Local Factors Rather than the Landscape Context Explain Species Richness and Functional Trait Diversity and Responses of Plant Assemblages of Mediterranean Cereal Field Margins

 , and
, and 
Abstract
:1. Introduction
2. Results
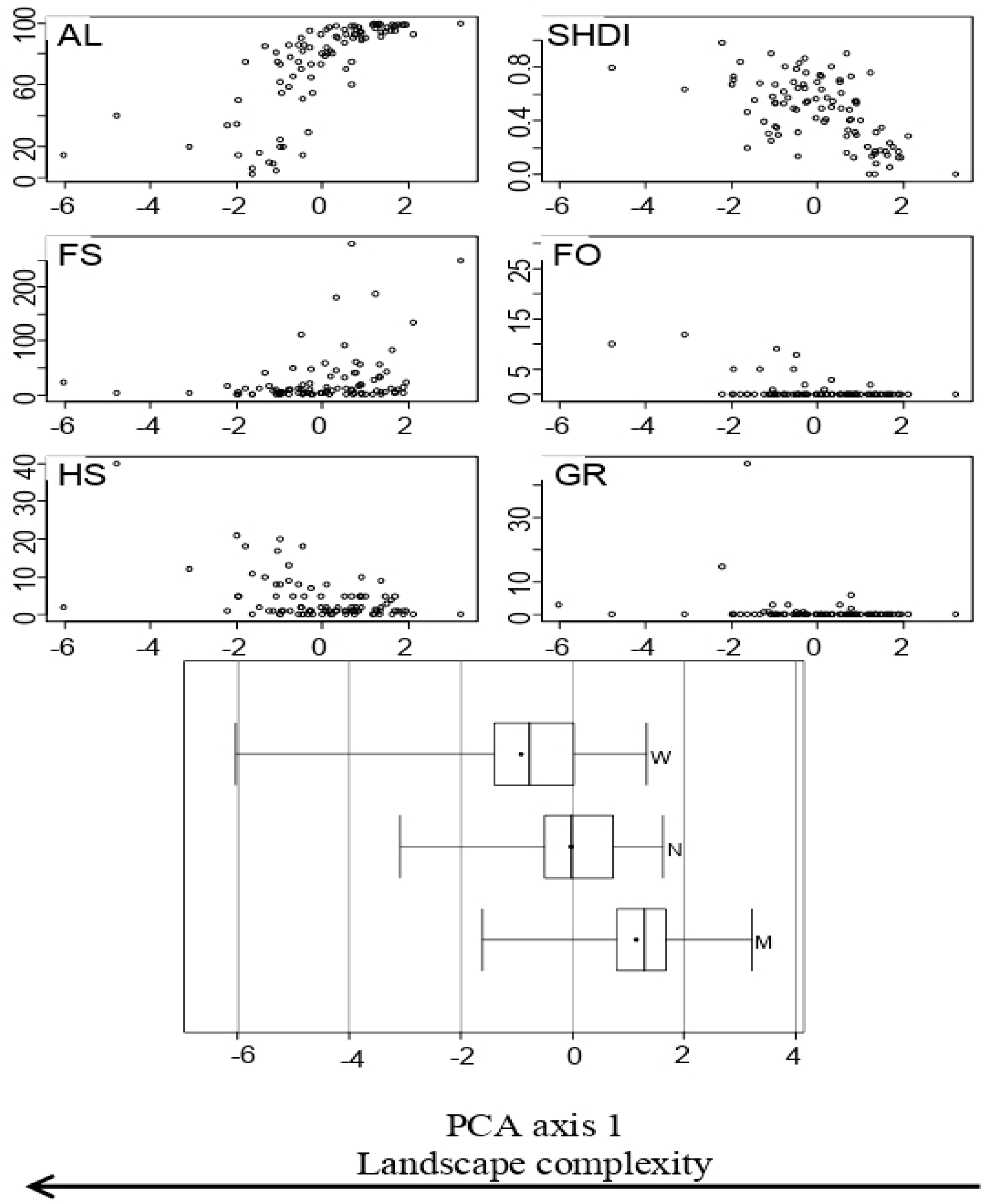
2.1. Landscape Complexity Gradient
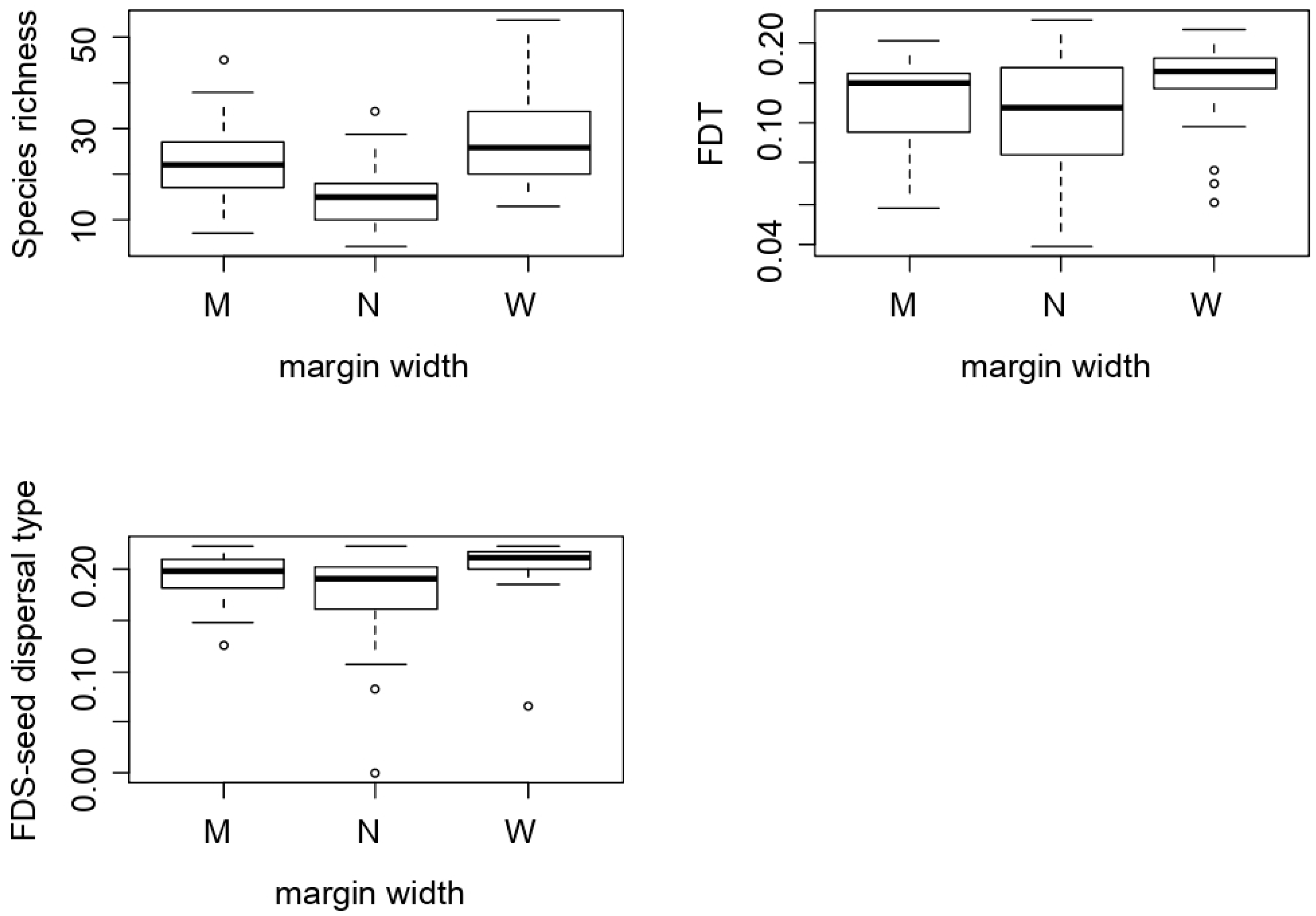
2.2. Agricultural Intensification Effects on Species Richness, Functional Diversity and Functional Traits
3. Discussion
4. Materials and Methods
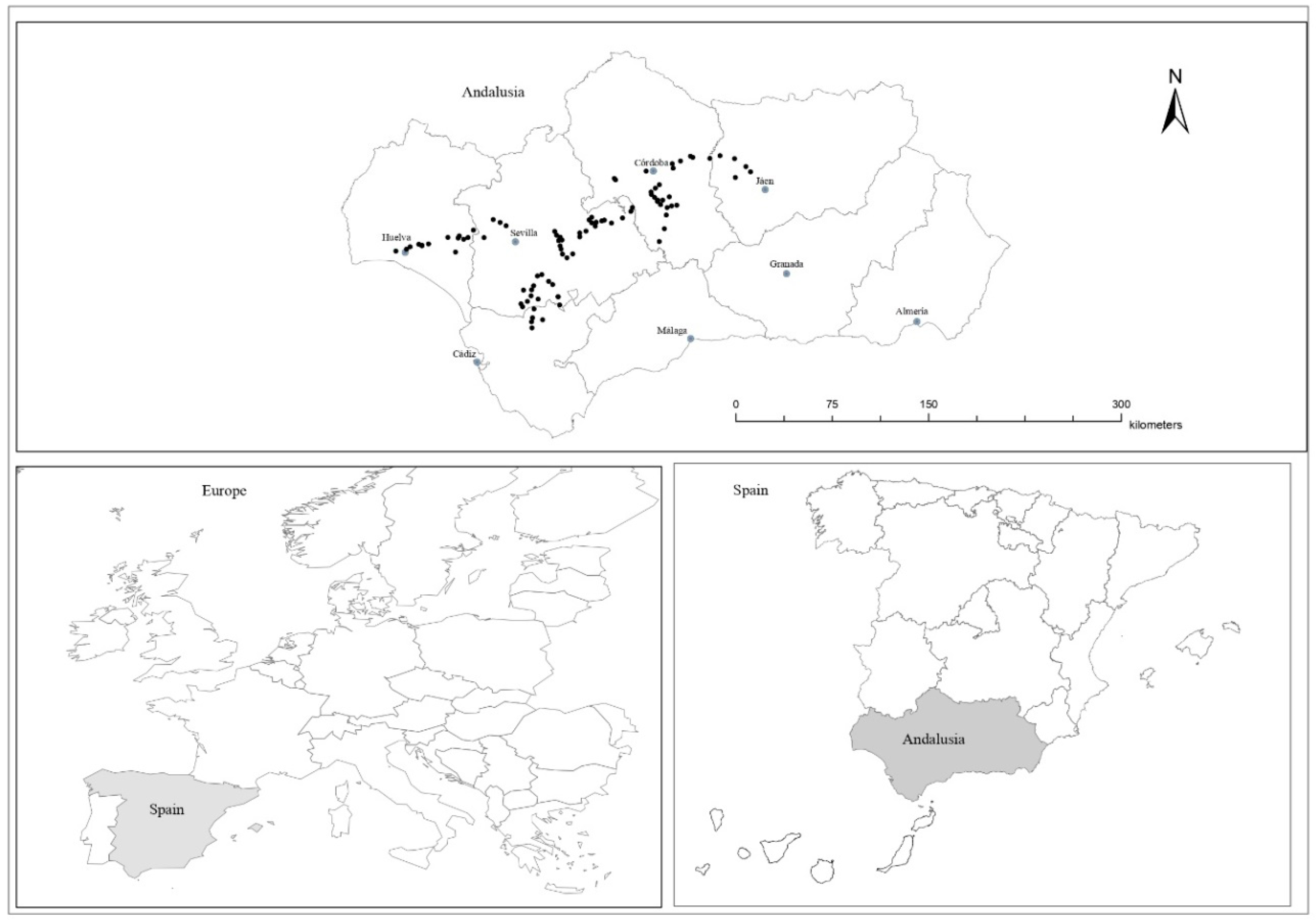
4.1. Study Area
4.2. Field Margin Selection
4.3. Margin Width and Landscape Features as Measures of Agricultural Intensification
4.4. Plant Surveys
4.5. Plant Functional Traits
4.6. Functional Diversity
4.7. Functional Diversity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Data Availability
Conflicts of Interest
Appendix A. List of the 306 Species Found in the 94 Field Margins and Their Frequency. The Species Are Sorted Alphabetically. In Bold, Are the 58 Species Considered in the RLQ, Fourth Corner and in the Functional Diversity Calculations
Appendix B. Species Traits (Q Table)
| Species | Life Form | Growth Form | Pollination | Seed Dispersal Type | Seed Mass (mg) |
| Anacyclus clavatus (Desf.) Pers. | therophyte | forb | entomogamy | anemochory | 0.50 |
| Anacyclus radiatus Loisel. | therophyte | forb | entomogamy | anemochory | 1.07 |
| Anagallis arvensis L. | therophyte | forb | entomogamy | barochory | 0.50 |
| Andryala integrifolia L. | hemicryptophyte | forb | entomogamy | anemochory | 0.15 |
| Avena sterilis L. | therophyte | grass | anemogamy | zoochory | 19.94 |
| Beta vulgaris L. | therophyte | forb | anemogamy | barochory | 12.70 |
| Bromus diandrus Roth | therophyte | grass | anemogamy | zoochory | 11.24 |
| Bromus lanceolatus Roth | therophyte | grass | anemogamy | zoochory | 3.90 |
| Bromus hordeaceus L. | therophyte | grass | anemogamy | zoochory | 1.48 |
| Bromus madritensis L. | therophyte | grass | anemogamy | zoochory | 3.33 |
| Calendula arvensis L. | therophyte | forb | entomogamy | zoochory | 5.20 |
| Campanula erinus L. | therophyte | forb | entomogamy | anemochory | 0.09 * |
| Centaurea melitensis L. | therophyte | forb | entomogamy | anemochory | 1.40 |
| Chenopodium vulvaria L. | therophyte | forb | anemogamy | barochory | 0.40 |
| Chrozophora tinctoria (L.) Raf. | therophyte | forb | entomogamy | barochory | 16.00 |
| Cinchorium intybus L. | hemicryptophyte | forb | entomogamy | anemochory | 5.50 |
| Convolvulus arvensis L. | geophyte | forb | entomogamy | barochory | 15.10 |
| Crepis vesicaria L. | hemicryptophyte | forb | entomogamy | anemochory | 0.36 |
| Cynodon dactylon (L.) Pers. | hemicryptophyte | grass | anemogamy | barochory | 0.20 |
| Daucus carota L. | hemicryptophyte | forb | entomogamy | zoochory | 1.00 |
| Diplotaxis virgata (Cav.) DC. | therophyte | forb | entomogamy | anemochory | 0.23 * |
| Ecballium elaterium (L.) A. Rich. | hemicryptophyte | forb | entomogamy | barochory | 12.10 |
| Echium plantagineum L. | therophyte | forb | entomogamy | barochory | 4.30 |
| Erodium malacoides (L.) L’Hér | therophyte | forb | entomogamy | barochory | 1.40 |
| Erodium moschatum (L.) L’Hér. | therophyte | forb | entomogamy | barochory | 2.62 |
| Euphorbia exigua L. | therophyte | forb | anemogamy | barochory | 0.35 |
| Galium aparine L. | therophyte | forb | entomogamy | zoochory | 8.70 |
| Galium parisiense L. | therophyte | forb | entomogamy | zoochory | 0.20 |
| Glebionis coronaria (L.) Spach | therophyte | forb | entomogamy | anemochory | 1.50 |
| Glebionis segetum (L.) Fourr. | therophyte | forb | entomogamy | anemochory | 1.52 |
| Heliotropium europaeum L. | therophyte | forb | entomogamy | barochory | 1.10 |
| Helminthotheca echioides (L.) Holub | hemicryptophyte | forb | entomogamy | anemochory | 1.31 |
| Hirschfeldia incana (L.) Lagr. Foss. | therophyte | forb | entomogamy | barochory | 0.23 |
| Hordeum murinum L. | therophyte | grass | anemogamy | zoochory | 10.50 |
| Lactuca serriola L. | therophyte | forb | autogamy | anemochory | 0.58 |
| Lavatera cretica L. | therophyte | forb | entomogamy | barochory | 7.01 |
| Lolium rigidum Gaudin | therophyte | grass | anemogamy | barochory | 3.34 |
| Malva nicaensis All. | therophyte | forb | entomogamy | barochory | 8.60 |
| Malva parviflora L. | therophyte | forb | entomogamy | barochory | 2.80 |
| Malva sylvestris L. | therophyte | forb | entomogamy | barochory | 5.40 |
| Medicago polymorpha L. | therophyte | forb | autogamy | zoochory | 2.95 |
| Papaver rhoeas L. | therophyte | forb | entomogamy | anemochory | 0.20 |
| Phalaris brachystachys Link | therophyte | grass | anemogamy | barochory | 1.90 |
| Phalaris minor Retz. | therophyte | grass | anemogamy | barochory | 1.60 |
| Phalaris paradoxa L. | therophyte | grass | anemogamy | barochory | 1.30 |
| Piptatherum miliaceum (L.) Coss. | hemicryptophyte | grass | anemogamy | barochory | 0.61 |
| Plantago lagopus L. | hemicryptophyte | forb | anemogamy | barochory | 0.30 |
| Polygonum aviculare L. | therophyte | forb | autogamy | barochory | 1.30 |
| Polypogon monspeliensis (L.) Desf. | therophyte | grass | anemogamy | barochory | 0.10 |
| Pulicaria paludosa Link | therophyte | forb | entomogamy | anemochory | 0.17 * |
| Rapistrum rugosum (L.) All. | therophyte | forb | entomogamy | barochory | 2.90 |
| Ridolfia segetum (L.) Moris | therophyte | forb | entomogamy | barochory | 0.60 |
| Scolymus maculatus L. | therophyte | forb | entomogamy | anemochory | 1.54 |
| Sonchus oleraceous L | therophyte | forb | entomogamy | anemochory | 0.30 |
| Silybum marianum (L.) Gaertn. | hemicryptophyte | forb | entomogamy | anemochory | 22.50 |
| Torilis arvensis (Huds.) Link | therophyte | forb | entomogamy | zoochory | 2.10 |
| Trisetaria panicea (Lam.) Paunero | therophyte | grass | anemogamy | barochory | 0.05 |
| Urospermum picrioides (L.) F. W. Schmidt | Therophyte | forb | entomogamy | anemochory | 1.6 |
| * The seed mass value was calculated by computing the average seed mass of congeneric species. | |||||
Appendix C. R-Scripts to Perform RLQ and Fourth-Corner Analyses
References
- Stoate, C.; Boatman, N.D.; Borralho, R.J.; Carvalho, C.R.; de Snoo, G.R.; Eden, P. Ecological impacts of arable intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef] [PubMed]
- Storkey, J.; Meyer, S.; Leuschner, C.; Still, K.S. The impact of agricultural intensification and land use change on the European arable flora. Proc. R. Soc. B Biol. Sci. 2012, 279, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Le Coeur, D.; Baudry, J.; Burel, F. Field margins plant assemblages: Variation partitioning between local and landscape factors. Landsc. Urban Plan. 1997, 37, 57–71. [Google Scholar] [CrossRef]
- Petit, S.; Stuart, R.C.; Gillespie, M.K.; Barr, C.J. Field boundaries in Great Britain: Stock and change between 1984, 1990 and 1998. J. Environ. Manag. 2003, 67, 229–238. [Google Scholar] [CrossRef]
- Baessler, C.; Klotz, S. Effects of changes in agricultural land-use on landscape structure and arable weed vegetation over the last 50 years. Agric. Ecosyst. Environ. 2006, 115, 43–50. [Google Scholar] [CrossRef]
- Ponisio, L.C.; M’Gonigle, L.K.; Kremen, C. On-farm habitat restoration counters biotic homogenization in intensively managed agriculture. Glob. Chang. Biol. 2016, 22, 704–715. [Google Scholar] [CrossRef] [Green Version]
- Morrison, J.; Hernández Plaza, E.; Izquierdo, J.; Gonzalez-Andujar, J.L. The role of field margins in supporting wild bees in Mediterranean cereal agroecosystems: Which biotic and abiotic factors are important? Agric. Ecosyst. Environ. 2017, 247, 216–224. [Google Scholar] [CrossRef]
- Cirujeda, A.; Pardo, G.; Marí, A.I.; Aibar, J.; Pallavicini, Y.; Gonzalez-Andujar, J.L.; Recasens, J.; Sole-Senan, X.O. The structural classification of field boundaries in Mediterranean arable cropping systems allows the prediction of weed abundances in the boundary and in the adjacent crop. Weed Res. 2019, 59, 300–311. [Google Scholar] [CrossRef]
- Sanchez, J.A.; Carrasco, A.; La Spina, M.; Pérez-Marcos, M.; Ortiz-Sánchez, F.J. How Bees Respond Differently to Field Margins of Shrubby and Herbaceous Plants in Intensive Agricultural Crops of the Mediterranean Area. Insects 2020, 11, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, E.J.P.; Moonen, A.C. Field margins in northern Europe: Their functions and interactions with agriculture. Agric. Ecosyst. Environ. 2002, 89, 5–21. [Google Scholar] [CrossRef]
- Cordeau, S.; Petit, S.; Reboud, X.; Chauvel, B. The impact of sown grass strips on the spatial distribution of weed species in adjacent boundaries and arable fields. Agric. Ecosyst. Environ. 2012, 155, 35–40. [Google Scholar] [CrossRef]
- Vickery, J.A.; Feber, R.E.; Fuller, R.J. Arable field margins managed for biodiversity conservation: A review of food resource provision for farmland birds. Agric. Ecosyst. Environ. 2009, 133, 1–13. [Google Scholar] [CrossRef]
- Poggio, S.L.; Chaneton, E.J.; Ghersa, C.M. Landscape complexity differentially affects alpha, beta, and gamma diversities of plants occurring in fencerows and crop fields. Biol. Conserv. 2010, 143, 2477–2486. [Google Scholar] [CrossRef]
- Bassa, M.; Chamorro, L.; José-María, L.; Blanco-Moreno, J.; Sans, F. Factors affecting plant species richness in field boundaries in the Mediterranean region. Biodivers. Conserv. 2012, 21, 1101–1114. [Google Scholar] [CrossRef]
- Schippers, P.; Joenje, W. Modelling the effect of fertiliser, mowing, disturbance and width on the biodiversity of plant communities of field boundaries. Agric. Ecosyst. Environ. 2002, 93, 351–365. [Google Scholar] [CrossRef]
- Tarmi, S.; Helenius, J.; Hyvönen, T. Importance of edaphic, spatial and management factors for plant communities of field boundaries. Agric. Ecosyst. Environ. 2009, 131, 201–206. [Google Scholar] [CrossRef]
- Marshall, E.J.P. The impact of landscape structure and sown grass margin strips on weed assemblages in arable crops and their boundaries. Weed Res. 2009, 49, 107–115. [Google Scholar] [CrossRef]
- José-María, L.; Armengot, L.; Blanco-Moreno, J.M.; Bassa, M.; Sans, F.X. Effects of agricultural intensification on plant diversity in Mediterranean dryland cereal fields. J. Appl. Ecol. 2010, 47, 832–840. [Google Scholar] [CrossRef]
- Jonason, D.; Andersson, G.K.S.; Öckinger, E.; Rundlöf, M.; Smith, H.G.; Bengtsson, J. Assessing the effect of the time since transition to organic farming on plants and butterflies. J. Appl. Ecol. 2011, 48, 543–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aparicio, A. Descriptive analysis of the ‘relictual’ Mediterranean landscape in the Guadalquivir River valley (southern Spain): A baseline for scientific research and the development of conservation action plans. Biodivers. Conserv. 2008, 17, 2219–2232. [Google Scholar] [CrossRef]
- Rodríguez, C.; Wiegand, K. Evaluating the trade-off between machinery efficiency and loss of biodiversity-friendly habitats in arable landscapes: The role of field size. Agric. Ecosyst. Environ. 2009, 129, 361–366. [Google Scholar] [CrossRef]
- Flynn, D.B.; Gogol-Prokurat, M.; Nogeire, T.; Molinari, N.; Trautman Richers, B.; Lin, B.B.; Simpson, N.; Mayfield, M.M.; DeClerck, F. Loss of functional diversity under land use intensification across multiple taxa. Ecol. Lett. 2009, 12, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Keddy, P.A. Assembly and response rules: Two goals for predictive community ecology. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Diaz, S.; Cabido, M.; Casanoves, F. Plant functional traits and environmental filters at a regional scale. J. Veg. Sci. 1998, 9, 113–122. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- José-María, L.; Blanco-Moreno, J.M.; Armengot, L.; Sans, F.X. How does agricultural intensification modulate changes in plant community composition? Agric. Ecosyst. Environ. 2011, 145, 77–84. [Google Scholar] [CrossRef]
- Fried, G.; Kazakou, E.; Gaba, S. Trajectories of weed communities explained by traits associated with species’ response to management practices. Agric. Ecosyst. Environ. 2012, 158, 147–155. [Google Scholar] [CrossRef]
- Ma, M.; Herzon, I. Plant functional diversity in agricultural margins and fallow fields varies with landscape complexity level: Conservation implications. J. Nat. Conserv. 2014, 26, 525–531. [Google Scholar] [CrossRef]
- Moreno, J.C. Lista Roja 2008 de la Flora Vascular Española; Dirección General de Medio Natural y Política Forestal (Ministerio de Medio Ambiente, y Medio Rural y Marino, y Sociedad Española de Biología de la conservación de Plantas): Madrid, Spain, 2008.
- Benvenuti, S. Weed seed movement and dispersal strategies in the agricultural environment. Weed Biol. Manag. 2007, 7, 141–157. [Google Scholar] [CrossRef]
- Blanca, G.; Cabezudo, B.; Cueto, M.; Morales-Torres, C.; Salazar, C. Flora Vascular de Andalucía Oriental, 2nd ed.; Universidades de Almería: Jaén y Málaga, Spain, 2011. [Google Scholar]
- Julve, P. Baseflor. Index Botanique Écologique et Chorologique de la Flore de France; Institut Catholique de Lille: Lille, France, 1998; Available online: http://perso.wanadoo.fr/philippe.julve/catminat.htm (accessed on 1 April 2019).
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; Van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA Traitbase: A database of life-history traits of the Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Royal Botanic Gardens. Seed Information Database (SID), Version 7.1; Royal Botanic Gardens: Kew, UK, 2008. [Google Scholar]
- Ma, M.; Tarmi, S.; Helenius, J. Revisiting the species–area relationship in a semi-natural habitat: Floral richness in agricultural buffer zones in Finland. Agric. Ecosyst. Environ. 2002, 89, 137–148. [Google Scholar] [CrossRef]
- Schmitz, J.; Schäfer, K.; Brühl, C.A. Agrochemicals in field margins—Field evaluation of plant reproduction effects. Agric. Ecosyst. Environ. 2014, 189, 82–91. [Google Scholar] [CrossRef]
- Gonzalez-Andujar, J.L.; Saavedra, M. Spatial distribution of annual grass weed populations in winter cereals. Crop Prot. 2003, 22, 629–633. [Google Scholar] [CrossRef]
- Dainese, M.; Montecchiari, S.; Sitzia, T.; Sigura, M.; Marini, L. High cover of hedgerows in the landscape supports multiple ecosystem services in Mediterranean cereal fields. J. Appl. Ecol. 2017, 54, 380–388. [Google Scholar] [CrossRef] [Green Version]
- Weibull, A.C.; Östman, Ö.; Granqvist, Å. Species richness in agroecosystems: The effect of landscape, habitat and farm management. Biodivers. Conserv. 2003, 12, 1335–1355. [Google Scholar] [CrossRef]
- Lososová, Z.; Chytrý, M.; Kühn, I.; Hájek, O.; Horáková, V.; Pyšek, P.; Tichý, L. Patterns of plant traits in annual vegetation of man-made habitats in central Europe. Perspect. Plant Ecol. 2006, 8, 69–81. [Google Scholar] [CrossRef]
- de Andalucía, J. Anuario de Estadísticas Agrarias y Pesqueras en Andalucía; Consejería de Agricultura, Pesca y Desarrollo Rural: Sevilla, Spain, 2013.
- Roschewitz, I.; Thies, C.; Tscharntke, T. Are landscape complexity and farm specialisation related to land-use intensity of annual crop fields? Agric. Ecosyst. Environ. 2005, 105, 87–99. [Google Scholar] [CrossRef]
- McIntyre, S.; Lavorel, S.; Tremont, R.M. Plant life-history attributes: Their relationship to disturbance response in herbaceous vegetation. J. Ecol. 1995, 83, 31–44. [Google Scholar] [CrossRef]
- Hawes, C.; Squire, G.R.; Hallett, P.D.; Watson, C.A.; Young, M. Arable plant communities as indicators of farming practice. Agric. Ecosyst. Environ. 2010, 138, 17–26. [Google Scholar] [CrossRef]
- Holzschuh, A.; Steffan-Dewenter, I.; Kleijn, D.; Tscharntke, T. Diversity of flower-visiting bees in cereal fields: Effects of farming system, landscape composition and regional context. J. Appl. Ecol. 2007, 44, 41–49. [Google Scholar] [CrossRef]
- Petit, S.; Alignier, A.; Colbach, N.; Joannon, A.; Cœur, D.; Thenail, C. Weed dispersal by farming at various spatial scales. A review. Agron. Sustain. Dev. 2012, 1–13. [Google Scholar] [CrossRef]
- Leishman, M.R. Does the seed size/number trade-off model determine plant community structure? An assessment of the model mechanisms and their generality. Oikos 2001, 93, 294–302. [Google Scholar] [CrossRef]
- Pakeman, R.J.; Garnier, E.; Lavorel, S.; Ansquer, P.; Castro, H.; Cruz, P.; Doležal, J.; Eriksson, O.; Freitas, H.; Golodets, C.; et al. Impact of abundance weighting on the response of seed traits to climate and land use. J. Ecol. 2008, 96, 355–366. [Google Scholar] [CrossRef]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; Wiley: New York, NY, USA, 1974. [Google Scholar]
- Kenkel, N.C.; Derksen, D.A.; Thomas, A.G.; Watson, P.R. Review: Multivariate analysis in weed science research. Weed Sci. 2002, 50, 281–292. [Google Scholar] [CrossRef]
- Rao, C.R. Diversity and dissimilarity coefficients: A unified approach. Theor. Popul. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.H.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Mason, N.W.; Bello, F.; Mouillot, D.; Pavoine, S.; Dray, S. A guide for using functional diversity indices to reveal changes in assembly processes along ecological gradients. J. Veg. Sci. 2013, 24, 794–806. [Google Scholar] [CrossRef]
- Hill, M.O.; Smith, A.J.E. Principal Component Analysis of Taxonomic Data with Multi-State Discrete Characters. Taxon 1976, 25, 249–255. [Google Scholar] [CrossRef]
- Dray, S.; Choler, P.; Doledec, S.; Peres-Neto, P.R.; Thuiller, W.; Pavoine, S.; ter Braak, C.J. Combining the Fourth-corner and the RLQ methods for assessing trait responses to environmental variation. Ecology 2014, 95, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Dolédec, S.; Chessel, D.; Braak, C.J.F.; Champely, S. Matching species traits to environmental variables: A new three-table ordination method. Environ. Ecol. Stat. 1996, 3, 143–166. [Google Scholar] [CrossRef]
- Legendre, P.; Galzin, R.; Harmelin-Vivien, M.L. Relating behavior to habitat: Solutions to the Fourth-corner problem. Ecology 1997, 78, 547–562. [Google Scholar] [CrossRef]
- Dray, S.; Legendre, P. Testing the species traits-environment relationships: The fourth-corner problem revisited. Ecology 2008, 89, 3400–3412. [Google Scholar] [CrossRef] [PubMed]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Dray, S.; Dufour, A.B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Harrell, F.; Dupont, C. Hmisc: Harrell Miscellaneous. R Package Version 3.14-0. 2014. Available online: http://CRAN.R-project.org/package=Hmisc (accessed on 10 December 2019).
- Giradoux, P. Pgirmess: Data Analysis in Ecology. R. Package Version 1.5.8. 2013. Available online: http://CRAN.R-project.org/package=pgirmess (accessed on 18 December 2019).




{kind=link}
{kind=link}
{kind=link}
| Traits | Category | Abbreviation | Number of Species | Min. | Max. | Mean ± SD | Source |
|---|---|---|---|---|---|---|---|
| Raunkiær’s life forms | Geophytes | geop | 1 | - | - | - | [33] |
| Hemicryptophytes | hemi | 10 | - | - | - | [33] | |
| Therophytes | thero | 47 | - | - | - | [33] | |
| Growth form | Forbs | forb | 44 | - | - | - | [33] |
| Grasses | gras | 14 | - | - | - | [33] | |
| Pollination | Anemogamy | aneg | 18 | - | - | - | [34] |
| Autogamy | auto | 3 | - | - | - | [34] | |
| Entomogamy | ento | 37 | - | - | - | [34] | |
| Seed dispersal type | Anemochory | anem | 18 | - | - | - | [33,35] |
| Barochory | baro | 28 | - | - | - | [33,35] | |
| Zoochory | zooc | 12 | - | - | - | [33,35] | |
| Seed mass (mg) | - | sm | 58 | 0.05 | 22.5 | 3.70 ± 5.20 | [36] |
| ρ | χ2 | p-Value | |
|---|---|---|---|
| Arable land | 0.84 | <0.001 | |
| Field Size | 0.28 | <0.004 | |
| Shannon index for land use diversity | −0.58 | <0.001 | |
| Woodland cover | −0.33 | 0.001 | |
| Grassland cover | −0.27 | 0.007 | |
| Cover of human settlements | −0.58 | 0.002 | |
| Margin width class | F = 2.03 | <0.001 |
| PCA1 | Arable Land Cover | Field Size | Shannon Index for Land Use Diversity | Woodland Cover | Grassland Cover | Human Settlements Cover | Margin Width Class | |
|---|---|---|---|---|---|---|---|---|
| Species richness | ρ = 0.06 | ρ = 0.09 | ρ = 0.17 | ρ = −0.11 | ρ = −0.04 | ρ = 0.09 | ρ = 0.05 | χ2 = 31.70 |
| FDT | ρ = 0.03 | ρ = 0.08 | ρ = −0.07 | ρ = −0.10 | ρ = 0.00 | ρ = −0.03 | ρ = 0.09 | χ2 = 6.45 |
| FDS-Raunkiær’s life form | ρ = 0.07 | ρ = 0.01 | ρ = −0.02 | ρ = 0.02 | ρ = 0.00 | ρ = −0.01 | ρ = 0.19 | χ2 = 2.74 |
| FDS-growth form | ρ = 0.04 | ρ = 0.06 | ρ = −0.04 | ρ = 0.02 | ρ = 0.05 | ρ = 0.03 | ρ = −0.01 | χ2 = 0.36 |
| FDS-pollination type | ρ = 0.14 | ρ = −0.15 | ρ = −0.17 | ρ = 0.02 | ρ = 0.13 | ρ = −0.03 | ρ = −0.07 | χ2 = 2.43 |
| FDS-seed dispersal mode | ρ = 0.07 | ρ = 0.05 | ρ = 0.05 | ρ = 0.03 | ρ = 0.03 | ρ = −0.11 | ρ = −0.06 | χ2 = 16.50 |
| FDS-seed mass | ρ = 0.02 | ρ = 0.03 | ρ = 0.00 | ρ = −0.04 | ρ = −0.07 | ρ = −0.02 | ρ = 0.07 | χ2 = 2.29 |
| Arable Land Cover | Grassland Cover | Woodland Cover | Human Settlements Cover | Shannon Index for Land Use Diversity | Field Size | Margin Width Class | |
|---|---|---|---|---|---|---|---|
| Raunkiær’s life forms | F = 0.40 | F = 0.32 | F = 0.90 | F = 0.23 | F = 0.00 | F = 2.37 | χ2 = 4.61 |
| Growth form | F = 0.50 | F = 0.87 | F = 0.46 | F = 0.25 | F = 2.86 | F = 0.13 | χ2 = 1.61 |
| Pollination | F = 1.26 | F = 0.19 | F = 0.25 | F = 0.25 | F = 1.99 | F = 0.43 | χ2 = 3.24 |
| Seed dispersal type | F = 1.26 | F = 0.92 | F = 1.91 | F = 0.55 | F = 0.42 | F = 0.11 | χ2 = 6.52 |
| Seed mass | r = −0.01 | r = −0.01 | r = 0.01 | r = 0.00 | r = 0.00 | r = −0.1 | F = 0.55 |
| Landscape Variables | Abbreviation | Category | Min. | Max. | Mean ± SD | Frequency |
|---|---|---|---|---|---|---|
| Arable land cover (%) | AL | 2.00 | 100 | 73.8 ± 29.90 | 94 | |
| Field Size (ha) | FS | 0.16 | 281.00 | 9.0 ± 49.10 | 94 | |
| Shannon habitat diversity index | SHDI | 0.00 | 1.10 | 0.5 ± 0.30 | 94 | |
| Forest (%) | FO | 0.00 | 30.00 | 0.9 ± 3.70 | 13 | |
| Grassland (%) | GR | 0.00 | 47.00 | 0.9 ± 5.10 | 12 | |
| Human settlements (%) | HS | 0.00 | 40.00 | 4.0 ± 6.10 | 74 | |
| Margin width | width | Narrow | 0 m | 0.99 m | - | 37 |
| Medium | 1 | 1.99 m | - | 26 | ||
| Wide | >2 m | - | 31 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pallavicini, Y.; Bastida, F.; Hernández-Plaza, E.; Petit, S.; Izquierdo, J.; Gonzalez-Andujar, J.L. Local Factors Rather than the Landscape Context Explain Species Richness and Functional Trait Diversity and Responses of Plant Assemblages of Mediterranean Cereal Field Margins. Plants 2020, 9, 778. https://doi.org/10.3390/plants9060778
Pallavicini Y, Bastida F, Hernández-Plaza E, Petit S, Izquierdo J, Gonzalez-Andujar JL. Local Factors Rather than the Landscape Context Explain Species Richness and Functional Trait Diversity and Responses of Plant Assemblages of Mediterranean Cereal Field Margins. Plants. 2020; 9(6):778. https://doi.org/10.3390/plants9060778
Chicago/Turabian StylePallavicini, Yesica, Fernando Bastida, Eva Hernández-Plaza, Sandrine Petit, Jordi Izquierdo, and Jose L. Gonzalez-Andujar. 2020. "Local Factors Rather than the Landscape Context Explain Species Richness and Functional Trait Diversity and Responses of Plant Assemblages of Mediterranean Cereal Field Margins" Plants 9, no. 6: 778. https://doi.org/10.3390/plants9060778
APA StylePallavicini, Y., Bastida, F., Hernández-Plaza, E., Petit, S., Izquierdo, J., & Gonzalez-Andujar, J. L. (2020). Local Factors Rather than the Landscape Context Explain Species Richness and Functional Trait Diversity and Responses of Plant Assemblages of Mediterranean Cereal Field Margins. Plants, 9(6), 778. https://doi.org/10.3390/plants9060778