Salicylic Acid Stimulates Antioxidant Defense and Osmolyte Metabolism to Alleviate Oxidative Stress in Watermelons under Excess Boron

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Plant Growth
2.2. Boron Uptake and Translocation
2.3. Mineral Uptake
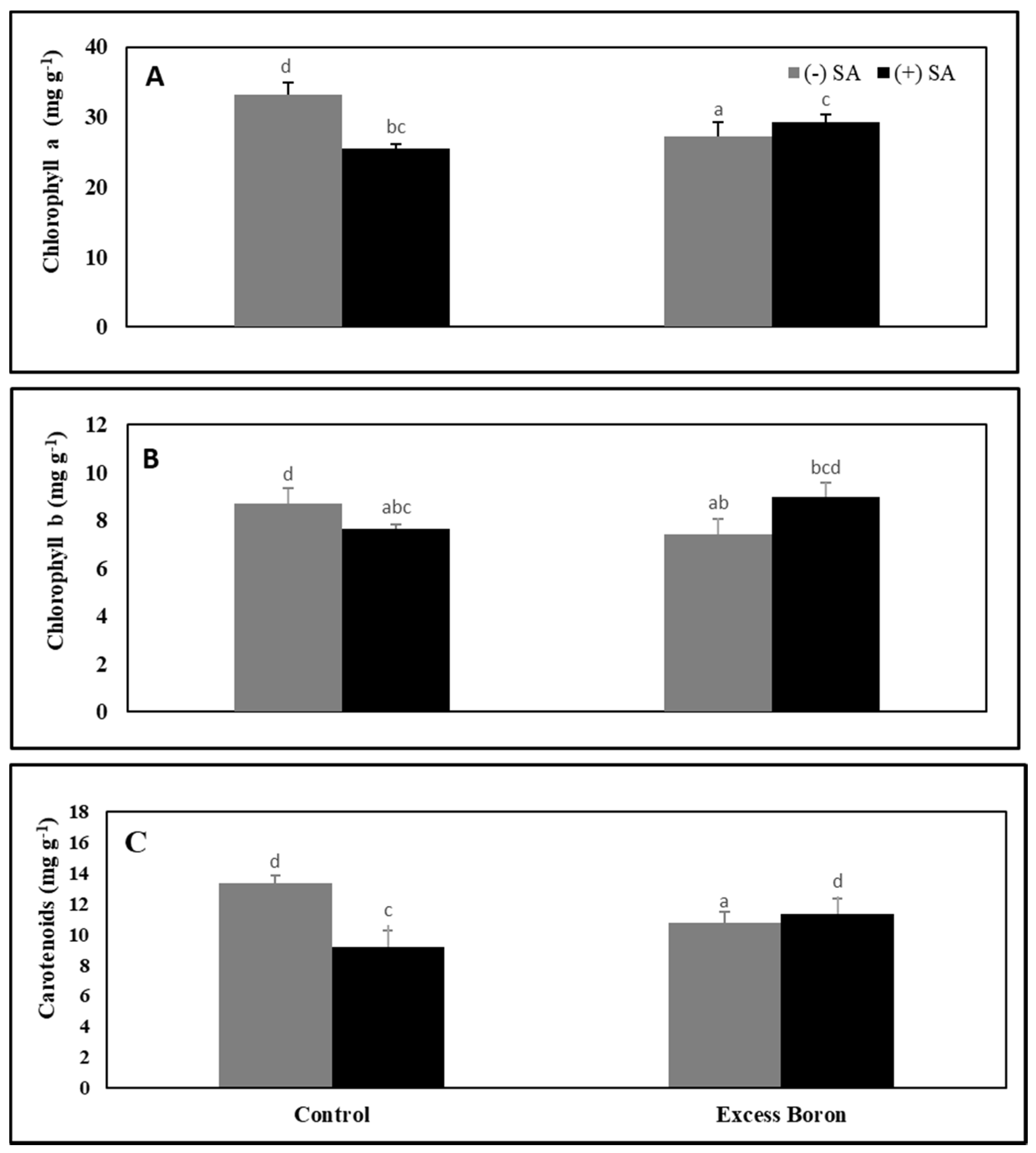
2.4. Chlorophyll and Carotenoid Content
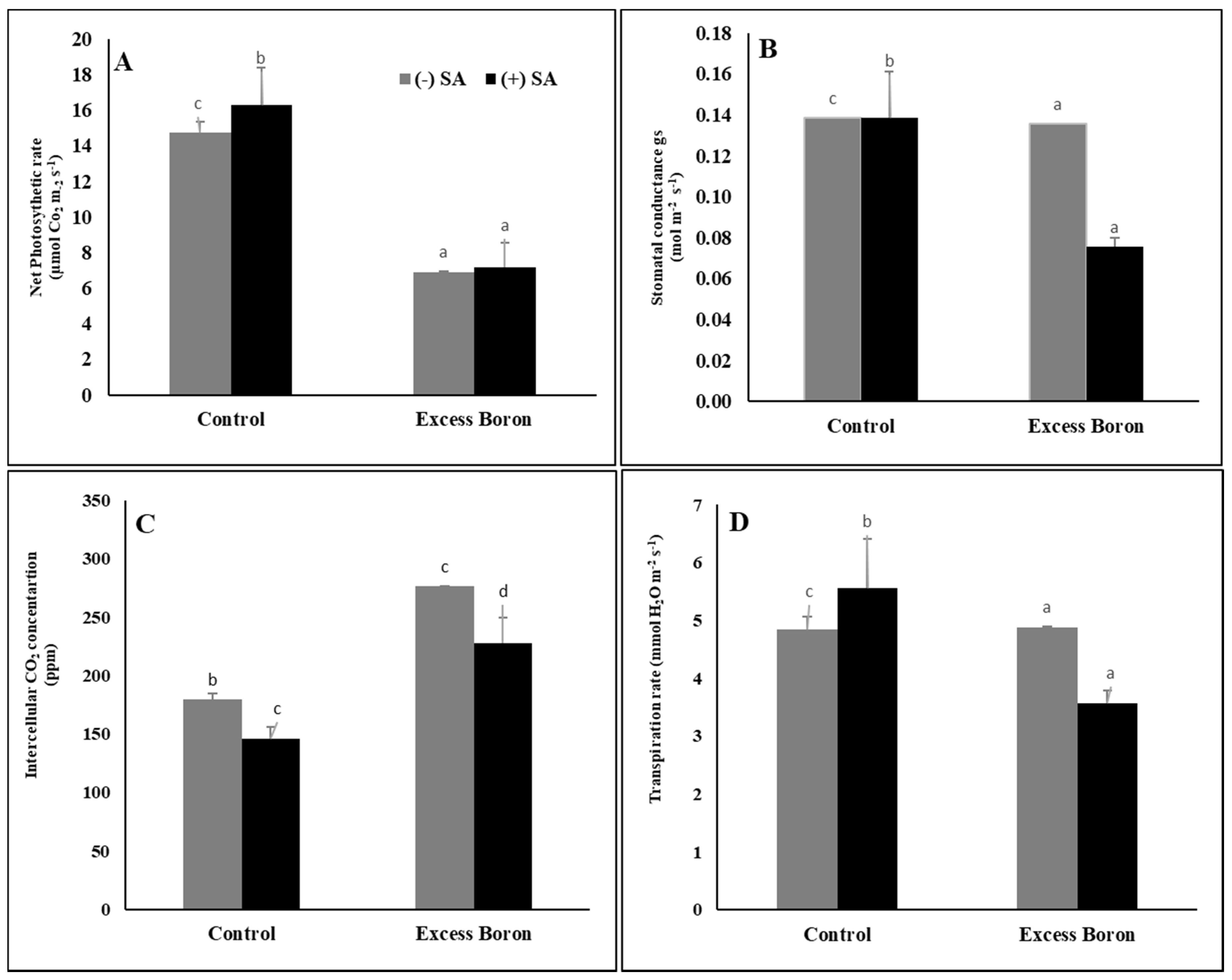
2.5. Leaf Gas Exchange
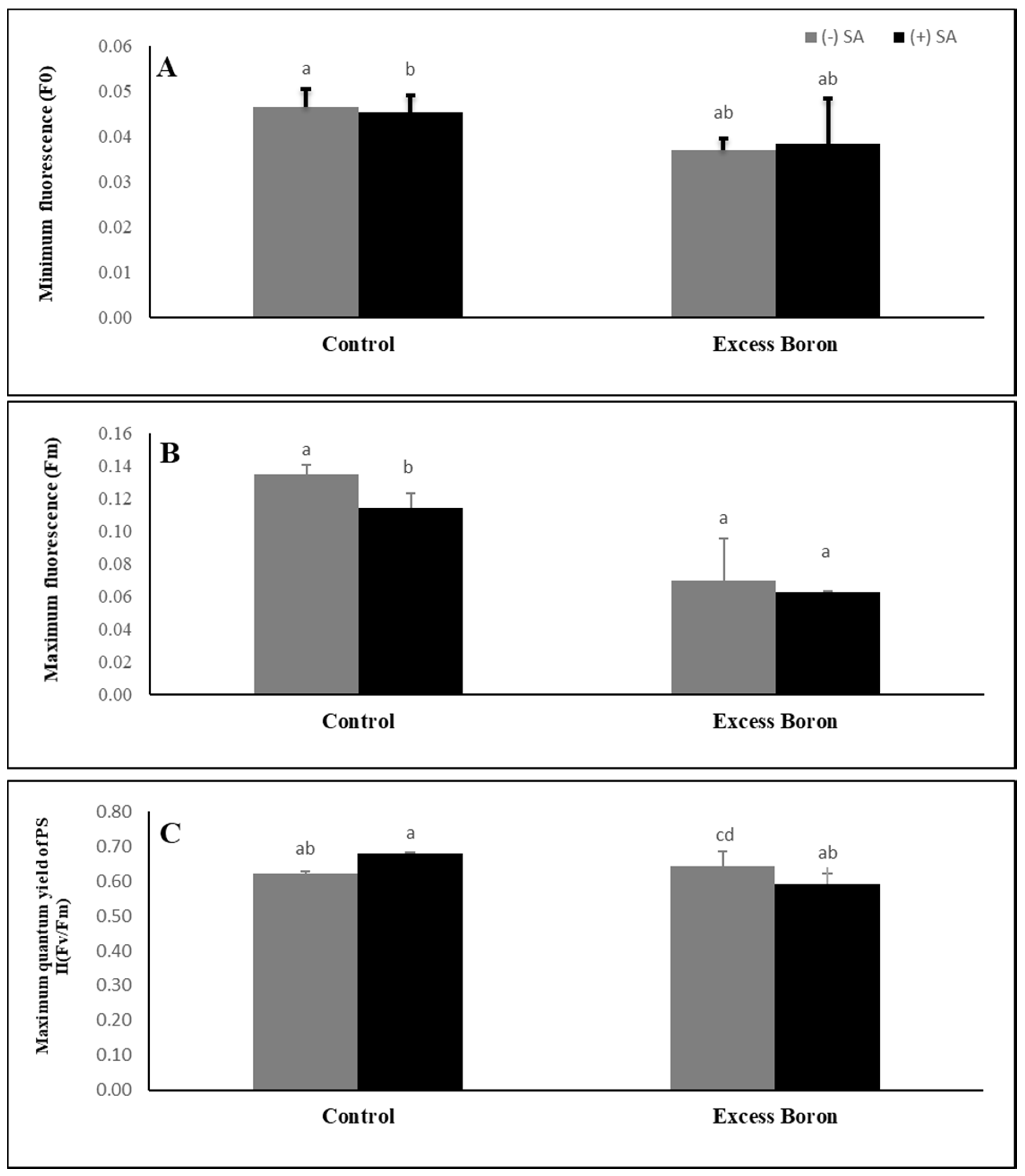
2.6. Chlorophyll Fluorescence
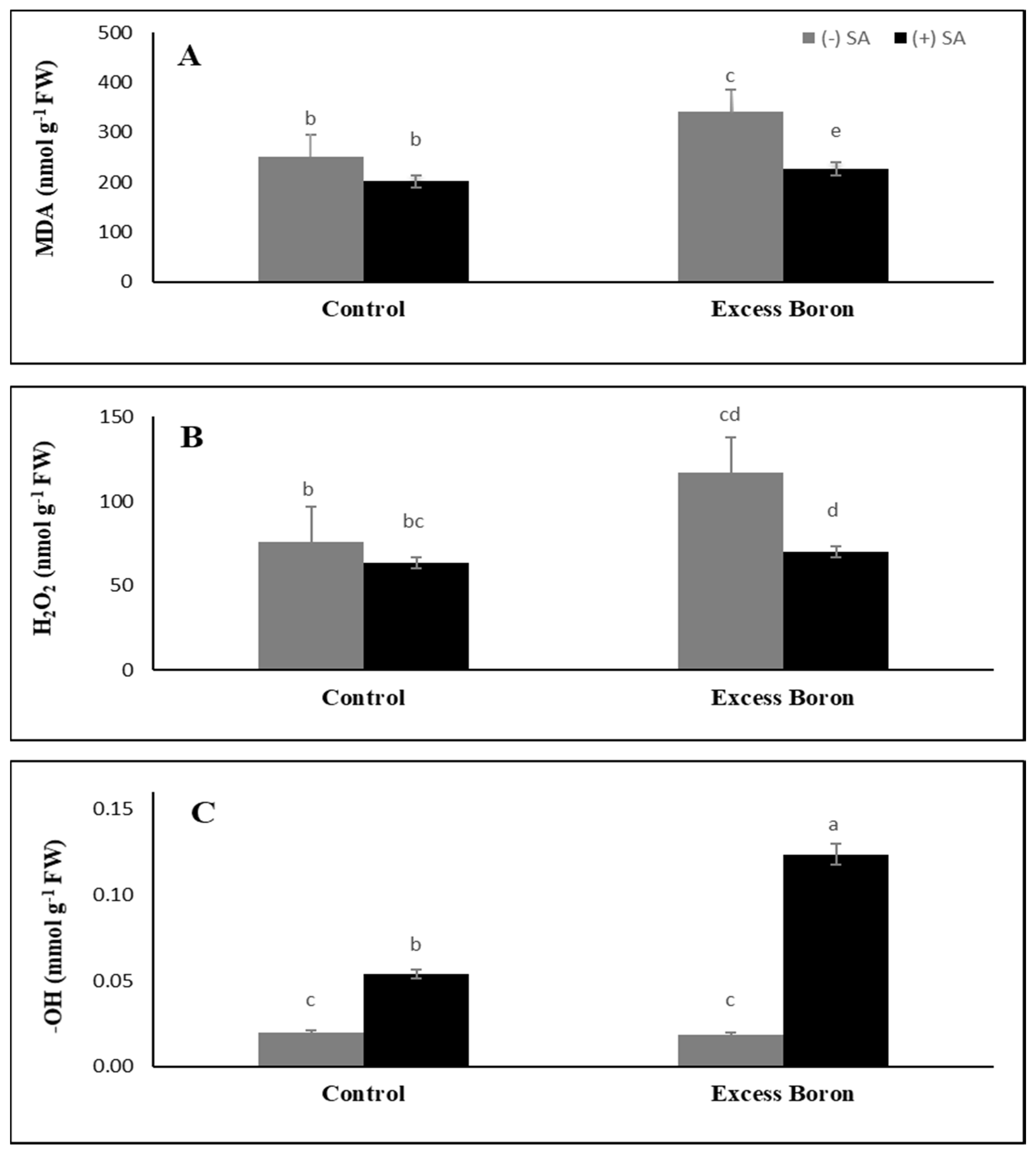
2.7. MDA Content and the Endogenous ROS Generation Rate
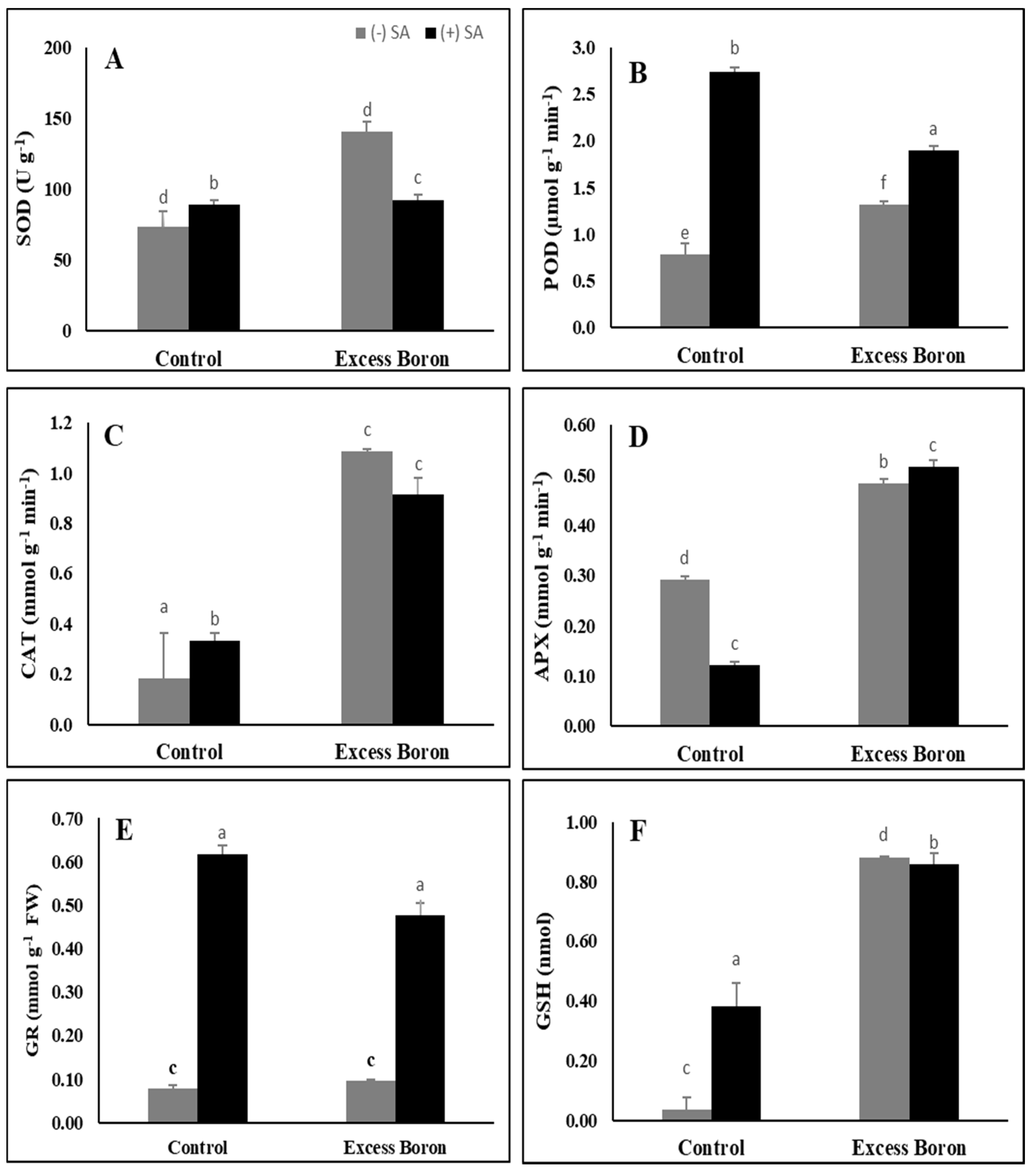
2.8. Antioxidant Enzyme Activities
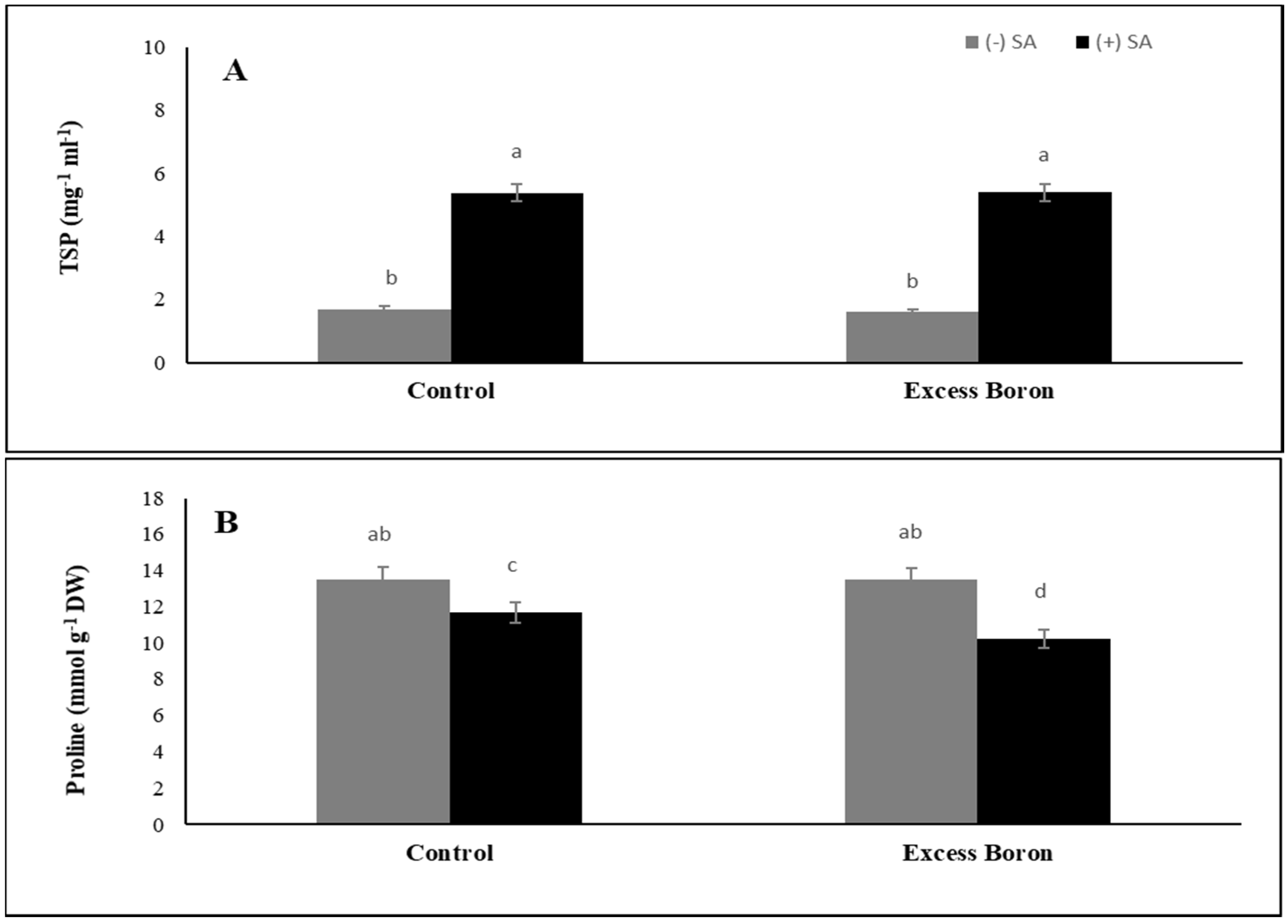
2.9. Total Soluble Protein (TSP) and Proline Content
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Treatments
4.2. Morphological Measurements
4.3. Determination of B and Uptake of Other Nutrients
4.4. Malondialdehyde (MDA) and Reactive Oxygen Species (ROS)
4.5. Chlorophyll and Carotenoid Determination
4.6. Photosynthetic Gas Exchange Parameters
4.7. Chlorophyll Fluorescence
4.8. Antioxidant Enzymes
4.9. Total Soluble Protein and Proline
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tanaka, M.; Fujiwara, T. Physiological roles and transport mechanisms of boron: Perspectives from plants. Pflügers Arch. Eur. J. Physiol. 2008, 456, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Cervilla, L.M.; Blasco, B.; RÍOs, J.J.; Romero, L.; Ruiz, J.M. Oxidative stress and antioxidants in tomato (solanum lycopersicum) plants subjected to boron toxicity. Ann. Bot. 2007, 100, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Moustafa-Farag, M.; Bingsheng, F.; Malangisha Guy, K.; Hu, Z.; Yang, J.; Zhang, M. Activated antioxidant enzymes-reduced malondialdehyde concentration, and improved mineral uptake-promoted watermelon seedlings growth under boron deficiency. J. Plant Nutr. 2016, 39, 1989–2001. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Najeeb, U.; Yang, J.; Hu, Z.; Fang, Z.M. Nitric oxide protects carbon assimilation process of watermelon from boron-induced oxidative injury. Plant Physiol. Biochem. 2017, 111, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Lovatt, C.J.; Bates, L.M. Early effects of excess boron on photosynthesis and growth of Cucurbita pepo. J. Exp. Bot. 1984, 35, 297–305. [Google Scholar] [CrossRef]
- Ardic, M.; Sekmen, A.H.; Turkan, I.; Tokur, S.; Ozdemir, F. The effects of boron toxicity on root antioxidant systems of two chickpea (Cicer arietinum L.) cultivars. Plant Soil 2009, 314, 99–108. [Google Scholar] [CrossRef]
- Roldán, M.; Belver, A.; Rodríguez-Rosales, P.; Ferrol, N.; Donaire, J.P. In vivo and in vitro effects of boron on the plasma membrane proton pump of sunflower roots. Physiol. Plant. 1992, 84, 49–54. [Google Scholar] [CrossRef]
- Choi, E.Y.; Kolesik, P.; McNeill, A.N.N.; Collins, H.; Zhang, Q.; Huynh, B.L.; Graham, R.; Stangoulis, J. The mechanism of boron tolerance for maintenance of root growth in barley (Hordeum vulgare L.). Plant, Cell Environ. 2007, 30, 984–993. [Google Scholar] [CrossRef]
- Ghanati, F.; Morita, A.; Yokota, H. Induction of suberin and increase of lignin content by excess boron in tobacco cells. Soil Sci. Plant Nutr. 2002, 48, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Senousy, H.H.; Abd Ellatif, S.; Ali, S. Assessment of the antioxidant and anticancer potential of different isolated strains of cyanobacteria and microalgae from soil and agriculture drain water. Environ. Sci. Pollut. Res. 2020, 27, 18463–18474. [Google Scholar] [CrossRef]
- Vwioko, E.; Adinkwu, O.; El-Esawi, M.A. Comparative Physiological, Biochemical, and Genetic Responses to Prolonged Waterlogging Stress in Okra and Maize Given Exogenous Ethylene Priming. Front. Physiol. 2017, 8, 632. [Google Scholar] [CrossRef]
- Mohamed, H.I.; Elsherbiny, E.A.; Abdelhamid, M.T. Physiological and biochemical responses of Vicia faba plants to foliar application with zinc and iron. Gesunde Pflanzen 2016, 68, 201–212. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.M.; Ali, H.M.; Alayafi, A.A.; Ahmad, M. Serratia liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef] [Green Version]
- Elkelish, A.A.; Soliman, M.H.; Alhaithloul, H.A.; El-Esawi, M.A. Selenium protects wheat seedlings against salt stress-mediated oxidative damage by up-regulating antioxidants and osmolytes metabolism. Plant Physiol. Biochem. 2019, 137, 144–153. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; EL-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of sweet pepper with stress tolerance-inducing compounds alleviates salinity stress oxidative damage by mediating the physio-biochemical activities and antioxidant systems. Agronomy 2020, 10, 26. [Google Scholar] [CrossRef] [Green Version]
- Loake, G.; Grant, M. Salicylic acid in plant defence—The players and protagonists. Curr. Opin. Plant Biol. 2007, 10, 466–472. [Google Scholar] [CrossRef]
- Taşgín, E.; Atící, Ö.; Nalbantoğlu, B. Effects of salicylic acid and cold on freezing tolerance in winter wheat leaves. Plant Growth Regul. 2003, 41, 231–236. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alayafi, A.A. Overexpression of Rice Rab7 Gene Improves Drought and Heat Tolerance and Increases Grain Yield in Rice (Oryza sativa L.). Genes 2019, 10, 56. [Google Scholar] [CrossRef] [Green Version]
- Elkelish, A.; Qari, S.H.; Mazrou, Y.S.A.; Abdelaal, K.A.A.; Hafez, Y.M.; Abu-Elsaoud, A.M.; Batiha, G.E.-S.; El-Esawi, M.A.; El Nahhas, N. Exogenous Ascorbic Acid Induced Chilling Tolerance in Tomato Plants Through Modulating Metabolism, Osmolytes, Antioxidants, and Transcriptional Regulation of Catalase and Heat Shock Proteins. Plants 2020, 9, 431. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A. Azospirillum lipoferum FK1 confers improved salt tolerance in chickpea (Cicer arietinum L.) by modulating osmolytes, antioxidant machinery and stress-related genes expression. Environ. Exp. Bot. 2019, 159, 55–65. [Google Scholar] [CrossRef]
- Sharma, Y.K.; León, J.; Raskin, I.; Davis, K.R. Ozone-Induced Responses in Arabidopsis thaliana: The Role of Salicylic Acid in the Accumulation of Defense-Related Transcripts and Induced Resistance. Proc. Natl. Acad. Sci. USA 1996, 93, 5099–5104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkeilsh, A.; Awad, Y.M.; Soliman, M.H.; Abu-Elsaoud, A.; Abdelhamid, M.T.; El-Metwally, I.M. Exogenous application of β-sitosterol mediated growth and yield improvement in water-stressed wheat (Triticum aestivum) involves up-regulated antioxidant system. J. Plant Res. 2019, 132, 881–901. [Google Scholar] [CrossRef] [PubMed]
- Metwally, A.; Finkemeier, I.; Georgi, M.; Dietz, K.-J. Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol. 2003, 132, 272–281. [Google Scholar] [CrossRef] [Green Version]
- Palma, J.M.; Sandalio, L.M.; Javier Corpas, F.; Romero-Puertas, M.C.; McCarthy, I.; del Río, L.A. Plant Proteases, Protein Degradation, and Oxidative Stress: Role of Peroxisomes; Elsevier Masson SAS: Paris, France, 2002; Volume 40, pp. 521–530. [Google Scholar]
- Drazic, G.; Mihailovic, N. Modification of cadmium toxicity in soybean seedlings by salicylic acid. Plant Sci. 2005, 168, 511–517. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Wang, G.; Yang, D.; Guan, C.; Ji, J. Foliar application of salicylic acid alleviate the cadmium toxicity by modulation the reactive oxygen species in potato. Ecotoxicol. Environ. Saf. 2019, 172, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Moradkhani, S.; Nejad, R.A.K.; Dilmaghani, K.; Chaparzadeh, N. Salicylic acid decreases Cd toxicity in sunflower plants. Ann. Biol. Res. 2013, 4, 135–141. [Google Scholar]
- Shi, Q.; Zhu, Z. Effects of exogenous salicylic acid on manganese toxicity, element contents and antioxidative system in cucumber. Environ. Exp. Bot. 2008, 63, 317–326. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Guo, K.; Elbaz, A.A.; Yang, Z.M. Salicylic acid alleviates mercury toxicity by preventing oxidative stress in roots of Medicago sativa. Environ. Exp. Bot. 2009, 65, 27–34. [Google Scholar] [CrossRef]
- Metwally, A.M.; Radi, A.A.; El-Shazoly, R.M.; Hamada, A.M. The role of calcium, silicon and salicylic acid treatment in protection of canola plants against boron toxicity stress. J. Plant Res. 2018, 131, 1015–1028. [Google Scholar] [CrossRef]
- Saleem, M.H.; Ali, S.; Rehman, M.; Rana, M.S.; Rizwan, M.; Kamran, M.; Imran, M.; Riaz, M.; Soliman, M.H.; Elkelish, A.; et al. Influence of phosphorus on copper phytoextraction via modulating cellular organelles in two jute (Corchorus capsularis L.) varieties grown in a copper mining soil of Hubei Province, China. Chemosphere 2020, 248, 126032. [Google Scholar] [CrossRef]
- El-Feky, S.S.; El-Shintinawy, F.A.; Shaker, E.M.; El-Din, H.A.S. Effect of elevated boron concentrations on the growth and yield of barley (hordeum vulgare l.) and alleviation of its toxicity using different plant growth modulators. Aust. J. Crop Sci. 2012, 6, 1687–1695. [Google Scholar]
- El-Shazoly, R.M.; Metwally, A.A.; Hamada, A.M. Salicylic acid or thiamin increases tolerance to boron toxicity stress in wheat. J. Plant Nutr. 2019, 42, 702–722. [Google Scholar] [CrossRef]
- Eraslan, F.; Inal, A.; Pilbeam, D.J.; Gunes, A. Interactive effects of salicylic acid and silicon on oxidative damage and antioxidant activity in spinach (Spinacia oleracea L. cv. Matador) grown under boron toxicity and salinity. Plant Growth Regul. 2008, 55, 207–219. [Google Scholar] [CrossRef]
- Chen, W.; Singh, K.B. The auxin, hydrogen peroxide and salicylic acid induced expression of the Arabidopsis GST6 promoter is mediated in part by an ocs element. Plant J. 1999, 19, 667–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-S.; Wang, J.; Yang, Z.-M.; Wang, Q.-Y.; Lü, B.; Li, S.-Q.; Lu, Y.-P.; Wang, S.-H.; Sun, X. Salicylic acid modulates aluminum-induced oxidative stress in roots of Cassia tora. Acta Bot. Sin. 2004, 46, 819–828. [Google Scholar]
- He, Y.; Liu, Y.; Cao, W.; Huai, M.; Xu, B.; Huang, B. Effects of Salicylic Acid on Heat Tolerance Associated with Antioxidant Metabolism in Kentucky Bluegrass. Crop Sci. 2005, 45, 988–995. [Google Scholar] [CrossRef]
- Wang, L.-J.; Li, S.-H. Thermotolerance and Related Antioxidant Enzyme Activities Induced by Heat Acclimation and Salicylic Acid in Grape (Vitis vinifera L.) Leaves. Plant Growth Regul. 2006, 48, 137–144. [Google Scholar] [CrossRef]
- Gholamnezhad, J.; Sanjarian, F.; Goltapeh, E.M.; Safaie, N.; Razavi, K. Effect of Salicylic Acid on Enzyme Activity in Wheat in Immediate Early Time after Infection with Mycosphaerella Graminicola. Sci. Agric. Bohem. 2016, 47, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, A.J.; Shaheed, A.I. Effects of salicylic acid and silicon on oxidative damage and antioxidant activity in mung bean cuttings under boron toxicity. J. Univ. Babylon 2012, 22, 335–348. [Google Scholar]
- Ozturk, M.; Sakcali, S.; Gucel, S.; Tombuloglu, H. Boron and Plants. In Plant Adaptation and Phytoremediation; Ashraf, M., Ozturk, M., Ahmad, M.S.A., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 275–311. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, Q.Q.; Tong, L.H. Boron-deficient soils and their distribution in China. Acta Pedol. Sin. 1980, 17, 228–239. [Google Scholar]
- Aftab, T.; Khan, M.M.A.; Naeem, M.; Idrees, M.; Moinuddin; Teixeira da Silva, J.A.; Ram, M. Exogenous nitric oxide donor protects Artemisia annua from oxidative stress generated by boron and aluminium toxicity. Ecotoxicol. Environ. Saf. 2012, 80, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Aquea, F.; Federici, F.; Moscoso, C.; Vega, A.; Jullian, P.; Haseloff, J.I.M.; Arce-Johnson, P. A molecular framework for the inhibition of Arabidopsis root growth in response to boron toxicity. Plant Cell Environ. 2012, 35, 719–734. [Google Scholar] [CrossRef] [PubMed]
- Josten, P.; Kutschera, U. The Micronutrient Boron Causes the Development of Adventitious Roots in Sunflower Cuttings. Ann. Bot. 1999, 84, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Uzunova, A.N.; Popova, L.P. Effect of Salicylic Acid on Leaf Anatomy and Chloroplast Ultrastructure of Barley Plants. Photosynthetica 2000, 38, 243–250. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Hayat, S.; Ahmad, A. Salicylic Acid Influences Net Photosynthetic Rate, Carboxylation Efficiency, Nitrate Reductase Activity, and Seed Yield in Brassica juncea. Photosynthetica 2003, 41, 281–284. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Almoneafy, A.; Mahmoud, A.; Elkelish, A.; Arnao, M.B.; Li, L.; Ai, S. Melatonin and Its Protective Role against Biotic Stress Impacts on Plants. Biomolecules 2020, 10, 54. [Google Scholar] [CrossRef] [Green Version]
- Arteca, R.N. Plant Growth Substances: Principles and Applications; Chapman & Hall: New York, NY, USA, 1996. [Google Scholar]
- Supanjani, S.; Lee, K.D. Hot pepper response to interactive effects of salinity and boron. Plant Soil Environ. 2006, 52, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Reid, R. Physiology and Metabalism of Boron in Plants. In Advances in Plant and Animal Boron Nutrition; Springer: Dordrecht, The Netherlands, 2007; pp. 83–90. [Google Scholar]
- El-Feky, S.S.; El-Feky, S.S.; El-Shintinawy, F.A.; Shaker, E.M. Role of CaCl2 and salicylic acid on metabolic activities and productivity of boron stressed barley (Hordeum vulgare L.). Int. J. Curr. Microbiol Appl. Sci. 2014, 3, 368–380. [Google Scholar]
- Reid, R.; Fitzpatrick, K. Influence of Leaf Tolerance Mechanisms and Rain on Boron Toxicity in Barley and Wheat. Plant Physiol. 2009, 151, 413–420. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Li, L.; Ren, F.; Lu, P.; Wei, P.; Cai, J.; Xin, L.; Zhang, J.; Chen, J.; Wang, X. Overexpression of the tonoplast aquaporin AtTIP5;1 conferred tolerance to boron toxicity in Arabidopsis. J. Genet. Genom. 2010, 37, 389–397. [Google Scholar] [CrossRef]
- Karabal, E.; Yücel, M.; Öktem, H.A. Antioxidant responses of tolerant and sensitive barley cultivars to boron toxicity. Plant Sci. 2003, 164, 925–933. [Google Scholar] [CrossRef]
- Wimmer, M.A.; Mühling, K.H.; LÄuchli, A.; Brown, P.H.; Goldbach, H.E. Boron Toxicity: The Importance of Soluble Boron. In Boron in Plant and Animal Nutrition; Goldbach, H.E., Brown, P.H., Rerkasem, B., Thellier, M., Wimmer, M.A., Bell, R.W., Eds.; Springer: Boston, MA, USA, 2002; pp. 241–253. [Google Scholar] [CrossRef]
- Namdjoyan, S.; Kermanian, H.; Soorki, A.A.; Modarres Tabatabaei, S.; Elyasi, N. Effects of exogenous salicylic acid and sodium nitroprusside on α-tocopherol and phytochelatin biosynthesis in zinc-stressed safflower plants. Turk. J. Bot. 2018, 42, 271–279. [Google Scholar] [CrossRef]
- Soliman; Alhaithloul; Hakeem; Alharbi; El-Esawi; Elkelish Exogenous Nitric Oxide Mitigates Nickel-Induced Oxidative Damage in Eggplant by Upregulating Antioxidants, Osmolyte Metabolism, and Glyoxalase Systems. Plants 2019, 8, 562. [CrossRef] [PubMed] [Green Version]
- Szalai, G.; Krantev, A.; Yordanova, R.; Popova, L.P.; Janda, T. Influence of salicylic acid on phytochelatin synthesis in Zea mays during Cd stress. Turk. J. Bot. 2013, 37, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Fodor, J.; Gullner, G.; Adam, A.L.; Barna, B.; Kömives, T.; Kiràly, Z. Local and systemic responses of antioxidants to tobacco mosaic virus infection and to salicylic acid in tobacco. Plant Physiol 1997, 114, 1443–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Király, Z.; Barna, B.; Kecskés, A.; Fodor, J. Down-regulation of antioxidative capacity in a transgenic tobacco which fails to develop acquired resistance to necrotization caused by tobacco mosaic virus. Free. Radic. Res. 2002, 36, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Janda, T.; Lejmel, M.A.; Molnár, A.B.; Majláth, I.; Pál, M.; Nguyen, Q.T.; Nguyen, N.T.; Le, V.N.; Szalai, G. Interaction between elevated temperature and different types of Na-salicylate treatment in Brachypodium dystachion. PLoS ONE 2020, 15, e0227608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janda, T.; Szalai, G.; Tari, I.; Paldi, E. Hydroponic treatment with salicylic acid decreases the effect of chilling injury in maize (Zea mays L.) plants. Planta 1999, 208, 175–180. [Google Scholar] [CrossRef]
- Deák, M.; Horváth, G.V.; Davletova, S.; Török, K.; Sass, L.; Vass, I.; Barna, B.; Király, Z.; Dudits, D. Plants ectopically expressing the ironbinding protein, ferritin, are tolerant to oxidative damage and pathogens. Nat. Biotechnol. 1999, 17, 192–196. [Google Scholar] [CrossRef]
- Mohamed, H.I. Molecular and biochemical studies on the effect of gamma rays on lead toxicity in cowpea (Vigna sinensis) plants. Biol Trace Elem Res 2011, 144, 1205–1218. [Google Scholar] [CrossRef]
- Horváth, E.; Szalai, G.; Janda, T. Induction of abiotic stress tolerance by salicylic acid signaling. J. Plant Growth Regul. 2007, 26, 290–300. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Garcia, P.C.; Rivero, R.M.; López-Lefebre, L.R.; Sánchez, E.; Ruiz, J.M.; Romero, L. Response of oxidative metabolism to the application of carbendazim plus boron in tobacco. Aust. J. Plant Physiol. 2001, 28, 801–806. [Google Scholar] [CrossRef]
- Kazemi, N.; Khavari-Nejad, R.A.; Fahimi, H.; Saadatmand, S.; Nejad-Sattari, T. Effects of exogenous salicylic acid and nitric oxide on lipid peroxidation and antioxidant enzyme activities in leaves of Brassica napus L. under nickel stress. Sci. Hortic. 2010, 126, 402–407. [Google Scholar] [CrossRef]
- Noctor, G.; Gomez, L.; Vanacker, H.; Foyer, C.H. Interactions between biosynthesis, compartmentation and transport in the control of glutathione homeostasis and signalling. J. Exp. Bot. 2002, 53, 1283–1304. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.K. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. South Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Cheng, I.F.; Zhao, C.P.; Amolins, A.; Galazka, M.; Doneski, L. A hypothesis for the in vivo antioxidant action of salicyclic acid. Biometals 1996, 9, 285–290. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, H.I. Reactive oxygen species, lipid peroxidation and antioxidative defense mechanism. Notulae Botanicae Horti Agrobotanici ClujNapoca. 2013, 41, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Akladious, S.A.; Mohamed, H.I. Physiological role of exogenous nitric oxide in improving performance, yield and some biochemical aspects of sunflower plant under zinc stress. Acta Biol. Hung. 2017, 68, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, H.I.; Latif, H.H.; Hanafy, R.S. Influence of Nitric Oxide Application on Some Biochemical Aspects, Endogenous Hormones, Minerals and Phenolic Compounds of Vicia faba Plant Grown under Arsenic Stress. Gesunde Pflanzen 2016, 68, 99–107. [Google Scholar] [CrossRef]
- El-Tayeb, M.A.; El-Enany, A.E.; Ahmed, N.L. Salicylic acid-induced adaptive response to copper stress in sunflower (Helianthus annuus L.). Plant Growth Regul. 2006, 50, 191–199. [Google Scholar] [CrossRef]
- Kováčik, J.; Grúz, J.; Hedbávný, J.; Klejdus, B.; Strnad, M. Cadmium and Nickel Uptake Are Differentially Modulated by Salicylic Acid in Matricaria chamomilla Plants. J. Agric. Food Chem. 2009, 57, 9848–9855. [Google Scholar] [CrossRef]
- Yu, M.; Shen, R.; Xiao, H.; Xu, M.; Wang, H.; Wang, H.; Zeng, Q.; Bian, J. Boron alleviates aluminum toxicity in pea (Pisum sativum). Plant Soil 2009, 314, 87–98. [Google Scholar] [CrossRef]
- Wu, C.-y.; Lu, L.-l.; Yang, X.-e.; Feng, Y.; Wei, Y.-y.; Hao, H.-l.; Stoffella, P.J.; He, Z.-l. Uptake, Translocation, and Remobilization of Zinc Absorbed at Different Growth Stages by Rice Genotypes of Different Zn Densities. J. Agric. Food Chem. 2010, 58, 6767–6773. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Leul, M. Uniconazole-induced alleviation of freezing injury in relation to changes in hormonal balance, enzyme activities and lipid peroxidation in winter rape. Plant Growth Regul. 1998, 26, 41–47. [Google Scholar] [CrossRef]
- Yordanov, I.; Velikova, V.; Tsonev, T. Plant Responses to Drought, Acclimation, and Stress Tolerance. Photosynthetica 2000, 38, 171–186. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C.; Aruoma, O.I. The deoxyribose method: A simple “test-tube” assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef]
- Vernon, L.P.; Seely, R.G. The Chlorophylls; Academic Press: New York, NY, USA, 1966. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Zhang, W.F.; Zhang, F.; Raziuddin, R.; Gong, H.J.; Yang, Z.M.; Lu, L.; Ye, Q.F.; Zhou, W.J. Effects of 5-aminolevulinic acid on oilseed rape seedling growth under herbicide toxicity stress. J. Plant Growth Regul. 2008, 27, 159–169. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of Paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Shoot Dry Weight | Root Dry Weight | Project Area (cm2) | Surface Area (cm2) | Avg Diameter (mm) | Root Volume (cm3) | Number of Tips |
|---|---|---|---|---|---|---|---|
| Control | 7.0 ± 0.42 a | 0.17 ± 0.010 b | 42.85 ± 2.57 b | 90 ± 5.4 c | 0.64 ± 0.04 d | 1.48 ± 0.09 b | 3173 ± 190 b |
| Excess B | 1.54 ± 0.09 d | 0.11 ± 0.007 e | 20.25 ± 1.21 e | 59 ± 3.6 e | 0.72 ± 0.04 c | 1.23 ± 0.07 c | 1161 ± 69 e |
| Control + SA | 3.78 ± 0.29 b | 0.29 ± 0.022 a | 51.16 ± 3.99 a | 160 ± 12.5 a | 0.80 ± 0.06 b | 3.20 ± 0.25 a | 4162 ± 324 a |
| Excess B + SA | 2.29 ± 0.22 c | 0.14 ± 0.014 d | 24.03 ± 2.40 d | 73 ± 7.4 d | 0.49 ± 0.05 e | 1.54 ± 0.15 b | 1921 ± 192 d |
| Treatment | B Level in Roots (mg kg−1) | B Level in Leaves (mg kg−1) | Translocation Factor |
|---|---|---|---|
| Control | 73.9 ± 3.05 c,d | 150.9 ± 20 c | 2.04 ± 0.35 b |
| Excess B | 408.0 ± 40 a | 1592.3 ± 400 a | 3.92 ± 1.36 a |
| Control + SA | 80.3 ± 6 c | 90.2 ± 6 c | 1.12 ± 0.07 c |
| Excess B + SA | 276.3 ± 40 b | 1146.3 ± 200 b | 4.18 ± 1.33 a |
| Treatment | Macro-Nutrient (mg kg−1 Shoot DW) | |||
|---|---|---|---|---|
| Ca2+ | K+ | Mg2+ | Na+ | |
| Control | 60.8 ± 5.03 a | 87.5 ± 4 b,c | 16.1 ± 2 a | 15.1 ± 2 c |
| Excess B | 56.9 ± 4 a,b | 79.8 ± 4 c | 14.0 ± 2 a | 19.0 ± 2 b |
| Control + SA | 54.6 ± 4 b | 103.7 ± 20 a | 14.1 ± 2 a | 1.2 ± 0.2 d |
| Excess B + SA | 49.8 ± 4 c | 96.7 ± 12 a,b | 15.4 ± 4 a | 1.7 ± 0.2 d |
| Treatment | Macro-Nutrient (mg kg−1 Root DW) | |||
| Ca2+ | K+ | Mg2+ | Na+ | |
| Control | 12.4 ± 4 c | 107.5 ± 20 a | 3.4 ± 0.4 c | 4.2 ± 0.4 b |
| Excess B | 14.5 ± 4 c | 112.8 ± 20 a | 3.5 ± 0.4 c | 3.4 ± 0.4 e |
| Control + SA | 24.8 ± 2 b | 68.2 ± 4 b | 6.0 ± 0.2 a | 8.9 ± 0.2 a |
| Excess B + SA | 22.7 ± 2 b | 34.8 ± 4 c | 4.9 ± 0.2 b | 4.2 ± 0.2 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moustafa-Farag, M.; Mohamed, H.I.; Mahmoud, A.; Elkelish, A.; Misra, A.N.; Guy, K.M.; Kamran, M.; Ai, S.; Zhang, M. Salicylic Acid Stimulates Antioxidant Defense and Osmolyte Metabolism to Alleviate Oxidative Stress in Watermelons under Excess Boron. Plants 2020, 9, 724. https://doi.org/10.3390/plants9060724
Moustafa-Farag M, Mohamed HI, Mahmoud A, Elkelish A, Misra AN, Guy KM, Kamran M, Ai S, Zhang M. Salicylic Acid Stimulates Antioxidant Defense and Osmolyte Metabolism to Alleviate Oxidative Stress in Watermelons under Excess Boron. Plants. 2020; 9(6):724. https://doi.org/10.3390/plants9060724
Chicago/Turabian StyleMoustafa-Farag, Mohamed, Heba I. Mohamed, Ahmed Mahmoud, Amr Elkelish, Amarendra N. Misra, Kateta Malangisha Guy, Muhammad Kamran, Shaoying Ai, and Mingfang Zhang. 2020. "Salicylic Acid Stimulates Antioxidant Defense and Osmolyte Metabolism to Alleviate Oxidative Stress in Watermelons under Excess Boron" Plants 9, no. 6: 724. https://doi.org/10.3390/plants9060724
APA StyleMoustafa-Farag, M., Mohamed, H. I., Mahmoud, A., Elkelish, A., Misra, A. N., Guy, K. M., Kamran, M., Ai, S., & Zhang, M. (2020). Salicylic Acid Stimulates Antioxidant Defense and Osmolyte Metabolism to Alleviate Oxidative Stress in Watermelons under Excess Boron. Plants, 9(6), 724. https://doi.org/10.3390/plants9060724