Artifleck: The Study of Artifactual Responses to Light Flecks with Inappropriate Leaves


Abstract
:
1. Introduction
2. Results
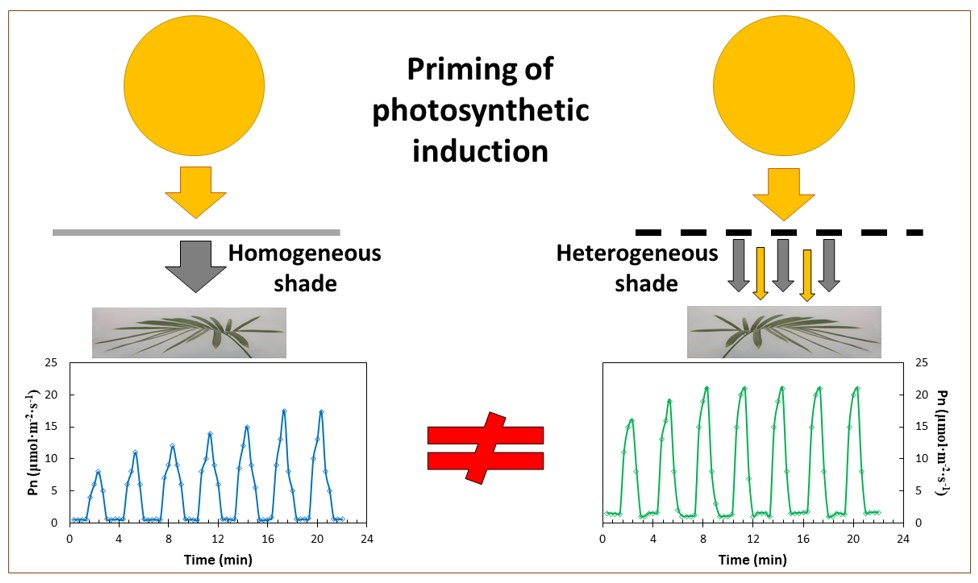
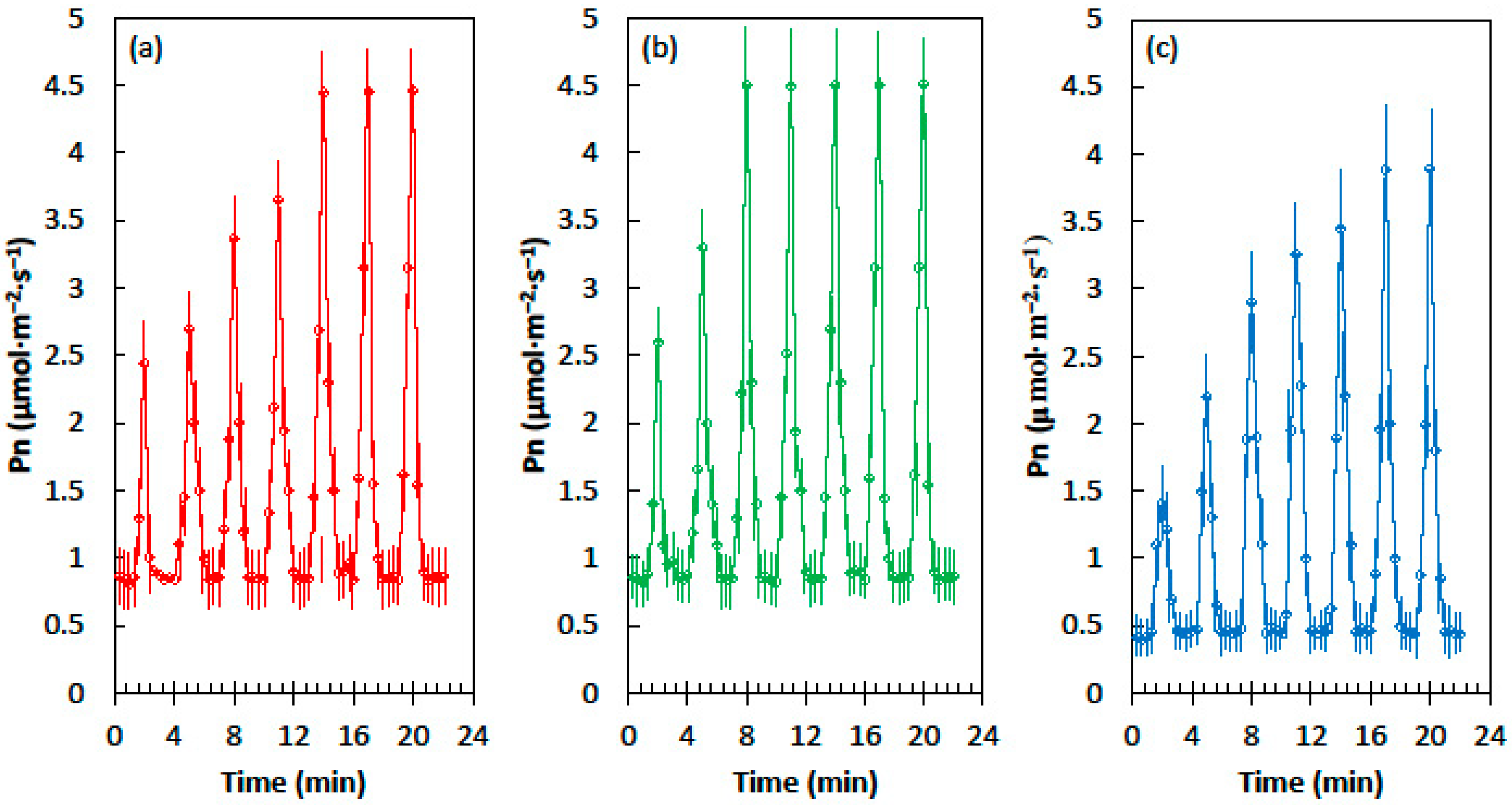
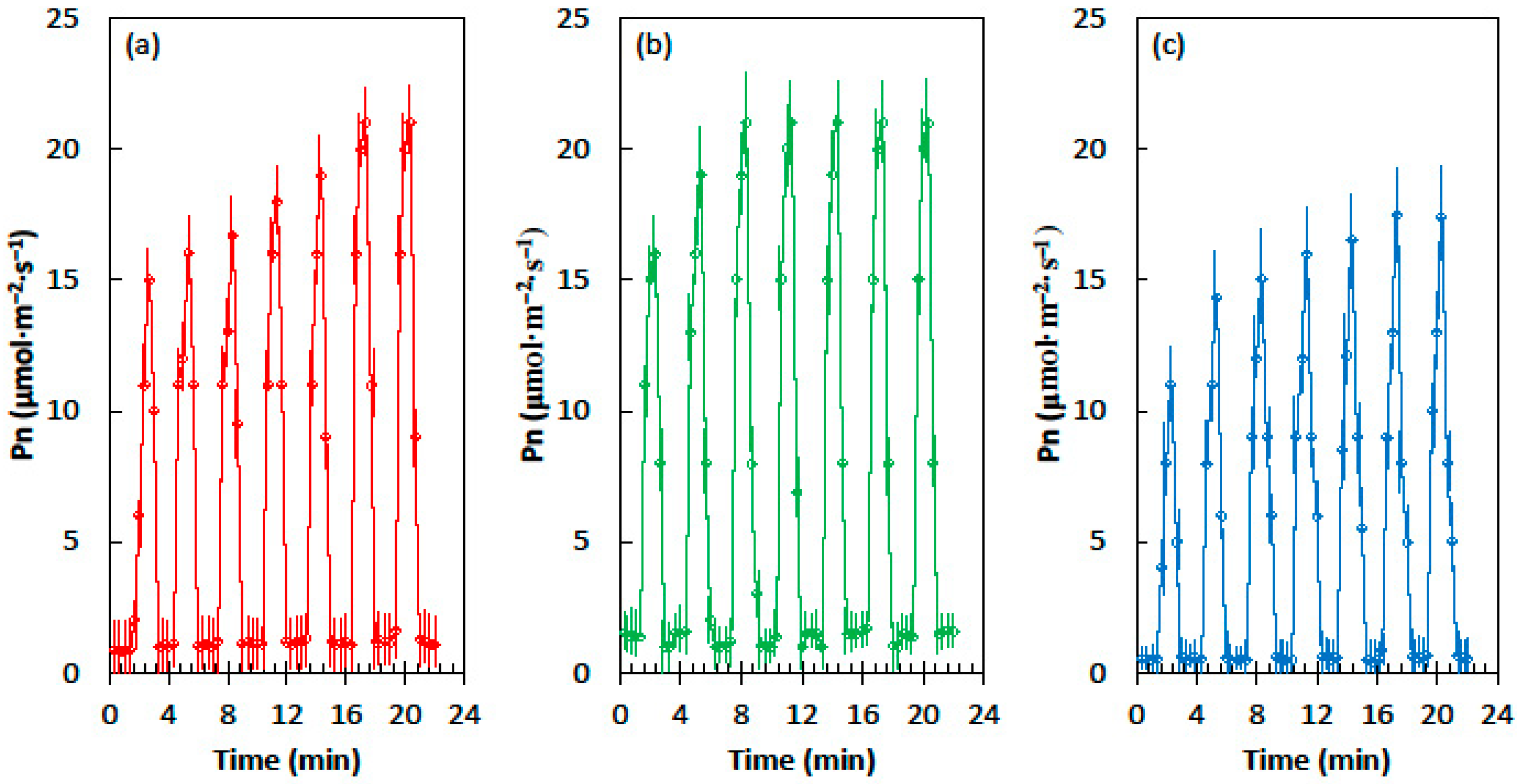
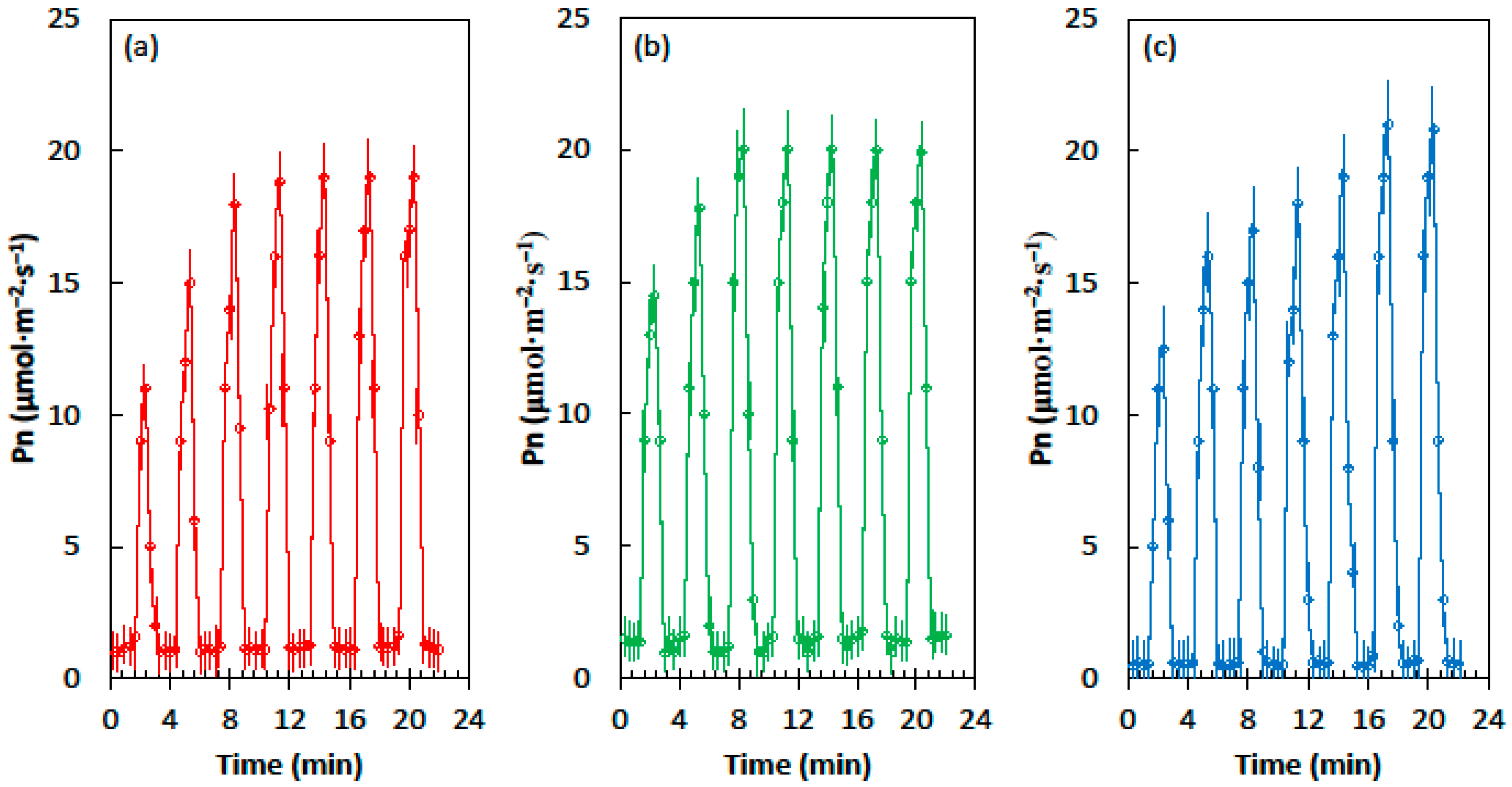
2.1. Light Flecks
2.2. Photosynthetic Induction
2.3. Leaf Nutrient Relations
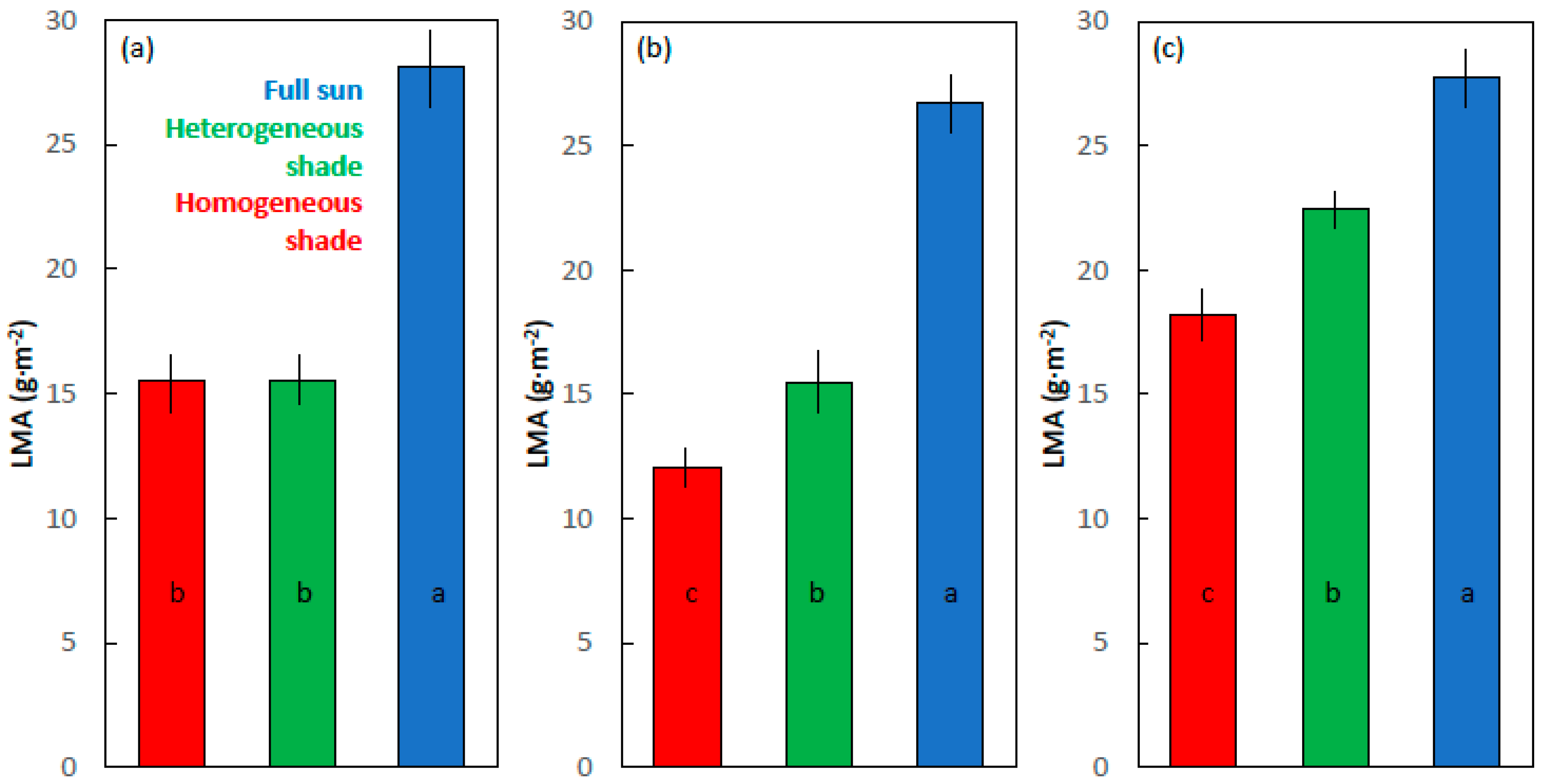
2.4. Leaf Mass per Area
3. Discussion
3.1. Looking Backward, Looking Forward
3.2. Complications
4. Materials and Methods
4.1. Experimental Conditions
4.2. Physiology Measurements
4.3. Leaf Mass per Area and Nutrient Relations
4.4. Calculations and Statistics
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Walker, D.A. Photosynthetic induction phenomena and the light activation of ribulose diphosphate carboxylase. New Phytol. 1972, 72, 209–235. [Google Scholar] [CrossRef]
- Pearcy, R.W. Sunflecks and photosynthesis in plant canopies. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 421–453. [Google Scholar] [CrossRef]
- Burgess, A.J.; Retkute, R.; Preston, S.P.; Jensen, O.E.; Pound, M.P.; Pridmore, T.P.; Murchie, E.H. The 4-dimensional plant: Effects of wind-induced canopy movement on light fluctuations and photosynthesis. Front. Plant Sci. 2016, 7, e1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, E.; Morales, A.; Harbinson, J. Fluctuating light takes crop photosynthesis on a rollercoaster ride. Plant Physiol. 2018, 176, 977–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, A.; Kaiser, E. Photosynthetic acclimation to fluctuating irradiance in plants. Front. Plant Sci. 2020, 11, 268. [Google Scholar] [CrossRef] [PubMed]
- Soleh, M.A.; Tanaka, Y.; Nomoto, Y.; Iwahashi, Y.; Nakashima, K.; Fukuda, Y.; Long, S.P.; Shiraiwa, T. Factors underlying genotypic differences in the induction of photosynthesis in soybean [Glycine max (L.) Merr.]. Plant Cell Environ. 2016, 39, 685–693. [Google Scholar] [CrossRef]
- Acevedo-Siaca, L.G.; Coe, R.; Wang, Y.; Kromdijk, J.; Quick, W.P.; Long, S.P. Variation in photosynthetic induction between rice accessions and its potential for improving productivity. New Phytol. 2020. [CrossRef] [Green Version]
- Wang, Y.; Burgess, S.J.; de Becker, E.M.; Long, S.P. Photosynthesis in the fleeting shadows: An overlooked opportunity for increasing crop productivity? Plant J. 2020, 101, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Allee, W.C. Measurement of environmental factors in the tropical rain forest of Panama. Ecology 1926, 7, 273–302. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Way, D.A. Two decades of sunfleck research: Looking back to move forward. Tree Physiol. 2012, 32, 1059–1061. [Google Scholar] [CrossRef] [Green Version]
- Serna-Saldivar, S.O. (Ed.) Corn, 3rd ed.; AACC International Press: Washington, DC, USA, 2019; pp. 435–467. [Google Scholar]
- Shea, Z.; Singer, W.M.; Zhang, B. Soybean Production, Versatility, and Improvement. In Legume Crops; Hasanuzaman, M., Ed.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Marler, T.E. Leaf physiology of shade-grown Cycas micronesica leaves following removal of shade. Bot. Rev. 2004, 70, 63–71. [Google Scholar] [CrossRef]
- Marler, T.E. Age influences photosynthetic capacity of Cycas micronesica leaves. Mem. N. Y. Bot. Garden 2007, 97, 193–203. [Google Scholar]
- Furbank, R.T.; Leegood, R.C. Carbon metabolism and gas exchange in leaves of Zea mays L. Planta 1984, 162, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Krall, J.P.; Pearcy, R.W. Concurrent measurements of oxygen and carbon dioxide exchange during lightflecks in maize (Zea mays L.). Plant Physiol. 1993, 103, 823–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Yang, Z.; Zhou, P.; Hai, M.-R.; Tang, T.-X.; Liang, Y.-L.; An, T.-X. Biomass accumulation and partitioning, photosynthesis, and photosynthetic induction in field-grown maize (Zea mays L.) under low- and high-nitrogen conditions. Acta Physiol. Plant 2013, 35, 95–105. [Google Scholar] [CrossRef]
- McAusland, L.; Vialet-Chabrand, S.; Davey, P.; Baker, N.R.; Brendel, O.; Lawson, T. Effects of kinetics of light-induced stomatal responses on photosynthesis and water-use efficiency. New Phytol. 2016, 211, 1209–1220. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Roden, J.S.; Gamon, J.A. Sunfleck dynamics in relation to canopy structure in a soybean (Glycine max (L.) Merr.) canopy. Agric. Forest Meteor. 1990, 52, 359–372. [Google Scholar] [CrossRef]
- Pons, T.L.; Pearcy, R.W. Photosynthesis in flashing light in soybean leaves grown in different conditions. II. Lightfleck utilization efficiency. Plant Cell Environ. 1992, 15, 577–584. [Google Scholar] [CrossRef]
- Pons, T.L.; Pearcy, R.W.; Seemann, J.R. Photosynthesis in flashing light in soybean leaves grown in different conditions. I. Photosynthetic induction state and regulation of ribulose-1,5-bisphosphate carboxylase activity. Plant Cell Environ. 1992, 15, 569–576. [Google Scholar] [CrossRef]
- Sassenrath-Cole, G.F.; Pearcy, R.W.; Steinmaus, S. The role of enzyme activation state in limiting carbon assimilation under variable light conditions. Photosynth. Res. 1994, 41, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Pearcy, R.W.; Gross, L.J.; He, D. An improved dynamic model of photosynthesis for estimation of carbon gain in sunfleck light regimes. Plant Cell Environ. 1997, 20, 411–424. [Google Scholar] [CrossRef]
- Pieruschka, R.; Chavarría-Krauser, A.; Schurr, U.; Jahnke, S. Photosynthesis in lightfleck areas of homobaric and heterobaric leaves. J. Exp. Bot. 2010, 61, 1031–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soleh, M.A.; Tanaka, Y.; Kim, S.Y.; Huber, S.C.; Sakoda, K.; Shiraiwa, T. Identification of large variation in the photosynthetic induction response among 37 soybean [Glycine max (L.) Merr.] genotypes that is not correlated with steady-state photosynthetic capacity. Photosynth. Res. 2017, 131, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E. Bi-directional acclimation of Cycas micronesica leaves to abrupt changes in incident light in understory and open habitats. Photosynthetica 2017, 56, 776–785. [Google Scholar] [CrossRef]
- Sims, D.A.; Pearcy, R.W. Sunfleck frequency and duration affects growth rate of the understorey plant, Alocasia macrorrhiza. Func. Ecol. 1993, 7, 683–689. [Google Scholar] [CrossRef]
- Watling, J.R.; Ball, M.C.; Woodrow, I.E. The Utilization of lightflecks for growth in four Australian rain-forest species. Func. Ecol. 1997, 11, 231–239. [Google Scholar] [CrossRef]
- Rey-Sanchez, C.; Posada, J.M. Effect of temporally heterogeneous light on photosynthetic light use efficiency, plant acclimation and growth in Abatia parviflora. Func. Plant Biol. 2019, 46, 684–693. [Google Scholar] [CrossRef] [Green Version]
- Kursar, T.A.; Coley, P.D. Photosynthetic induction times in shade-tolerant species with long and short-lived leaves. Oecologia 1993, 93, 165–170. [Google Scholar] [CrossRef]
- Boonstra, R. The ecology of stress: A marriage of disciplines. Func. Ecol. 2013, 27, 7–10. [Google Scholar] [CrossRef]
- Knapp, A.K.; Smith, W.K. Contrasting stomatal responses to variable sunlight in two subalpine herbs. Am. J. Bot. 1990, 77, 226–231. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, Q.; Tabassum, M.A.; Ye, M.; Peng, S.; Li, Y. The inhibition of photosynthesis under water deficit conditions is more severe in flecked than uniform irradiance in rice (Oryza sativa) plants. Func. Plant Biol. 2017, 44, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Ye, M.; Peng, S.; Lim, Y. Nitrogen can improve the rapid response of photosynthesis to changing irradiance in rice (Oryza sativa L.) plants. Sci. Rep. 2016, 6, 31305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Kaiser, E.; Marcelis, L.F.; Yang, Q.; Li, T. Salt stress and fluctuating light have separate effects on photosynthetic acclimation, but interactively affect biomass. Plant Cell Environ. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Leakey, A.D.B.; Press, M.C.; Scholes, J.D.; Watling, J.R. Relative enhancement of photosynthesis and growth at elevated CO2 is greater under sunflecks than uniform irradiance in a tropical rain forest tree seedling. Plant Cell Environ. 2002, 25, 1701–1714. [Google Scholar] [CrossRef]
- Smith, H. Phytochromes and light signal perception by plants—An emerging synthesis. Nature 2000, 407, 585–591. [Google Scholar] [CrossRef]
- Casal, J.J. Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol. 2013, 64, 403–427. [Google Scholar] [CrossRef]
- Ballaré, C.L.; Pierik, R. The shade-avoidance syndrome: Multiple signals and ecological consequences. Plant Cell Environ. 2017, 40, 2530–2543. [Google Scholar] [CrossRef]
- Kono, M.; Kawaguchi, H.; Mizusawa, N.; Yamori, W.; Suzuki, Y.; Terashima, I. Far-red light accelerates photosynthesis in the low-light phases of fluctuating light. Plant Cell Physiol. 2020, 61, 192–202. [Google Scholar] [CrossRef]
- Zellweger, F.; Baltensweiler, A.; Schleppi, P.; Huber, M.; Küchler, M.; Ginzler, C.; Jonas, T. Estimating below-canopy light regimes using airborne laser scanning: An application to plant community analysis. Ecol. Evol. 2019, 9, 9149–9159. [Google Scholar] [CrossRef] [Green Version]
- Smith, W.K.; Knapp, A.K.; Reiners, W.A. Penumbral effects on sunlight penetration in plant communities. Ecology 1989, 70, 1603–1609. [Google Scholar] [CrossRef]
- Dumas, J.B.A. Procedes de L’analyse Organique. Ann. Chim. Phys. 1831, 47, 198–205. [Google Scholar]
- Hou, X.; Jones, B.T. Inductively coupled plasma/optical emission spectrometry. In Encyclopedia of Analytical Chemistry; Meyers, R.A., Ed.; John Wiley & Sons: Chichester, UK, 2000; pp. 9468–9485. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Homogeneous Shade | Heterogeneous Shade | Full Sun | F2,9 | p |
|---|---|---|---|---|---|
| Cycas micronesica | |||||
| Coefficient a | 1.87 ± 0.21a | 1.93 ± 0.22a | 0.96 ± 0.21b | 6.881 | 0.015 |
| Coefficient b | −0.10 ± 0.02b | −0.09 ± 0.03b | 0.20 ± 0.06a | 21.223 | 0.001 |
| Coefficient c | 2.85 ± 0.08a | 3.07 ± 0.21a | 2.05 ± 0.11b | 7.144 | 0.014 |
| Amax (µmol·m−2·s−1) | 8.50 ± 0.18c | 9.63 ± 0.23b | 11.25 ± 0.27a | 47.860 | <0.001 |
| IT 50% (min) | 3.23 ± 0.46b | 3.25 ± 0.43b | 6.13 ± 0.51a | 15.171 | 0.001 |
| IT 90% (min) | 9.63 ± 0.72b | 7.75 ± 0.46c | 13.13 ± 0.81a | 16.160 | 0.001 |
| Glycine max | |||||
| Coefficient a | 8.90 ± 0.23b | 10.36 ± 0.22a | 5.50 ± 0.37c | 57.551 | <0.001 |
| Coefficient b | −0.01 ± 0.06b | −0.29 ± 0.06c | 0.66 ± 0.08a | 56.685 | <0.001 |
| Coefficient c | 4.05 ± 0.37b | 7.00 ± 0.40a | 3.73 ± 0.31b | 24.762 | 0.001 |
| Amax (µmol·m−2·s−1) | 21.01 ± 0.61c | 25.75 ± 0.63b | 30.50 ± 0.87a | 56.998 | <0.001 |
| IT 50% (min) | 1.80 ± 0.17b | 1.74 ± 0.13b | 6.31 ± 0.31a | 142.101 | <0.001 |
| IT 90% (min) | 10.58 ± 0.82b | 8.75 ± 0.66c | 14.45 ± 0.88a | 16.250 | 0.001 |
| Zea mays | |||||
| Coefficient a | 9.48 ± 0.18b | 10.35 ± 0.14a | 4.77 ± 0.18c | 197.121 | <0.001 |
| Coefficient b | 0.03 ± 0.07b | −0.50 ± 0.05c | 0.70 ± 0.08a | 70.279 | <0.001 |
| Coefficient c | 4.70 ± 0.61b | 10.05 ± 0.38a | 5.38 ± 0.29b | 42.101 | <0.001 |
| Amax (µmol·m−2·s−1) | 23.89 ± 0.61c | 31.03 ± 0.80b | 34.75 ± 0.75a | 63.982 | <0.001 |
| IT 50% (min) | 2.08 ± 0.83b | 1.84 ± 0.44b | 5.95 ± 0.46a | 188.271 | 0.001 |
| IT 90% (min) | 11.11 ± 0.18b | 8.58 ± 0.17c | 13.98 ± 0.16a | 20.078 | <0.001 |
| Trait | Homogeneous Shade | Heterogeneous Shade | Full Sun | F2,9 | p |
|---|---|---|---|---|---|
| Carbon (mg·g−1) | 475.12 ± 9.71 | 484.73 ± 9.60 | 483.33 ± 10.98 | 0.305 | 0.745 |
| Nitrogen (mg·g−1) | 37.11 ± 1.61a | 28.320.79b | 22.650.67c | 49.660 | <0.001 |
| Phosphorus (mg·g−1) | 2.89 ± 0.09a | 2.62 ± 0.08b | 2.82 ± 0.06a | 5.531 | 0.027 |
| Potassium (mg·g−1) | 16.38 ± 0.53 | 14.90 ± 0.67 | 14.85 ± 0.49 | 2.462 | 0.140 |
| Carbon/Nitrogen | 12.91 ± 0.61c | 17.14 ± 0.48b | 21.37 ± 0.50a | 108.88 | <0.001 |
| Carbon/Phosphorus | 164.92 ± 3.60 | 185.01 ± 5.55 | 171.64 ± 5.18 | 3.823 | 0.063 |
| Carbon/Potassium | 29.06 ± 0.63 | 32.65 ± 0.95 | 32.68 ± 0.96 | 3.468 | 0.077 |
| Calcium (mg·g−1) | 3.12 ± 0.07a | 2.80 ± 0.06ab | 2.54 ± 0.05b | 22.464 | 0.001 |
| Magnesium (mg·g−1) | 2.30 ± 0.05 | 2.21 ± 0.04 | 2.38 ± 0.06 | 2.565 | 0.131 |
| Sulfur (mg·g−1) | 1.64 ± 0.04a | 1.21 ± 0.04b | 1.26 ± 0.03b | 40.880 | <0.001 |
| Trait | Homogeneous Shade | Heterogeneous Shade | Full Sun | F2,9 | p |
|---|---|---|---|---|---|
| Carbon (mg·g−1) | 420.75 ± 5.14 | 427.75 ± 3.49 | 430.50 ± 4.44 | 0.2.180 | 0.169 |
| Nitrogen (mg·g−1) | 39.63 ± 1.67a | 34.63 ± 1.58b | 29.23 ± 1.59c | 24.286 | <0.001 |
| Phosphorus (mg·g−1) | 8.69 ± 0.16a | 6.59 ± 0.13b | 5.57 ± 0.15c | 145.55 | <0.001 |
| Potassium (mg·g−1) | 36.90 ± 0.79a | 28.53 ± 0.46b | 21.19 ± 0.69c | 49.93 | <0.001 |
| Carbon/Nitrogen | 10.63 ± 0.28c | 12.37 ± 0.25b | 14.88 ± 0.92a | 14.91 | 0.001 |
| Carbon/Phosphorus | 48.40 ± 0.38c | 65.03 ± 1.32b | 77.52 ± 2.03a | 106.58 | <0.001 |
| Carbon/Potassium | 11.41 ± 0.22c | 15.01 ± 0.23b | 20.67 ± 0.49a | 28.311 | <0.001 |
| Calcium (mg·g−1) | 5.39 ± 0.06a | 5.13 ± 0.05b | 3.72 ± 0.09c | 125.41 | <0.001 |
| Magnesium (mg·g−1) | 3.02 ± 0.09c | 3.39 ± 0.08b | 3.89 ± 0.10a | 21.590 | <0.001 |
| Sulfur (mg·g−1) | 3.82 ± 0.06a | 3.29 ± 0.10b | 2.12 ± 0.07c | 116.58 | <0.001 |
| Trait | Homogeneous Shade | Heterogeneous Shade | Full Sun | F2,9 | p |
|---|---|---|---|---|---|
| Carbon (mg·g−1) | 400.42 ± 11.63 | 404.08 ± 10.91 | 400.70 ± 9.27 | 0.043 | 0.958 |
| Nitrogen (mg·g−1) | 63.87 ± 1.17a | 53.91 ± 1.79b | 53.13 ± 1.61b | 17.310 | 0.001 |
| Phosphorus (mg·g−1) | 11.23 ± 0.48a | 9.41 ± 0.24b | 10.12 ± 0.64a | 5.874 | 0.023 |
| Potassium (mg·g−1) | 43.55 ± 1.61a | 38.13 ± 1.40b | 36.68 ± 1.28b | 6.345 | 0.019 |
| Carbon/Nitrogen | 6.28 ± 0.27b | 7.49 ± 0.44a | 7.56 ± 0.39a | 5.107 | 0.033 |
| Carbon/Phosphorus | 35.97 ± 2.39 | 44.94 ± 2.23 | 39.70 ± 2.71 | 3.316 | 0.083 |
| Carbon/Potassium | 9.92 ± 0.55 | 10.61 ± 0.64 | 10.97 ± 0.65 | 3.389 | 0.080 |
| Calcium (mg·g−1) | 17.25 ± 1.42 | 14.88 ± 0.90 | 14.13 ± 0.94 | 2.468 | 0.140 |
| Magnesium (mg·g−1) | 8.55 ± 1.78 | 8.43 ± 2.11 | 9.13 ± 2.18 | 0.034 | 0.967 |
| Sulfur (mg·g−1) | 6.23 ± 0.27a | 4.95 ± 0.43b | 5.95 ± 0.35a | 4.842 | 0.037 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marler, T.E. Artifleck: The Study of Artifactual Responses to Light Flecks with Inappropriate Leaves. Plants 2020, 9, 905. https://doi.org/10.3390/plants9070905
Marler TE. Artifleck: The Study of Artifactual Responses to Light Flecks with Inappropriate Leaves. Plants. 2020; 9(7):905. https://doi.org/10.3390/plants9070905
Chicago/Turabian StyleMarler, Thomas E. 2020. "Artifleck: The Study of Artifactual Responses to Light Flecks with Inappropriate Leaves" Plants 9, no. 7: 905. https://doi.org/10.3390/plants9070905
APA StyleMarler, T. E. (2020). Artifleck: The Study of Artifactual Responses to Light Flecks with Inappropriate Leaves. Plants, 9(7), 905. https://doi.org/10.3390/plants9070905