1. Introduction
Sugarcane (
Saccharum officinarum L.) is an important crop cultivated globally, mainly for sugar and biofuel. In the Philippines, it contributes approximately 70 billion Philippine pesos (PHP) annually and is currently one of the top agricultural crops in the country, in terms of area harvested (437,070 hectares devoted) and yield (29.29.7 million metric tons), which is equivalent to a national yield average of 67 tons/ha [
1]. Variable weather conditions, such as extreme dry spells and excessive rains, were identified as one of the chief causes of low sugarcane production in the Philippines and elsewhere [
2].
Drought is one of the major abiotic stresses experienced by sugarcane grown under rainfed conditions, causing a negative impact on crop development and productivity. Studies have found that with limited moisture, cane yield and total dry matter can be reduced by 17–52% and 20–56%, respectively [
3]. Yield components, such as the number of millable canes, stalk diameter, and cane length, were also adversely affected [
4,
5,
6]. Leaf traits, such as leaf area, leaf number, and leaf area index were also reduced [
4,
7]. During drought, photosynthesis is highly affected, as is stomatal conductance [
7,
8,
9]. In one study, the monitoring of photosynthetic activities under water deficit showed reduced daily leaf carbon balance due to drought [
8]. Recent reports have also shown the ability of photosynthetic machinery to recover after the removal of stress [
9].
Since sugar cane is a crop with high water requirements, planting less water-requiring or drought-tolerant varieties is recommended as one of the strategies to meet the challenge of water depletion [
10]. The sugarcane varieties cultivated in the Philippines were developed for high cane yield, and disease and pest resistance. Although these varieties have been evaluated under a wide range of environmental conditions, evaluation focused on performance under limiting and excessive moisture conditions has not yet been conducted. The opportunity for varietal improvement for drought tolerance exists as shown by significant differences in varietal response to drought under greenhouse and field conditions, covering both agronomic and physiological parameters [
6,
11,
12,
13,
14,
15,
16]. Understanding the different mechanisms that affect the drought response of a crop will be useful in selecting and identifying genotypes with potential drought tolerance.
The sugarcane life cycle consists of distinct growth stages that require a steady water supply to maintain normal growth and development [
17]. Earlier studies have shown the impact of limited moisture on agronomic and physiological traits at formative stages [
11,
18,
19,
20,
21]. These studies have also reported on the differential sensitivity of sugarcane to moisture stress at different growth stages; however, there have been no studies yet on the monitoring of sugarcane response to drought imposed at two formative stages (tillering and stalk elongation) under field conditions.
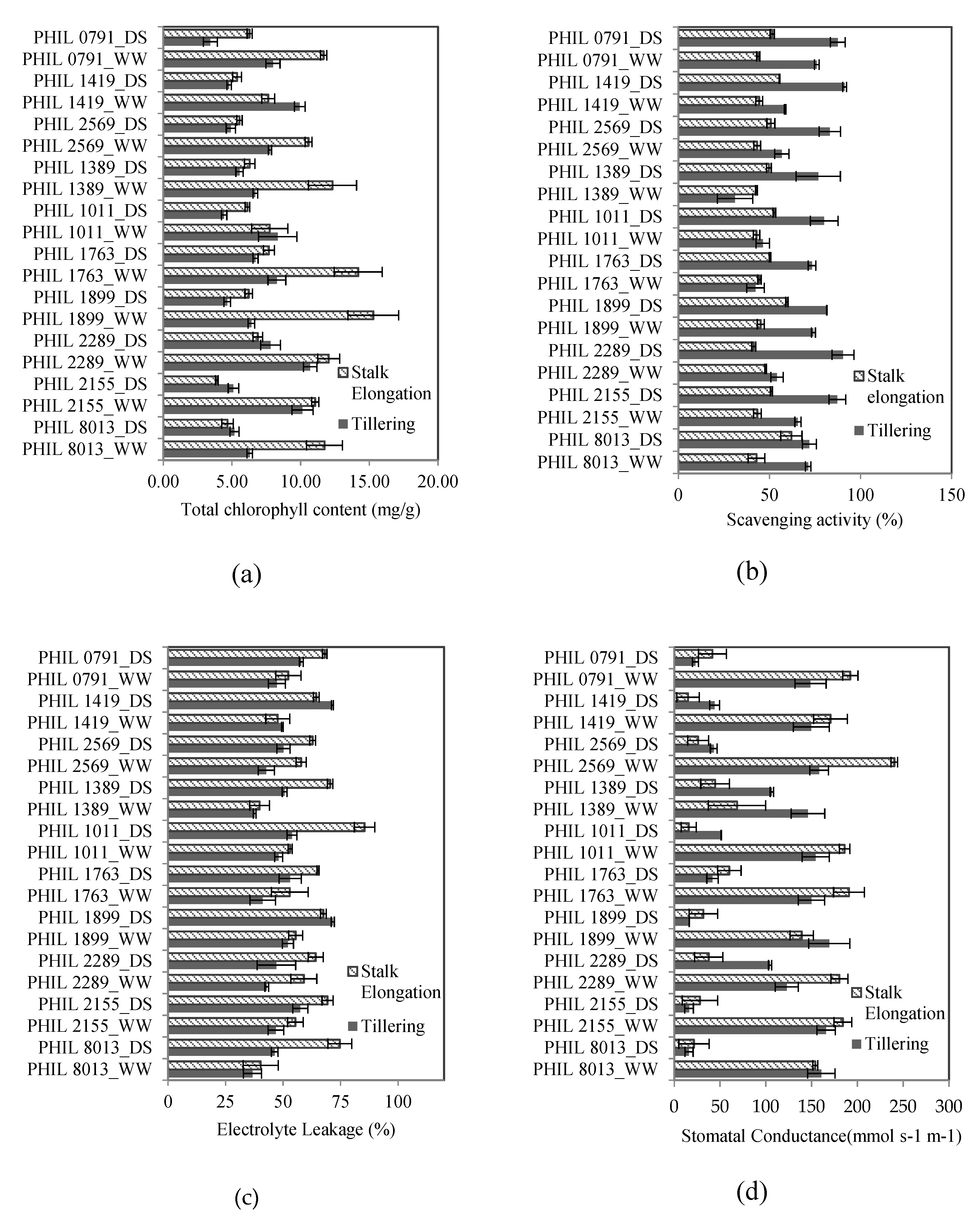
Thus, this study compared the differences in the responses of 10 sugarcane varieties to drought at two formative stages (tillering and stalk elongation). The physiological parameters used to evaluate the water treatment (E) (drought stress vs. well-watered) responses of each sugarcane genotype (G) at each formative stage (T) were total chlorophyll content (Chl), total scavenging/antioxidant activity (AOA), electrolyte leakage (EC), and stomatal conductance (Gs). Moreover, the agronomic parameters that were used to evaluate yield were plant height (PH), stalk diameter (SD), internode length (IL), and tiller count (TIL).
3. Discussion
Drought has always been an important factor affecting plant growth. It triggers different biochemical and physiological responses that could alter several morpho-agronomic characteristics related to yield. In the current study, it is hypothesized that the different formative stages of sugarcane have varying sensitivity to drought could be governed by several critical biochemical and physiological traits. Several studies have already compared the response of different crops at different developmental stages of growth, especially between vegetative and reproductive stages. Studies in rice [
24], sorghum [
25], and wheat [
26] suggest that drought at different developmental stages of growth influence both the agronomic and physiological responses of the crop. Critical growth stages (i.e., tillering and stalk elongation) of sugarcane have been identified, both of which are part of the vegetative phase [
17]. However, studies comparing the effect of drought at these growth stages on different genotypes have not yet been addressed. Therefore, this study discusses the differences in the agro-physiological response of sugarcane subjected to drought at a particular formative stage, to assist in the identification of tolerant and susceptible sugarcane genotypes.
Different parameters can be used to evaluate the effects of drought [
27,
28,
29]. Chlorophyll content has been widely used as a parameter for stress evaluation [
30]. Several studies have reported the significant effect of drought on the total chlorophyll content in sugarcane. In this study, the average chlorophyll content in the drought stress treatment was significantly lower than the well-watered treatment. Results were similar to other reported studies [
27,
30], although the present study utilized pigment extraction analysis instead of the SPAD index estimate in order to evaluate chlorophyll content directly. Chlorophyll reduction due to chloroplast pigment and membrane degradation was associated with the production of free radicals during water deficit conditions [
29]. Higher chlorophyll content during drought was identified as a good indicator of tolerance because it connotes less chloroplast membrane damage [
29]. As a major component of the chloroplast, chlorophyll content has been reported and argued to be correlated with yield maintenance during stressed conditions [
31] and, therefore, it may play a vital role in the photosynthetic capability of a crop under stress.
Higher average chlorophyll content values were observed in plants in the tillering stage (6.76 mg/g) and the stalk elongation stage (8.66 mg/g). Chlorophyll content of most plant species was observed to be significantly lower during its early vegetative stages and increases as it reaches maturity [
32]. Moreover, the intense growth stage of sugarcane, which coincides with the tillering stage in this study, was reported to be more susceptible to drought stress [
33].
In order to alleviate membrane damage due to free radicals or reactive oxygen species (ROS), an efficient scavenging system to reduce the concentration of these harmful molecules is needed, which also serves as a signaling system to ensure induction of gene expression of different antioxidant enzymes related to drought tolerance [
22,
34]. This is evident in the higher antioxidant levels of the drought-stressed plants compared to the well-watered plants in this study. The ROS scavenging activity of different sugarcane genotypes was highly dependent on the inherent adaptive mechanism of each variety [
35]. In this study, the average scavenging percentage values of each genotype were observed to be significantly different from each other, suggesting possible variation in terms of drought tolerance capability. Sugarcane species with higher levels of scavenging/antioxidant activity through different enzymes, such as superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX), tend to perform better under abiotic stresses, such as water deficit [
27,
35,
36,
37,
38]. However, this study evaluated total antioxidant activity through DPPH antioxidant assay, which does not account for the specific effect of each enzymatic or non-enzymatic antioxidant defense mechanism.
DPPH is considered to be a model of a stable lipophilic radical. Antioxidants react with DPPH, reducing the number of DPPH free radicals to the number of their available hydroxyl groups [
39]. The amount of DPPH free radical scavenged accounted for the total amount of antioxidants present in the sample. Using DPPH assay, antioxidant activity in extracts from the sugarcane leaves of each genotype varies, suggesting a difference in the total antioxidant activity and ROS-reducing capacity of each variety [
39]. An increase in concentrations of several antioxidants (i.e., CAT and APX) during drought stress, especially during the tillering or early vegetative stages of sugarcane, have been observed [
33]. These past findings are consistent with this study’s results, which observed higher scavenging activity during drought at the tillering stage. Furthermore, increases in antioxidant concentration (CAT, peroxidase, and glutathione reductase) during drought at different stages of the growth of maize were significantly different across genotypes. It was also observed that maize exposure to drought in its early stages of growth (4 to 5 leaf stage) leads to higher antioxidant activity compared to drought exposure during the anthesis and grain filling stages [
34].
Membrane damage due to oxidative stress could be evaluated by measuring the electrolyte leakage of leaf samples. During drought or heat stress, membrane permeability increases; therefore, electrolytes and ions can easily diffuse [
23]. Measuring the electrical conductivity could help assess the relative membrane injury of the leaf samples. Results showed an increase in the EC values of leaf samples from the drought-stressed plots. These results agree with previous studies in sugarcane [
20,
40], which found that elevated levels of electrolyte leakage were indicative of relative injury in sensitive sugarcane genotypes during moisture stress conditions. Results also suggest that most of the genotypes (i.e., PHIL 2006-1899, PHIL 2000-1419, and PHIL 2000-0791) with higher levels (between 62.6–64.5%) of AOA tend to have lower EC (between 49.1–56.33%) compared to genotypes PHIL 2004-1011, PHIL 2005-1763, and PHIL 2003-1389, with AOA values ranging from 52.7% to 55.6% and corresponding EC values ranging from 52.8% to 60.8%. This trend is also evident in the Pearson’s correlation analysis showing a significant negative correlation between AOA and EC. This relationship was also observed in durum wheat [
41], coconut [
42], and maize [
34], providing further indication of the variability of EC leakage values between stress-tolerant and susceptible genotypes.
In general, higher EC values can be observed in the stalk elongation plots relative to the tillering plots. Interestingly, plants from the stalk elongation plots have significantly lower AOA concentration than the tillering plots. Differences in electrolyte leakage at different developmental stages of maize and sorghum have been reported, and results showed that pre-flowering dehydration produced higher electrolyte leakage values than post-flowering dehydration, which suggest an apparent difference in AOA activities during both of the stages of growth [
43].
Another widely used indicator of drought response is stomatal conductance. Reduction in stomatal conductance is the immediate response of plants during water deficit conditions, and serves as an indicator of effective plant signaling during stressed conditions [
44]. Significant stomatal conductance reduction during stress could affect photosynthetic processes, such as gas exchange [
40]. Changes in photosynthetic rates by stomatal regulation not only limit gas exchange capacity but also affects CO
2 assimilation that could result in a series of photochemical reactions that lead to oxidative damage to the photosynthetic systems of the plant [
45]. Drought-tolerant genotypes have been observed to have mechanisms to adjust to these particular conditions, such as efficient osmotic adjustment, as well as better antioxidant systems [
22]. This study showed that in general, drought reduced the stomatal conductance of all genotypes evaluated, however, significant variation among genotypes under stress was observed for both growth stages. Also, the maintenance of stomatal conductance close to the non-stress level is genotype- and growth stage-dependent. Sugarcane genotypes experienced a significant reduction under drought [
8,
46]. Correlation analysis also showed a significant negative correlation between stomatal conductance and AOA, which was consistent with other drought-related studies in sugarcane [
45].
The drought tolerance indicators also correlated with different agronomic traits related to yield. It was evident in the results that plant height and internode length were both significantly affected by drought. However, the effect of drought among genotypes varied, which could be attributed to the different mechanisms previously discussed. These results were consistent with previous drought experiments [
6,
47,
48]. Similar results focusing on the effect of drought on several cane attributes and its correlation with each other have also been reported [
17].
On the other hand, this study established a significant correlation between plant height and plant physiological parameters for each growth stage of stress imposition, with r values varying for each formative stage. This was the same with the internode length and the physiological parameters; however, it was only significant during the tillering stage and not during the stalk elongation stage. Drought has a higher impact on dry matter accumulation and leaf area during the tillering stage, which could be supported by the observed effects of drought on the physiological characteristics of sugarcane during the same developmental stage [
49]. Additionally, both stalk height and internode length ultimately dictate the yield of sugarcane; therefore, any decrease in these parameters will result in a reduction in the final yield [
40].
Stalk diameter values for each genotype were also significantly different from the highest average values in PHIL 2155 (33.81 mm) and the lowest values in PHIL 2004-1011 (26.82 mm). Variation in stalk diameter under drought stress was mostly affected by genotype [
48], which is evident in the present study as no significant differences between the formative stages were observed. Correlation between AOA and Gs was present during the stalk elongation while Chl and Gs during the tillering stage. However, both have relatively low coefficient values (0.26–0.30). Drought conditions affected stalk height to a greater extent than stalk diameter [
47].
For the tiller count, the average values among genotypes were not significantly different. However, a significant reduction was observed in the average number of tillers in the drought stress treatment (5.24) in comparison with the control (5.92). The number of tillers was already observed to be significantly reduced in sugarcane plants subjected to drought stress during their formative stages [
30]; this includes tillering and the grand growth stage (stalk elongation stage). Higher tiller count was also observed in the tillering stage (6.40) than the stalk elongation stage (4.76). Similarly, water deficit during the earlier developmental stage of sugarcane tends to promote tillering in order to compensate for the reduced assimilation production during drought [
49].
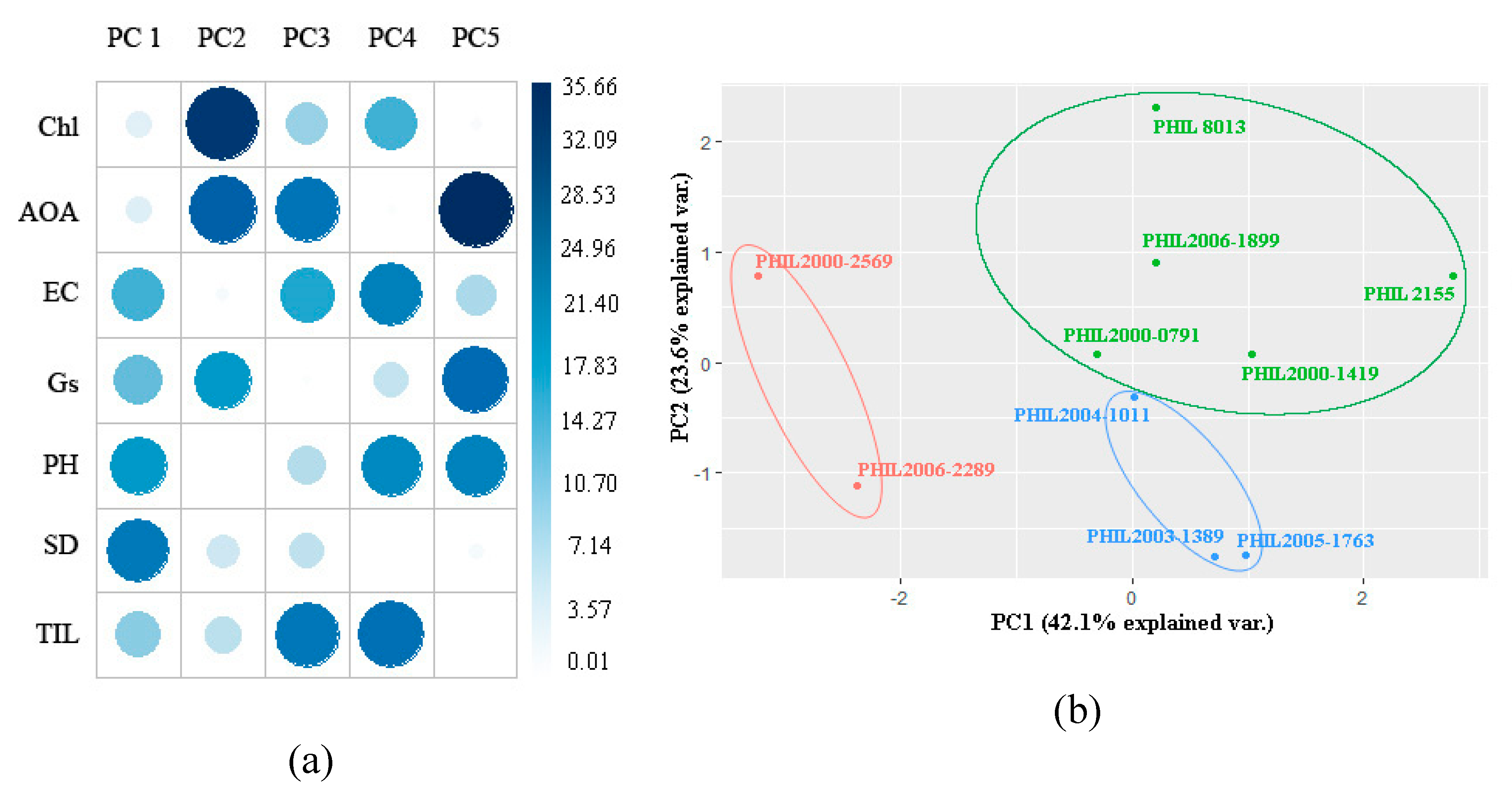
PCA during each of the formative stages produced different sets of groupings, as well as varying contribution values of each parameter. At stalk elongation, the majority of the contributions in PC1 were from EC (17.26), Gs (15.33), plant height (20.51) and stalk diameter (25.51 while 83.58% of the total contributions to the PC2 came from Chl 34.05), AOA (29.06) and Gs, (20.47) while at the tillering stage, Chl (22.62), plant height (23.94), and EC (17.79) contribute highly to PC1, and for PC3, Chl (20.35), AOA (20.75), Gs (15.31), and stalk diameter (30.82) were the major contributors. Most of the physiological parameters were identified as a major contributor to at least one principal component. These parameters were most likely to have the most effect on the overall variation of the data set and could be identified as significant or key contributors to the drought response of each sugarcane genotype during stalk elongation. Higher loadings/contributions of each parameter suggest that the following has the strongest association within each PC, and thus could be used as a selection criterion for selection and breeding for drought tolerance [
50]. Different studies also observed the efficiency of utilizing several physiological parameters in discriminating sugarcane genotypes, as well as different crops [
51,
52,
53,
54].
Furthermore, examining each cluster revealed the similarities in the agro-physiological response of each genotype belonging in the same group at each formative stage. One interesting trend observed in both formative stages was that significantly higher Gs during water deficit conditions appear to have better agronomic characteristics and lower EC values, as well as higher AOA. It is already known how significant stomatal conductance is during drought conditions [
55]. Reduced photosynthetic rate, as well as stomatal conductance during drought, resulted in low levels of internal CO
2 concentration due to inhibition of photosynthetic enzymes such as Rubisco [
45,
56]. This decrease in photosynthetic capacity could limit the growth and development of sugarcane genotypes during drought resulting in poor agronomic performance.
However, PCA-based bi-plots of each experiment showed different clustering for each of the sugarcane genotypes. A possible explanation was the differences in the PCA loading values for each of the developmental stages, as well as changes in the tolerance capability of a genotype as it grows and develops. Sugarcane genotypes at later stages, such as the stalk elongation stage, were more tolerant to drought compared to earlier developmental stages, such as the tillering stage [
56].
This observation provides a possible explanation for the changes in the groupings for both of the formative stages.



{kind=link}
{kind=link}
{kind=link}
{kind=link}



