a |
| 100 | 1502.3 ± 351.5 b | 599.3 ± 202.8 b | 903.1 ± 311.0 ab | 308.8 ± 53.0 b | 6.0 ± 2.1 ab | ||
| Spagnolo | 0 | 698.3 ± 202.5 c | 346.2 ± 133.3 c | 352.0 ± 128.3 b | 113.2 ± 31.5 c | 2.0 ± 0.1 c | |
| 100 | 578.2 ± 84.1 c | 220.0 ± 51.8 c | 358.2 ± 36.7 b | 50.0 ± 2.8 d | 3.1 ± 0.2 bc | ||
| 21 | Bianco Avorio | 0 | 8.2 ± 2.6 b | 4.5 ± 0.9 b | 3.6 ± 1.8 b | 0.8 ± 0.1 b | 2.3 ± 1.4 b |
| 100 | 285.7 ± 61.9 a | 156.2 ± 46.0 a | 129.5 ± 28.2 a | 17.2 ± 1.9 a | 9.0 ± 3.0 a | ||
| Spagnolo | 0 | 8.9 ± 1.4 b | 4.8 ± 0.6 b | 4.1 ± 1.8 b | 3.5 ± 1.9 b | 4.3 ± 0.9 b | |
| 100 | 248.9 ± 62.1 a | 83.0 ± 28.2 a | 165.9 ± 34.7 a | 57.9 ± 25.9 a | 13.4 ± 5.8 a | ||
| Time (Days) | Genotypes | NaCl (mM) | SOD (U mg−1 Protein) | CAT µmol H2O2 min−1 mg−1 Protein | APX µmol ASA min−1 mg−1 Protein |
|---|---|---|---|---|---|
| 2 | Bianco Avorio | 0 | 571.7 ± 81.8 a | 0.54 ± 0.05 a | 9.30 ± 1.52 a |
| 100 | 253.6 ± 9.8 b | 0.31 ± 0.14 b | 2.26 ± 0.90 b | ||
| Spagnolo | 0 | 236.9 ± 29.6 a | 0.37 ± 0.06 a | 3.58 ± 1.54 b | |
| 100 | 200.7 ± 15.3 b | 0.24 ± 0.05 b | 6.46 ± 0.92 a | ||
| 21 | Bianco Avorio | 0 | 358.4 ± 68.9 a | 0.39 ± 0.03 a | 3.21 ± 1.04 a |
| 100 | 279.3 ± 23.6 b | 0.35 ± 0.05 a | 1.79 ± 0.21 b | ||
| Spagnolo | 0 | 383.5 ± 25.2 a | 0.34 ± 0.02 a | 2.71 ± 0.29 a | |
| 100 | 258.6 ± 60.6 b | 0.31 ± 0.05 a | 1.62 ± 0.87 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Docimo, T.; De Stefano, R.; Cappetta, E.; Piccinelli, A.L.; Celano, R.; De Palma, M.; Tucci, M. Physiological, Biochemical, and Metabolic Responses to Short and Prolonged Saline Stress in Two Cultivated Cardoon Genotypes. Plants 2020, 9, 554. https://doi.org/10.3390/plants9050554
Docimo T, De Stefano R, Cappetta E, Piccinelli AL, Celano R, De Palma M, Tucci M. Physiological, Biochemical, and Metabolic Responses to Short and Prolonged Saline Stress in Two Cultivated Cardoon Genotypes. Plants. 2020; 9(5):554. https://doi.org/10.3390/plants9050554
Chicago/Turabian StyleDocimo, Teresa, Rosalba De Stefano, Elisa Cappetta, Anna Lisa Piccinelli, Rita Celano, Monica De Palma, and Marina Tucci. 2020. "Physiological, Biochemical, and Metabolic Responses to Short and Prolonged Saline Stress in Two Cultivated Cardoon Genotypes" Plants 9, no. 5: 554. https://doi.org/10.3390/plants9050554
APA StyleDocimo, T., De Stefano, R., Cappetta, E., Piccinelli, A. L., Celano, R., De Palma, M., & Tucci, M. (2020). Physiological, Biochemical, and Metabolic Responses to Short and Prolonged Saline Stress in Two Cultivated Cardoon Genotypes. Plants, 9(5), 554. https://doi.org/10.3390/plants9050554