Genome-Wide Analysis Reveals Transcription Factors Regulated by Spider-Mite Feeding in Cucumber (Cucumis sativus)


Abstract
:1. Introduction
2. Results
2.1. TF Genes in the Cucumber Genome
2.2. Cucumber Genes Responsive to Spider-Mite Feeding
2.3. Regulation of TF Expression in Response to Spider-Mite Herbivory
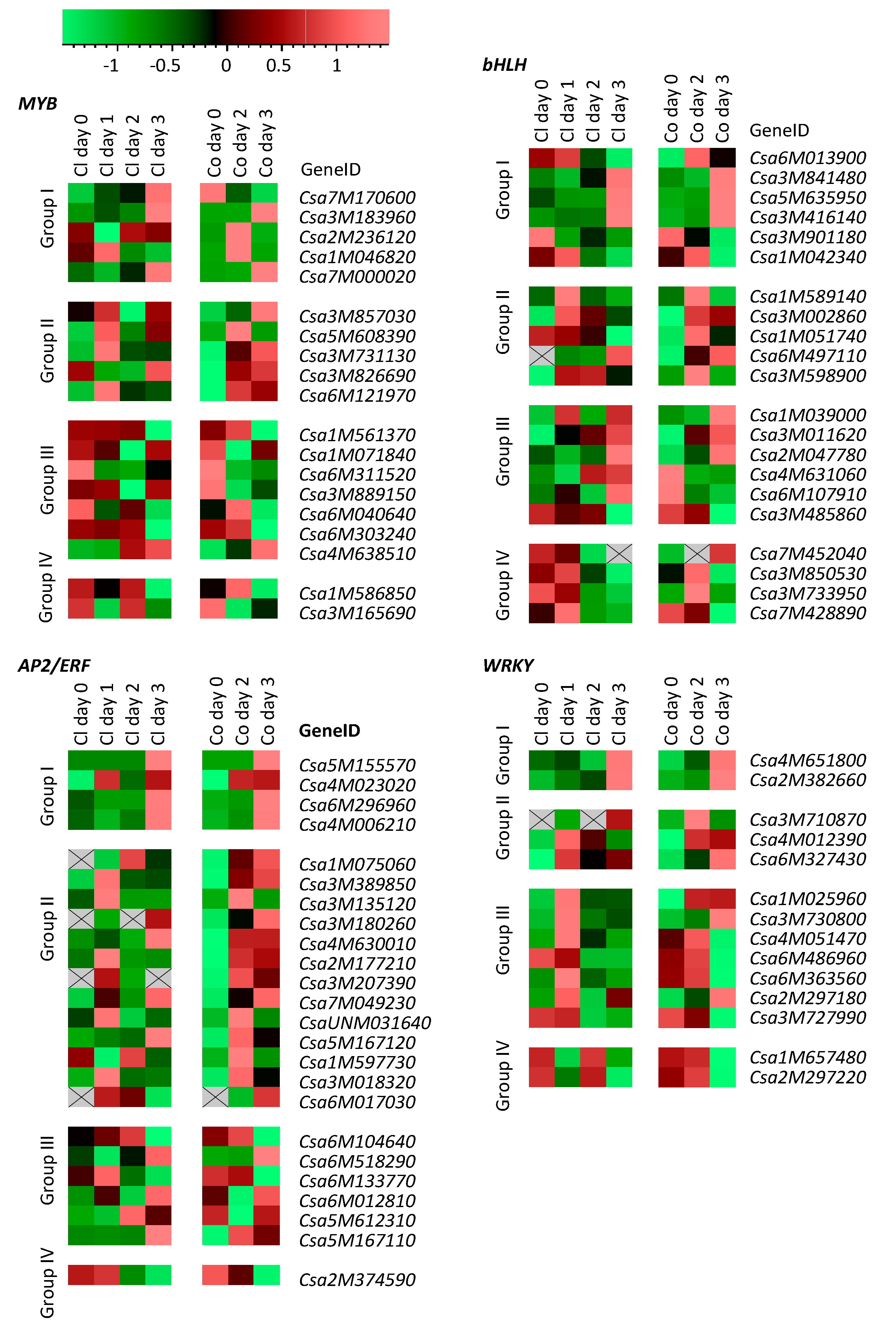
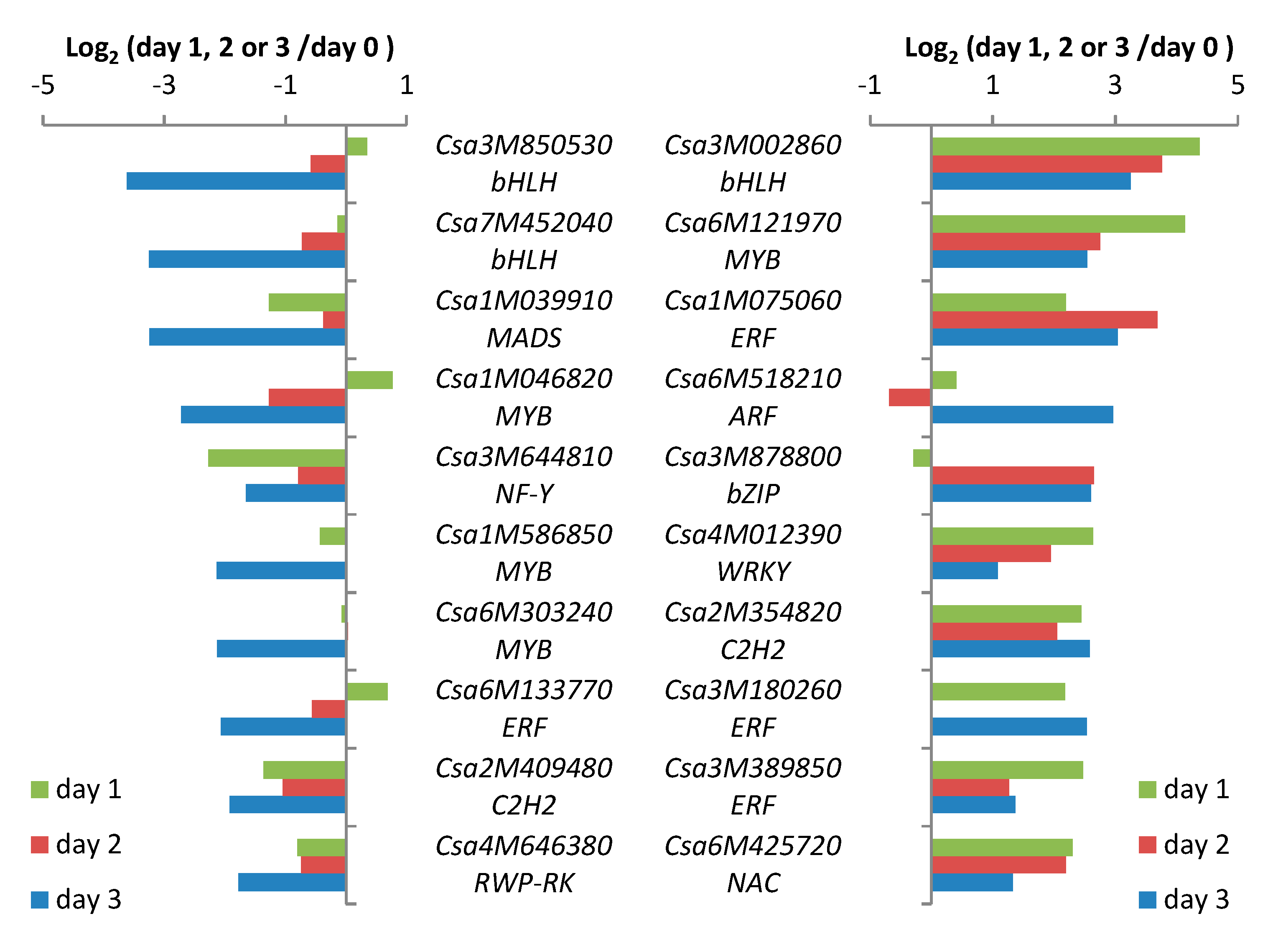
2.4. TF Genes Most Strongly Influenced by Spider-Mite Feeding
2.5. Co-Expression of TF Genes with Defense-Related Genes
2.6. Motifs in Promoters of Defense-Related Genes
3. Discussion
3.1. TF Genes in the Cucumber Genome
3.2. TF Genes Responsive to Spider-Mite Feeding
3.3. Possible Regulatory Relationships between TFs and Metabolite Biosynthesis Associated Genes
4. Materials and Methods
4.1. Plants and Mites
4.2. Identification of TF Genes in the Cucumber Genome
4.3. Grouping of the DEGs and Identification of TFs Responses to Feeding by Spider Mites or Downy Mildew
4.4. Co-Expression Analysis of TF Genes and Plant Defense-Associated Genes
4.5. Identification of Binding Motifs in the Promoter Region
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Transcription factors | |
| AP2/ERF | apetala2/ethylene response factor |
| ARF | auxin response factors |
| bHLH | basic/helix-loop-helix |
| bZIP | basic leucine zipper |
| C2H2 | Cys2His2 zinc finger |
| DOF | DNA binding with one finger |
| GRAS | GAI, gibberellin-acid insensitive |
| RGA, repressor of GA1 | |
| SCR, scarecrow | |
| HOX | homeodomain-containing |
| HSF | heat stress transcription factor |
| MADS | MCM1, mini chromosome maintenance 1 of yeast (Saccharomyces cerevisiae) |
| AG, agamous of Arabidopsis (Arabidopsis thaliana) | |
| DEF, deficiens of snapdragon (Antirrhinum majus) | |
| SRF, serum response factor of humans (Homo sapiens) | |
| MYB | myeloblastosis |
| NAC | NAM, no apical meristem |
| ATAF1-2 | |
| CUC2, cup-shaped cotyledon | |
| NF-Y | nuclear factor Y |
| SBP | SQUAMOSA promoter binding proteins |
| TCP | TB1, teosinte branched1 from maize (Zea mays) |
| CYC, CYCLOIDEA from snapdragon (Antirrhinum majus) | |
| PCF, proliferating cell factors from rice (Oryza sativa) | |
| WRKY | WRKYGQK heptapeptide |
| Others | |
| CARE | cis-acting regulatory element |
| Cl | Chinese long |
| Co | corona |
| DEG | differentially expressed gene |
| DXS | 1-deoxy-d-xylulose-5-phosphate synthase |
| LOX | lipoxygenase |
| RPKM | reads per kilobase of transcript per million mapped reads |
| SUT | sucrose transporter |
| TF | transcription factor |
| TPS | terpene synthases |
| UGT | UDP-glycosyltransferase |
References
- Latchman, D.S. Transcription factors: An overview. Int. J. Biochem. Cell B 1997, 29, 1305–1312. [Google Scholar] [CrossRef] [Green Version]
- Wittkopp, P.J.; Kalay, G. Cis-regulatory elements: Molecular mechanisms and evolutionary processes underlying divergence. Nat. Rev. Genet. 2012, 13, 59–69. [Google Scholar] [CrossRef]
- Jin, J.; Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, G. Regulation of the initiation of eukaryotic transcription. Essays Biochem. 2001, 37, 33–43. [Google Scholar] [PubMed]
- Narlikar, G.J.; Fan, H.Y.; Kingston, R.E. Cooperation between complexes that regulate chromatin structure and transcription. Cell 2002, 108, 475–487. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Glass, C.K.; Rosenfeld, M.G. Coactivator and corepressor complexes in nuclear receptor function. Curr. Opin. Genet. Dev. 1999, 9, 140–147. [Google Scholar] [CrossRef]
- Guo, A.Y.; He, K.; Liu, D.; Bai, S.N.; Gu, X.C.; Wei, L.P.; Luo, J.C. DATF: A database of Arabidopsis transcription factors. Bioinformatics 2005, 21, 2568–2569. [Google Scholar] [CrossRef]
- Lehti-Shiu, M.D.; Shiu, S.H. Diversity, classification and function of the plant protein kinase superfamily. Philos. Trans. R. Soc. B 2012, 367, 2619–2639. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; He, K.; Tang, X.; Li, Z.; Lv, L.; Zhao, Y.; Luo, J.; Gao, G. An Arabidopsis transcriptional regulatory map reveals distinct functional and evolutionary features of novel transcription factors. Mol. Biol. Evol. 2015, 32, 1767–1773. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K.; et al. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.W.; Li, R.Q.; Zhang, Z.H.; Li, L.; Gu, X.F.; Fan, W.; Lucas, W.J.; Wang, X.W.; Xie, B.Y.; Ni, P.X.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhang, Z.; Yan, P.; Huang, S.; Fei, Z.; Lin, K. RNA-Seq improves annotation of protein-coding genes in the cucumber genome. BMC Genom. 2011, 12, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mithofer, A.; Boland, W. Recognition of herbivory-associated molecular patterns. Plant Physiol. 2008, 146, 825–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazebnik, J.; Frago, E.; Dicke, M.; van Loon, J.J.A. Phytohormone mediation of interactions between herbivores and plant pathogens. J. Chem. Ecol. 2014, 40, 730–741. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. In Annual Review of Cell and Developmental Biology; Schekman, R., Ed.; Annual Reviews: Palo Alto, CA, USA, 2012; Volume 28, pp. 489–521. [Google Scholar]
- Wu, J.Q.; Baldwin, I.T. New insights into plant responses to the attack from insect herbivores. Annu. Rev. Genet. 2010, 44, 1–24. [Google Scholar] [CrossRef]
- Berr, A.; Menard, R.; Heitz, T.; Shen, W.H. Chromatin modification and remodelling: A regulatory landscape for the control of Arabidopsis defence responses upon pathogen attack. Cell Microbiol. 2012, 14, 829–839. [Google Scholar] [CrossRef]
- Li, D.; Halitschke, R.; Baldwin, I.T.; Gaquerel, E. Information theory tests critical predictions of plant defense theory for specialized metabolism. Sci. Adv. 2020, 6, eaaz0381. [Google Scholar] [CrossRef]
- Perez-Rodriguez, P.; Riano-Pachon, D.M.; Correa, L.G.; Rensing, S.A.; Kersten, B.; Mueller-Roeber, B. PlnTFDB: Updated content and new features of the plant transcription factor database. Nucleic Acids Res. 2010, 38, D822–D827. [Google Scholar] [CrossRef] [Green Version]
- Seo, E.; Choi, D. Functional studies of transcription factors involved in plant defenses in the genomics era. Brief. Funct. Genom. 2015, 14, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Gigolashvili, T.; Berger, B.; Mock, H.P.; Muller, C.; Weisshaar, B.; Fluegge, U.I. The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana. Plant J. 2007, 50, 886–901. [Google Scholar] [CrossRef] [Green Version]
- Onkokesung, N.; Reichelt, M.; van Doorn, A.; Schuurink, R.C.; van Loon, J.J.A.; Dicke, M. Modulation of flavonoid metabolites in Arabidopsis thaliana through overexpression of the MYB75 transcription factor: Role of kaempferol-3,7-dirhamnoside in resistance to the specialist insect herbivore Pieris brassicae. J. Exp. Bot. 2014, 65, 2203–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chezem, W.R.; Memon, A.; Li, F.-S.; Weng, J.-K.; Clay, N.K. SG2-Type R2R3-MYB Transcription Factor MYB15 Controls Defense-Induced Lignification and Basal Immunity in Arabidopsis. Plant Cell 2017, 29, 1907–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, H.; YamaguchiShinozaki, K.; Urao, T.; Iwasaki, T.; Hosokawa, D.; Shinozaki, K. Role of Arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression. Plant Cell 1997, 9, 1859–1868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, H.; Heinzel, N.; Schottner, M.; Baldwin, I.T.; Galis, I. R2R3-NaMYB8 regulates the accumulation of phenylpropanoid-polyamine conjugates, which are essential for local and systemic defense against insect herbivores in nicotiana attenuata. Plant Physiol. 2010, 152, 1731–1747. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, O.; Chico, J.M.; Sanchez-Serrano, J.J.; Solano, R. Jasmonate-insensitive1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 2004, 16, 1938–1950. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Calvo, P.; Chini, A.; Fernandez-Barbero, G.; Chico, J.M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, D.; Schweizer, F.; Godoy, M.; Franco-Zorrilla, J.M.; et al. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell 2011, 23, 701–715. [Google Scholar] [CrossRef] [Green Version]
- Goossens, J.; Mertens, J.; Goossens, A. Role and functioning of bHLH transcription factors in jasmonate signalling. J. Exp. Bot. 2016, 68, 1333–1347. [Google Scholar] [CrossRef]
- Li, S. Transcriptional control of flavonoid biosynthesis: Fine-tuning of the MYB-bHLH-WD40 (MBW) complex. Plant Signal. Behav. 2014, 9, e27522. [Google Scholar] [CrossRef]
- Dombrecht, B.; Xue, G.P.; Sprague, S.J.; Kirkegaard, J.A.; Ross, J.J.; Reid, J.B.; Fitt, G.P.; Sewelam, N.; Schenk, P.M.; Manners, J.M.; et al. MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis. Plant Cell 2007, 19, 2225–2245. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, F.; Fernandez-Calvo, P.; Zander, M.; Diez-Diaz, M.; Fonseca, S.; Glauser, G.; Lewsey, M.G.; Ecker, J.R.; Solano, R.; Reymond, P. Arabidopsis basic helix-loop-helix transcription factors MYC2, MYC3, and MYC4 regulate glucosinolate biosynthesis, insect performance, and feeding behavior. Plant Cell 2013, 25, 3117–3132. [Google Scholar] [CrossRef] [Green Version]
- Moffat, C.S.; Ingle, R.A.; Wathugala, D.L.; Saunders, N.J.; Knight, H.; Knight, M.R. ERF5 and ERF6 play redundant roles as positive regulators of JA/Et-mediated defense against Botrytis cinerea in Arabidopsis. PLoS ONE 2012, 7, e35995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Wei, T.; Yin, K.Q.; Chen, Z.L.; Gu, H.Y.; Qu, L.J.; Qin, G.J. Arabidopsis RAP2.2 plays an important role in plant resistance to Botrytis cinerea and ethylene responses. New Phytol. 2012, 195, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Ju, H.P.; Zhou, G.X.; Zhu, C.S.; Erb, M.; Wang, X.P.; Wang, P.; Lou, Y.G. An EAR-motif-containing ERF transcription factor affects herbivore-induced signaling, defense and resistance in rice. Plant J. 2011, 68, 583–596. [Google Scholar] [CrossRef]
- Deslandes, L.; Olivier, J.; Theulieres, F.; Hirsch, J.; Feng, D.X.; Bittner-Eddy, P.; Beynon, J.; Marco, Y. Resistance to Ralstonia solanacearum in Arabidopsis thaliana is conferred by the recessive RRS1-R gene, a member of a novel family of resistance genes. Proc. Natl. Acad. Sci. USA 2002, 99, 2404–2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.Y.; Abu Qamar, S.; Chen, Z.X.; Mengiste, T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J. 2006, 48, 592–605. [Google Scholar] [CrossRef]
- Skibbe, M.; Qu, N.; Galis, I.; Baldwin, I.T. Induced plant defenses in the natural environment: Nicotiana attenuata WRKY3 and WRKY6 coordinate responses to herbivory. Plant Cell 2008, 20, 1984–2000. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Zhang, J.; Li, J.; Zhou, G.; Wang, Q.; Bian, W.; Erb, M.; Lou, Y. Prioritizing plant defence over growth through WRKY regulation facilitates infestation by non-target herbivores. eLife 2015, 4, e04805. [Google Scholar] [CrossRef] [Green Version]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari: A review. Insect Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Zhurov, V.; Navarro, M.; Bruinsma, K.A.; Arbona, V.; Santamaria, M.E.; Cazaux, M.; Wybouw, N.; Osborne, E.J.; Ens, C.; Rioja, C.; et al. Reciprocal responses in the interaction between arabidopsis and the cell-content-feeding chelicerate herbivore spider mite. Plant Physiol. 2014, 164, 384–399. [Google Scholar] [CrossRef] [Green Version]
- Martel, C.; Zhurov, V.; Navarro, M.; Martinez, M.; Cazaux, M.; Auger, P.; Migeon, A.; Santamaria, M.E.; Wybouw, N.; Diaz, I.; et al. Tomato whole genome transcriptional response to Tetranychus urticae identifies divergence of spider mite-induced responses between tomato and Arabidopsis. Mol. Plant-Microbe Interact. 2015, 28, 343–361. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Riquelme, J.; Zhurov, V.; Rioja, C.; Pérez-Moreno, I.; Torres-Pérez, R.; Grimplet, J.; Carbonell-Bejerano, P.; Bajda, S.; Van Leeuwen, T.; Martínez-Zapater, J.M.; et al. Comparative genome-wide transcriptome analysis of Vitis vinifera responses to adapted and non-adapted strains of two-spotted spider mite, Tetranyhus urticae. BMC Genom. 2016, 17, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Wang, G.-Q.; Zhou, Q.; Lu, W.; Ma, J.-Q.; Huang, J.-H. Transcriptomic and proteomic response of Manihot esculenta to Tetranychus urticae infestation at different densities. Exp. Appl. Acarol. 2019, 78, 273–293. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Bouwmeester, H.J.; Dicke, M.; Kappers, I.F. Transcriptional and metabolite analysis reveal a shift in direct and indirect defences in response to spider-mite infestation in cucumber (Cucumis sativus). Plant Mol. Biol. 2020, 103, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.N.; Savory, E.A.; Vaillancourt, B.; Childs, K.L.; Hamilton, J.P.; Day, B.; Buell, C.R. Expression profiling of Cucumis sativus in response to infection by Pseudoperonospora cubensis. PLoS ONE 2012, 7, e34954. [Google Scholar] [CrossRef] [Green Version]
- Gigot, C.; Ongena, M.; Fauconnier, M.L.; Wathelet, J.P.; Du Jardin, P.; Thonart, P. The lipoxygenase metabolic pathway in plants: Potential for industrial production of natural green leaf volatiles. Biotechnol. Agron. Soc. 2010, 14, 451–460. [Google Scholar]
- Mercke, P.; Kappers, I.F.; Verstappen, F.W.A.; Vorst, O.; Dicke, M.; Bouwmeester, H.J. Combined transcript and metabolite analysis reveals genes involved in spider mite induced volatile formation in cucumber plants. Plant Physiol. 2004, 135, 2012–2024. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- King, C.D.; Rios, G.R.; Green, M.D.; Tephly, T.R. UDP-glucuronosyltransferases. Curr. Drug Metab. 2000, 1, 143–161. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Shang, Y.; Ma, Y.S.; Zhou, Y.; Zhang, H.M.; Duan, L.X.; Chen, H.M.; Zeng, J.G.; Zhou, Q.; Wang, S.H.; Gu, W.J.; et al. Biosynthesis, regulation, and domestication of bitterness in cucumber. Science 2014, 346, 1084–1088. [Google Scholar] [CrossRef]
- Kappers, I.F.; Verstappen, F.W.; Luckerhoff, L.L.; Bouwmeester, H.J.; Dicke, M. Genetic variation in jasmonic acid- and spider mite-induced plant volatile emission of cucumber accessions and attraction of the predator Phytoseiulus persimilis. J. Chem. Ecol. 2010, 36, 500–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappers, I.F.; Hoogerbrugge, H.; Bouwmeester, H.J.; Dicke, M. Variation in herbivory-induced volatiles among cucumber (cucumis sativus l.) varieties has consequences for the attraction of carnivorous natural enemies. J. Chem. Ecol. 2011, 37, 150–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spider Mites | Downy Mildew | Spider Mites and Downy Mildew | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TF Family | Group I | Group II | Group III | Group IV | In Total | % * | Nr | % | Nr | % |
| MYB | 5 | 5 | 7 | 2 | 19 | 10% | 26 | 14% | 5 | 3% |
| bHLH | 6 | 5 | 6 | 4 | 21 | 14% | 22 | 15% | 6 | 4% |
| AP2/ERF | 4 | 13 | 6 | 1 | 24 | 16% | 29 | 20% | 14 | 10% |
| C2H2 | 1 | 4 | 4 | 1 | 10 | 8% | 18 | 14% | 2 | 2% |
| NAC | 1 | 2 | 3 | 1 | 7 | 8% | 28 | 33% | 4 | 5% |
| bZIP | 1 | 0 | 2 | 0 | 3 | 4% | 15 | 21% | 0 | 0% |
| WRKY | 2 | 3 | 7 | 2 | 14 | 21% | 31 | 46% | 12 | 18% |
| MADS | 0 | 0 | 1 | 0 | 1 | 2% | 4 | 10% | 0 | 0% |
| GRAS | 0 | 0 | 3 | 0 | 3 | 8% | 10 | 27% | 1 | 3% |
| DOF | 0 | 0 | 3 | 0 | 3 | 8% | 4 | 11% | 0 | 0% |
| HSF | 0 | 0 | 1 | 0 | 1 | 3% | 7 | 21% | 0 | 0% |
| TCP | 1 | 0 | 0 | 0 | 1 | 4% | 5 | 19% | 0 | 0% |
| GATA | 1 | 0 | 0 | 0 | 1 | 4% | 5 | 19% | 0 | 0% |
| NF-Y | 0 | 0 | 1 | 1 | 2 | 8% | 5 | 19% | 0 | 0% |
| ARF | 1 | 0 | 0 | 0 | 1 | 10% | 3 | 15% | 0 | 0% |
| HOX | 1 | 0 | 1 | 0 | 2 | 12% | 0 | 0% | 0 | 0% |
| SBP | 0 | 0 | 0 | 0 | 0 | 0% | 0 | 0% | 0 | 0% |
| others | 4 | 0 | 1 | 1 | 6 | 6% | 18 | 18% | 2 | 2% |
| Total | 28 | 32 | 46 | 13 | 119 | 10% | 241 | 19% | 46 | 4% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Bouwmeester, H.J.; Dicke, M.; Kappers, I.F. Genome-Wide Analysis Reveals Transcription Factors Regulated by Spider-Mite Feeding in Cucumber (Cucumis sativus). Plants 2020, 9, 1014. https://doi.org/10.3390/plants9081014
He J, Bouwmeester HJ, Dicke M, Kappers IF. Genome-Wide Analysis Reveals Transcription Factors Regulated by Spider-Mite Feeding in Cucumber (Cucumis sativus). Plants. 2020; 9(8):1014. https://doi.org/10.3390/plants9081014
Chicago/Turabian StyleHe, Jun, Harro J. Bouwmeester, Marcel Dicke, and Iris F. Kappers. 2020. "Genome-Wide Analysis Reveals Transcription Factors Regulated by Spider-Mite Feeding in Cucumber (Cucumis sativus)" Plants 9, no. 8: 1014. https://doi.org/10.3390/plants9081014
APA StyleHe, J., Bouwmeester, H. J., Dicke, M., & Kappers, I. F. (2020). Genome-Wide Analysis Reveals Transcription Factors Regulated by Spider-Mite Feeding in Cucumber (Cucumis sativus). Plants, 9(8), 1014. https://doi.org/10.3390/plants9081014