Stand-Alone and Combinatorial Effects of Plant-based Biostimulants on the Production and Leaf Quality of Perennial Wall Rocket

 ,
,  ,
, 
 ,
,  ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Results
2.1. Yield and Dry Biomass Production
2.2. Soil Plant Analysis Development (SPAD) index and International Commission on Illumination (CIELAB) Color Parameters
2.3. Leaf Mineral Composition
2.4. Qualitative Parameters
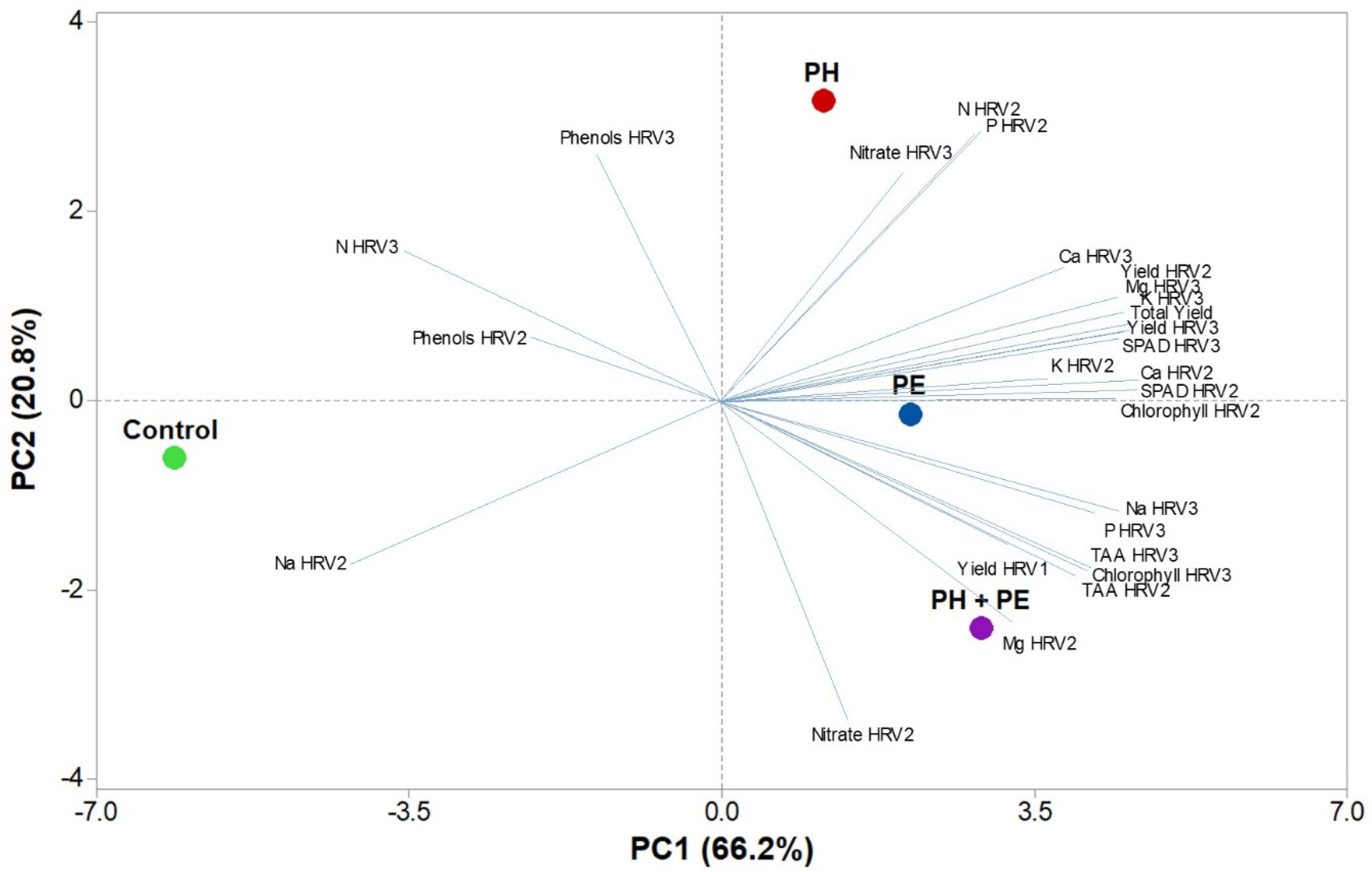
2.5. Principal Component Analysis (PCA)
2.6. Partial Budget Analysis (PBA) of Biostimulant-Treated Wall Rocket Production
3. Discussion
4. Materials and Methods
4.1. Experimental Site, Plant Material and Greenhouse Conditions
4.2. Experimental Protocol and Biostimulant Treatments
4.3. Harvest and Yield Determination
4.4. SPAD and Leaf Color Parameters
4.5. Total Nitrogen and Minerals Content
4.6. Total Chlorophyll Content
4.7. Total Ascorbic Acid and Total Phenols
4.8. Partial Budget Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Colla, G.; Rouphael, Y. Biostimulants in horticulture. Sci. Hortic. 2015, 196, 1–2. [Google Scholar] [CrossRef]
- Zhang, X.; Schmidt, R.E. The impact of growth regulators on alphatocopherol status of water-stressed Poa pratensis L. Int. Turfgrass Soc. Res. J. 1997, 8, 1364–1373. [Google Scholar]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. HortScience 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [Green Version]
- Arora, N.K.; Mishra, J. Prospecting the roles of metabolites and additives in future bioformulations for sustainable agriculture. Appl. Soil Ecol. 2016, 107, 405–407. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global prospective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.; Bonini, P.; Colla, G. Plant and seaweed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Di Filippo-Herrera, D.A.; Muñoz-Ochoa1, M.; Hernández-Herrera, R.M.; Hernández-Carmona, G. Biostimulant activity of individual and blended seaweed extracts on the germination and growth of the mung bean. J. Appl. Phycol. 2019, 31, 2025–2037. [Google Scholar] [CrossRef]
- Caruso, G.; Parrella, G.; Giorgini, M.; Nicoletti, R. Crop systems, quality and protection of Diplotaxis tenuifolia. Agriculture 2018, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Giordano, M.; El-Nakhel, C.; Cuciniello, A.; Cenvinzo, V.; Colla, G.; Rouphael, Y. Protein hydrolysate or plant extract-based biostimulants enhanced yield and quality performances of greenhouse perennial wall rocket grown in different seasons. Plants 2019, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Conforti, F.; Perri, V.; Menichini, F.; Marrelli, M.; Uzunov, D.; Statti, G.A.; Menichini, F. Wild Mediterranean dietary plants as inhibitors of pancreatic lipase. Phytother. Res. 2012, 26, 600–604. [Google Scholar] [CrossRef] [PubMed]
- O’Hare, T.J.; Wong, L.S.; Irving, D.E. Asian and western horticultural species of the Brassica family with anti-cancer potential. Acta Hortic. 2005, 694, 457–462. [Google Scholar] [CrossRef]
- Martínez-Sánchez, A.; Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F. A comparative study of flavonoid compounds, vitamin C, and antioxidant properties of baby leaf Brassicaceae species. J. Agric. Food Chem. 2008, 56, 2330–2340. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic action of a microbial-based biostimulant and a plant derived-protein hydrolysate enhances lettuce tolerance to alkalinity and salinity. Front. Plant Sci. 2017, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; et al. Morphological and physiological responses induced by protein hydrolysate-based biostimulant and nitrogen rates in greenhouse spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Pimpini, F.; Giannini, M.; Lazzarin, R. Ortaggi da Foglia e da Taglio; Veneto Agricoltura: Venezia, Italy, 2005; p. 118. [Google Scholar]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Giordano, M.; El-Nakhel, C.; Sacco, A.; Rouphael, Y.; Colla, G.; Mori, M. Plant-based biostimulants influence the agronomical, physiological, and qualitative responses of baby rocket leaves under diverse nitrogen conditions. Plants 2019, 8, 522. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, M.; Ertani, A.; Nardi, S. Effects of an alfalfa protein hydrolysate on the gene expression and activity of enzymes of the tricarboxylic acid (TCA) cycle and nitrogen metabolism in Zea mays L. J. Agric. Food Chem. 2008, 56, 11800–11808. [Google Scholar] [CrossRef]
- Foroutan Nia, A.; Naghdi Badi, H.; Mehrafarin, A.; Bahman, S.; Seif Sahandi, M. Changes in the essential oil content and terpene composition of rosemary (Rosmarinus officinalis L.) by using plant biostimulants. Acta Agric. 2016, 107, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Cerdán, M.; Sánchez-Sánchez, A.; Jordá, J.D.; Juárez, M.; Sánchez-Andreu, J. Effect of commercial amino acids on iron nutrition of tomato plants grown under lime-induced iron deficiency. J. Plant Nutr. Soil Sci. 2013, 176, 859–866. [Google Scholar] [CrossRef]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activities of two protein hydrolysates on the growth and nitrogen metabolism in maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Canali, S.; Diacono, M.; Ciaccia, C.; Masetti, O.; Tittarelli, F.; Montemurro, F. Alternative strategies for nitrogen fertilization of overwinter processing spinach (Spinacia oleracea L.) in Southern Italy. Eur. J. Agron. 2014, 54, 47–53. [Google Scholar] [CrossRef]
- Citak, S.; Sonmez, S. Effects of conventional and organic fertilization on spinach (Spinacea oleracea L.) growth, yield, vitamin C and nitrate concentration during two successive seasons. Sci. Hortic. 2010, 126, 415–420. [Google Scholar] [CrossRef]
- Pinto, E.; Agostinho, A.; Aguiar, A.R.M.; Ferreira Isabel, M.P.L.V.O. Changes in macrominerals, trace elements and pigments content during lettuce (Lactuca sativa L.) growth: Nfluence of soil composition. Food Chem. 2014, 152, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Burns, I.G.; Zhang, K.; Turner, M.K.; Meacham, M.; Al-Redhiman Khalid, L.J.; Broadley, M.R.; Hand, P.; Pink, D. Screening for genotype and environment effects on nitrate accumulation in 24 species of young lettuce. J. Sci. Food Agric. 2011, 91, 553–562. [Google Scholar] [CrossRef]
- Burns, I.G.; Durnford, J.; Lynn, J.; McClement, S.; Hand, P.; Pink, D. The influence of genetic variation and nitrogen source on nitrate accumulation and iso-osmotic regulation by lettuce. Plant Soil 2012, 352, 321–339. [Google Scholar] [CrossRef]
- Chen, B.M.; Wang, Z.H.; Li, S.X.; Wang, G.X.; Song, H.X.; Wang, X.N. Effects of nitrate supply on plant growth, nitrate accumulation, metabolic nitrate concentration and nitrate reductase activity in three leafy vegetables. Plant Sci. 2004, 167, 635–643. [Google Scholar] [CrossRef]
- Chung, J.B.; Yong-Woo, L.; Hee-Youl, C.; Yong, P.; Moon-Soo, C. Genotypical variation in nitrate accumulation of lettuce and spinach. Korean J. Soil Sci. Fert. 2005, 38, 38–44. [Google Scholar]
- Correia, M.; Ângela Barroso, M.; Barroso, F.; Soares, D.; Oliveira, M.B.P.P.; Delerue-Matos, C. Contribution of different vegetable types to exogenous nitrate and nitrite exposure. Food Chem. 2010, 120, 960–966. [Google Scholar] [CrossRef] [Green Version]
- Prasad, S.; Chetty, A.A. Nitrate-N determination in leafy vegetables: Study of the effects of cooking and freezing. Food Chem. 2008, 106, 772–780. [Google Scholar] [CrossRef]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Santamaria, P.; Gonnella, M.; Elia, A.; Parente, A.; Serio, F. Ways of reducing rocket salad nitrate content. Acta Hortic. 2001, 548, 529–536. [Google Scholar] [CrossRef]
- Santamaria, P.; Elia, A.; Serio, F. Effect of solution nitrogen concentration on yield, leaf element content, and water and nitrogen use efficiency of three hydroponically-grown rocket salad genotypes. J. Plant Nutr. 2002, 25, 245–258. [Google Scholar] [CrossRef]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health beneficts of lettuce (Lactuca sativa L.). J. Food Comp. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Buchanan, B.B. The carbon (formerly dark) reactions of photosynthesis. Photosynth. Res. 2016, 128, 215–217. [Google Scholar] [CrossRef]
- Guo, W.; Nazim, H.; Liang, Z.; Yang, D. Magnesium deficiency in plants: An urgent problem. Crop. J. 2016, 4, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, P.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant application with a tropical plant extract enhances Corchorus olitorius adaptation to sub-optimal nutrient regimens by improving physiological parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [Green Version]
- Smirnoff, N. Ascorbic acid metabolism and functions: A comparison of plants and mammals. Free Radic. Biol. Med. 2018, 122, 116–129. [Google Scholar] [CrossRef]
- Bonasia, A.; Lazzizera, C.; Elia, A.; Conversa, G. Nutritional, biophysical and physiological characteristics of wild rocket genotypes as affected by soilless cultivation system, salinity level of nutrient solution and growing period. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Salvatore, S.; Pellegrini, N.; Brenna, O.V.; Del Rio, D.; Frasca, G.; Brighenti, F.; Tumino, R. Antioxidant characterization of some Sicilian edible wild greens. J. Agric. Food Chem. 2005, 53, 9465–9471. [Google Scholar] [CrossRef]
- Durazzo, A.; Azzini, E.; Lazzè, M.C.; Raguzzini, A.; Pizzala, R.; Maiani, G. Italian wild rocket [Diplotaxis tenuifolia (L.) D.C.]: Influence of agricultural practices on antioxidant molecules and on cytotoxicity and antiproliferative effects. Agriculture 2013, 3, 285–298. [Google Scholar] [CrossRef] [Green Version]
- USDA. National Nutrient Database for Standard Reference Release 28; USDA: Washington, DC, USA, 2015. [Google Scholar]
- Machado, R.M.A.; Alves-Pereira, I.; Ferreira, R.M.A. Plant growth, phytochemical accumulation and antioxidant activity of substrate-grown spinach. Heliyon 2018, 4, e00751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purushothaman, B.; Suganthi, N.; Shanmugam, K. Qualitative and quantitative determination of various extracts of Ocimum basilicum L. leaves. J. Nat. Remedies 2019. [Google Scholar] [CrossRef]
- Ertani, A.; Pizzeghello, O.; Francioso, P.; Sambo, S.; Sanchez-Cortes, S.; Nardi, S. Capsicum chinensis L. growth and nutraceutical properties are enhanced by biostimulants in a long-term period: Chemical metabolomic approaches. Front. Plant Sci. 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landi, M.; Fambrini, M.; Basile, A.; Salvini, M.; Guidi, L.; Pugliesi, C. Overexpression of L-galactono-1,4-lactone dehydrogenase (L-GalLDH) gene correlates with increased ascorbate concentration and reduced browning in leaves of Lactuca sativa L. after cutting. Plant Cell Tissue Organ. Cult. 2015, 123, 109–120. [Google Scholar] [CrossRef]
- Bremner, J.M. Total nitrogen. In Methods of Soil Analysis; Black, C.A., Evans, D.D., White, D.D., Ensminger, E., Clark, F.E., Eds.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 1149–1178. [Google Scholar]
- Lichtentahler, H.K.; Buschmann, C. Chlorophylls and carotenoids; Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Van Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Oxid. Antioxid. Part A 1999, 299, 152–178. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Biostimulant Treatments | Yield (g m−2) | Total Yield (g m−2) | Dry Biomass (g m−2) | Total Dry Biomass (g m−2) | ||||
|---|---|---|---|---|---|---|---|---|
| Harvest I | Harvest II | Harvest III | Harvest I | Harvest II | Harvest III | |||
| Control | 462.8 ± 30.7 | 832.7 ± 15 b | 403.7 ± 36 b | 1699.1 ± 20 b | 55.6 ± 1.5 | 67.3 ± 1.5 b | 30.0 ± 2.9 b | 153.0 ± 4.5 b |
| PH | 493.0 ± 47.6 | 1246.6 ± 109 a | 821.4 ± 111 a | 2561.1 ± 174 a | 53.6 ± 4.9 | 102.4 ± 13.5 a | 61.1 ± 8.5 a | 217.0 ± 16.5 a |
| PE | 476.8 ± 59.8 | 1252.2 ± 27 a | 726.8 ± 73 a | 2455.7 ± 87 a | 49.0 ± 4.8 | 99.1 ± 3.5 a | 50.9 ± 3.0 a | 198.9 ± 3.1 a |
| PH + PE | 542.7 ± 63.9 | 1176.4 ± 111 a | 816.8 ± 53 a | 2535.9 ± 19 a | 57.3 ± 5.8 | 95.1 ± 14.2 a | 61.2 ± 5.5 a | 213.5 ± 10.3 a |
| Significance | ns | * | * | *** | ns | * | * | ** |
| Biostimulant Treatments | Harvest II | Harvest III | ||||||
|---|---|---|---|---|---|---|---|---|
| SPAD | L* | a* | b* | SPAD | L* | a* | b* | |
| Control | 34.3 ± 1.20 b | 39.7 ± 0.26 | 13.8 ± 0.34 | 19.8 ± 0.48 | 32.0 ± 1.53 b | 40.4 ± 0.15 | 13.6 ± 0.39 | 19.6 ± 0.64 |
| PH | 38.7 ± 0.67 a | 38.3 ± 0.57 | 13.3 ± 0.26 | 19.4 ± 0.43 | 37.7 ± 1.45 a | 39.3 ± 0.77 | 14.1 ± 0.28 | 20.9 ± 0.64 |
| PE | 37.7 ± 0.88 a | 39.0 ± 0.85 | 13.7 ± 0.52 | 20.0 ± 0.73 | 39.7 ± 1.45 a | 39.8 ± 0.49 | 14.5 ± 0.14 | 21.3 ± 0.19 |
| PH + PE | 38.7 ± 1.76 a | 39.1 ± 0.59 | 13.6 ± 0.34 | 19.5 ± 0.79 | 39.7 ± 1.33 a | 39.5 ± 0.19 | 14.0 ± 0.31 | 20.3 ± 0.63 |
| Significance | * | ns | ns | ns | * | ns | ns | ns |
| Biostimulant Treatments | N (g kg−1 dw) | P (g kg−1 dw) | K (g kg−1 dw) | Ca (g kg−1 dw) | Mg (g kg−1 dw) | Na (g kg−1 dw) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Harvest II | Harvest III | Harvest II | Harvest III | Harvest II | Harvest III | Harvest II | Harvest III | Harvest II | Harvest III | Harvest II | Harvest III | |
| Control | 4.85 ± 0.10 | 5.03 ± 0.10 | 2.83 ± 0.19 | 2.93 ± 0.16 | 44.5 ± 0.51 b | 37.5 ± 1.43 b | 18.1 ± 0.10 b | 26.2 ± 0.20 b | 3.03 ± 0.24 | 2.90 ± 0.21 b | 2.96 ± 0.10 | 2.64 ± 0.78 |
| PH | 5.01 ± 0.14 | 4.99 ± 0.18 | 3.20 ± 0.05 | 3.02 ± 0.14 | 48.1 ± 0.64 b | 54.3 ± 1.34 a | 23.8 ± 0.38 a | 34.0 ± 0.54 a | 3.13 ± 0.12 | 4.16 ± 0.35 a | 2.16 ± 0.43 | 4.03 ± 0.14 |
| PE | 4.95 ± 0.18 | 4.71 ± 0.16 | 3.14 ± 0.13 | 3.08 ± 0.06 | 52.9 ± 2.26 a | 53.9 ± 2.19 a | 25.0 ± 0.56 a | 30.0 ± 2.10 ab | 3.31 ± 0.34 | 3.87 ± 0.24 a | 2.57 ± 0.66 | 3.47 ± 0.67 |
| PH + PE | 4.89 ± 0.16 | 4.80 ± 0.15 | 2.87 ± 0.10 | 3.10 ± 0.17 | 48.3 ± 1.28 b | 56.6 ± 1.61 a | 25.1 ± 0.34 a | 32.4 ± 1.47 a | 3.32 ± 0.32 | 4.02 ± 0.10 a | 2.02 ± 0.87 | 4.42 ± 0.22 |
| Significance | ns | ns | ns | ns | * | *** | *** | * | ns | * | ns | ns |
| Biostimulant Treatments | Chlorophyll | Nitrate | Total Phenols | Total Ascorbic Acid | ||||
|---|---|---|---|---|---|---|---|---|
| (mg g−1 fw) | (mg kg−1 fw) | (mg gallic acid eq. g−1 dw) | (mg g−1 fw) | |||||
| Harvest II | Harvest III | Harvest II | Harvest III | Harvest II | Harvest III | Harvest II | Harvest III | |
| Control | 0.96 ± 0.14 | 0.88 ± 0.09 b | 4630 ± 644 | 4428 ± 446 | 3.98 ± 0.07 | 4.71 ± 0.11 | 17.3 ± 3.22 c | 14.9 ± 2.11 b |
| PH | 1.34 ± 0.17 | 1.03 ± 0.06 ab | 4558 ± 375 | 5611 ± 377 | 3.95 ± 0.38 | 5.03 ± 0.67 | 23.6 ± 1.11 bc | 22.0 ± 2.08 b |
| PE | 1.22 ± 0.14 | 1.11 ± 0.05 a | 4634 ± 211 | 5897 ± 489 | 3.46 ± 0.20 | 4.32 ± 0.14 | 28.0 ± 0.53 b | 31.3 ± 2.29 a |
| PH + PE | 1.43 ± 0.18 | 1.22 ± 0.04 a | 4772 ± 396 | 4380 ± 631 | 3.84 ± 0.06 | 4.46 ± 0.10 | 37.0 ± 1.79 a | 36.3 ± 3.25 a |
| Significance | ns | * | ns | ns | ns | ns | *** | *** |
| Biostimulant | Added Gross Returns (€ ha−1) | Added Variable Costs (€ ha−1) | Added Net Returns (€ ha−1) | |||
|---|---|---|---|---|---|---|
| Biostimulant treatment | Foliar spraying | Machine harvest | Total | |||
| Protein Hydrolysates (PH) | 11354.4 | 676.0 | 390 | 343.3 | 1409.3 | 9945.1 |
| Tropical Plant Extract (PE) | 9905.8 | 806.0 | 390 | 301.7 | 1497.7 | 8408.1 |
| PH + PE | 11276.6 | 741.0 | 390 | 333.4 | 1464.4 | 9812.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giordano, M.; El-Nakhel, C.; Caruso, G.; Cozzolino, E.; De Pascale, S.; Kyriacou, M.C.; Colla, G.; Rouphael, Y. Stand-Alone and Combinatorial Effects of Plant-based Biostimulants on the Production and Leaf Quality of Perennial Wall Rocket. Plants 2020, 9, 922. https://doi.org/10.3390/plants9070922
Giordano M, El-Nakhel C, Caruso G, Cozzolino E, De Pascale S, Kyriacou MC, Colla G, Rouphael Y. Stand-Alone and Combinatorial Effects of Plant-based Biostimulants on the Production and Leaf Quality of Perennial Wall Rocket. Plants. 2020; 9(7):922. https://doi.org/10.3390/plants9070922
Chicago/Turabian StyleGiordano, Maria, Christophe El-Nakhel, Gianluca Caruso, Eugenio Cozzolino, Stefania De Pascale, Marios C. Kyriacou, Giuseppe Colla, and Youssef Rouphael. 2020. "Stand-Alone and Combinatorial Effects of Plant-based Biostimulants on the Production and Leaf Quality of Perennial Wall Rocket" Plants 9, no. 7: 922. https://doi.org/10.3390/plants9070922
APA StyleGiordano, M., El-Nakhel, C., Caruso, G., Cozzolino, E., De Pascale, S., Kyriacou, M. C., Colla, G., & Rouphael, Y. (2020). Stand-Alone and Combinatorial Effects of Plant-based Biostimulants on the Production and Leaf Quality of Perennial Wall Rocket. Plants, 9(7), 922. https://doi.org/10.3390/plants9070922