Molecular Mechanisms Supporting Rice Germination and Coleoptile Elongation under Low Oxygen

Abstract
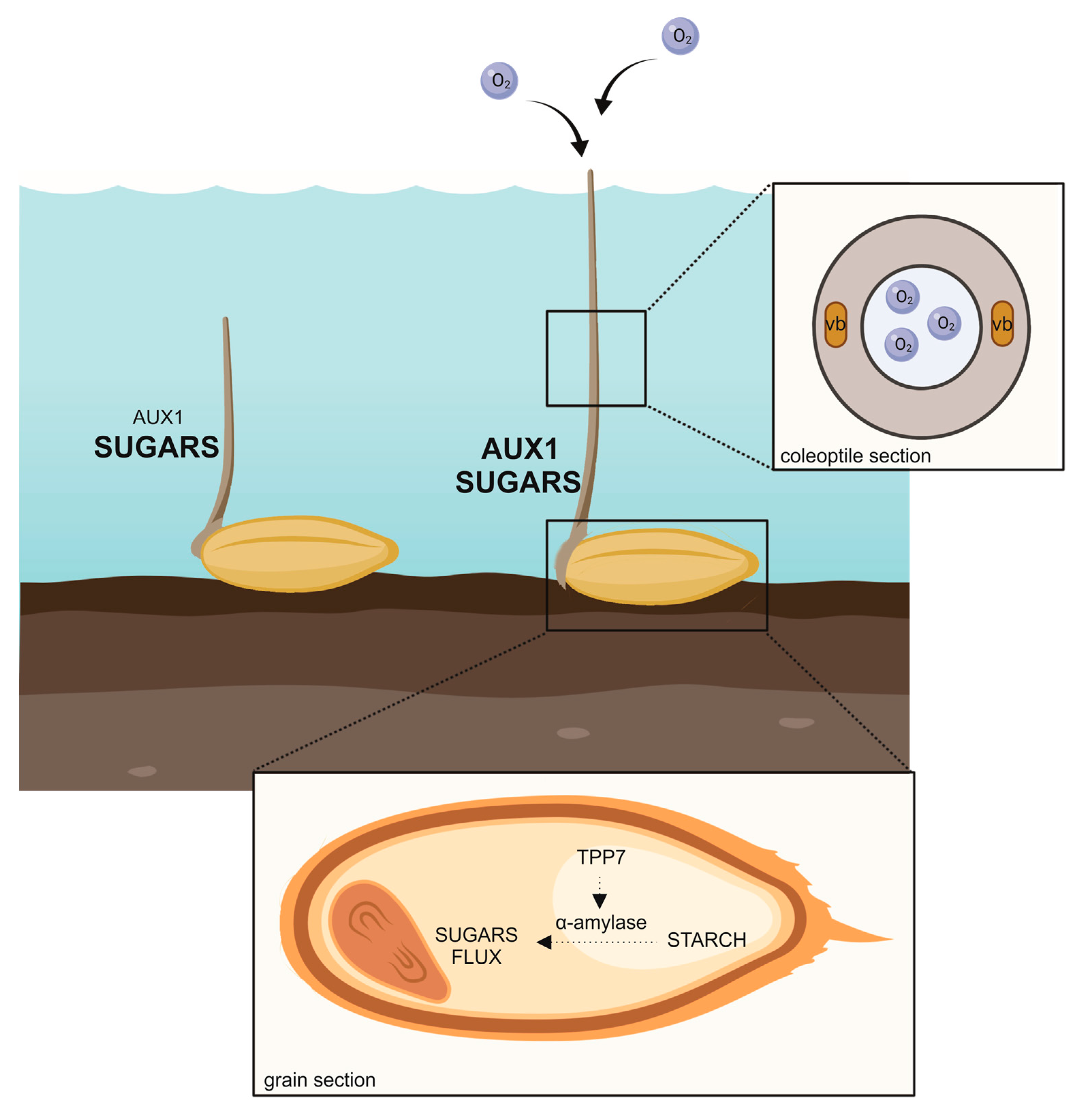
:1. Introduction
2. The Coleoptile Elongation under Submergence
3. Anaerobic Gene Regulation in Rice Coleoptile
4. Chromosomal Regions Regulating Coleoptile Elongation under Oxygen Shortage
5. Starch Degradation during Anaerobic Rice Germination
6. Hormonal Regulation of Anaerobic Rice Coleoptile Elongation
7. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Ismail, A.M.; Singh, U.S.; Singh, S.; Dar, M.H.; Mackill, D.J. The contribution of submergence-tolerant (Sub1) rice varieties to food security in flood-prone rainfed lowland areas in Asia. Field Crop. Res. 2013, 152, 83–93. [Google Scholar] [CrossRef]
- Septiningsih, E.M.; Ignacio, J.C.I.; Sendon, P.M.D.; Sanchez, D.L.; Ismail, A.M.; Mackill, D.J. QTL mapping and confirmation for tolerance of anaerobic conditions during germination derived from the rice landrace Ma-Zhan Red. Theor. Appl. Genet. 2013, 126, 1357–1366. [Google Scholar] [CrossRef]
- Singh, S.; Mackill, D.J.; Ismail, A.M. Responses of SUB1 rice introgression lines to submergence in the field: Yield and grain quality. Field Crop. Res. 2009, 113, 12–23. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Fukao, T.; Ronald, P.; Ismail, A.; Heuer, S.; Mackill, D. Submergence tolerant rice: SUB1′s journey from landrace to modern cultivar. Rice 2010, 3, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Perata, P.; Pozueta-Romero, J.; Akazawa, T.; Yamaguchi, J. Effect of anoxia on starch breakdown in rice and wheat seeds. Planta 1992, 188, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Guglielminetti, L.; Perata, P.; Alpi, A. Effect of anoxia on carbohydrate metabolism in rice seedlings. Plant Physiol. 1995, 108, 735–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasanthi-Kudahettige, R.; Magneschi, L.; Loreti, E.; Gonzali, S.; Licausi, F.; Novi, G.; Beretta, O.; Vitulli, F.; Alpi, A.; Perata, P. Transcript profiling of the anoxic rice coleoptile. Plant Physiol. 2007, 144, 218–231. [Google Scholar] [CrossRef] [Green Version]
- Loreti, E.; Alpi, A.; Perata, P. α-amylase expression under anoxia in rice seedlings: An update. Russ. J. Plant Physiol. 2003, 50, 737–742. [Google Scholar] [CrossRef]
- Fröhlich, M.; Kutschera, U. Changes in soluble sugars and proteins during development of rye coleoptiles. J. Plant Physiol. 1995, 146, 121–125. [Google Scholar] [CrossRef]
- Narsai, R.; Edwards, J.M.; Roberts, T.H.; Whelan, J.; Joss, G.H.; Atwell, B.J. Mechanisms of growth and patterns of gene expression in oxygen-deprived rice coleoptiles. Plant J. 2015, 82, 25–40. [Google Scholar] [CrossRef]
- Inada, N.; Sakai, A.; Kuroiwa, H.; Kuroiwa, T. Three-dimensional progression of programmed death in the rice coleoptile. Int. Rev. Cytol. 2002, 218, 221–258. [Google Scholar] [CrossRef] [PubMed]
- Inada, N.; Sakai, A.; Kuroiwa, H.; Kuroiwa, T. Senescence in the nongreening region of the rice (Oryza sativa) coleoptile. Protoplasma 2000, 214, 180–193. [Google Scholar] [CrossRef]
- Kawai, M.; Uchimiya, H. Coleoptile senescence in rice (Oryza sativa L.). Ann. Bot. 2000, 86, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Narsai, R.; Secco, D.; Schultz, M.D.; Ecker, J.R.; Lister, R.; Whelan, J. Dynamic and rapid changes in the transcriptome and epigenome during germination and in developing rice (Oryza sativa) coleoptiles under anoxia and re-oxygenation. Plant J. 2017, 89, 805–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, T.J.; Rost, T.L. The developmental anatomy and ultrastructure of somatic embryos from rice (Oryza sativa L.) scutellum epithelial cells. Bot. Gaz. 1989, 150, 41–49. [Google Scholar] [CrossRef]
- Takahashi, H.; Saika, H.; Matsumura, H.; Nagamura, Y.; Tsutsumi, N.; Nishizawa, N.K.; Nakazono, M. Cell division and cell elongation in the coleoptile of rice alcohol dehydrogenase 1-deficient mutant are reduced under complete submergence. Ann. Bot. 2011, 108, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Atwell, B.J.; Waters, I.; Greenway, H. The effect of oxygen and turbulence on elongation of coleoptiles of submergence-tolerant and -intolerant rice cultivars. J. Exp. Bot. 1982, 33, 1030–1044. [Google Scholar] [CrossRef]
- Kordan, H.A. Patterns of shoot and root growth in rice seedlings germinating under water. J. Appl. Ecol. 1974, 11, 685. [Google Scholar] [CrossRef]
- Nghi, K.N.; Tagliani, A.; Mariotti, L.; Weits, D.A.; Perata, P.; Pucciariello, C. Auxin is required for the long coleoptile trait in japonica rice under submergence. New Phytol. 2020. [Google Scholar] [CrossRef]
- Nghi, K.N.; Tondelli, A.; Valè, G.; Tagliani, A.; Marè, C.; Perata, P.; Pucciariello, C. Dissection of coleoptile elongation in japonica rice under submergence through integrated genome-wide association mapping and transcriptional analyses. Plant Cell Environ. 2019, 42, 1832–1846. [Google Scholar] [CrossRef]
- Magneschi, L.; Kudahettige, R.L.; Alpi, A.; Perata, P. Expansin gene expression and anoxic coleoptile elongation in rice cultivars. J. Plant Physiol. 2009, 166, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- Pucciariello, C.; Banti, V.; Perata, P. ROS signaling as common element in low oxygen and heat stresses. Plant Physiol. Biochem. 2012, 59, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, B.; Herath, V.; Wijaya, E.; Yeo, H.C.; de los Reyes, B.G.; Lee, D.Y. Patterns of cis-element enrichment reveal potential regulatory modules involved in the transcriptional regulation of anoxia response of japonica rice. Gene 2012, 511, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-K.; Tung, C.-W. RNA-Seq analysis of diverse rice genotypes to identify the genes controlling coleoptile growth during submerged germination. Front. Plant Sci. 2017, 8, 762. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.K.; Tung, C.W. Genetic mapping of anaerobic germination-associated QTLs controlling coleoptile elongation in rice. Rice 2015, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Angaji, S.A.; Septiningsih, E.M.; Mackill, D.J.; Ismail, A.M. QTLs associated with tolerance of flooding during germination in rice (Oryza sativa L.). Euphytica 2010, 172, 159–168. [Google Scholar] [CrossRef]
- Zhang, M.; Lu, Q.; Wu, W.; Niu, X.; Wang, C.; Feng, Y.; Xu, Q.; Wang, S.; Yuan, X.; Yu, H.; et al. Association mapping reveals novel genetic loci contributing to flooding tolerance during germination in indica rice. Front. Plant Sci. 2017, 8, 678. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, T.; Sasaki, K.; Yamaguchi, T.; Takahashi, H.; Junko Yamagishi, J.; Kato, Y. Detection and characterization of quantitative trait loci for coleoptile elongation under anaerobic conditions in rice. Plant Prod. Sci. 2020, 23, 374–383. [Google Scholar] [CrossRef] [Green Version]
- Kretzschmar, T.; Pelayo, M.A.F.; Trijatmiko, K.R.; Gabunada, L.F.M.; Alam, R.; Jimenez, R.; Mendioro, M.S.; Slamet-Loedin, I.H.; Sreenivasulu, N.; Bailey-Serres, J.; et al. A trehalose-6-phosphate phosphatase enhances anaerobic germination tolerance in rice. Nat. Plants 2015, 1, 15124. [Google Scholar] [CrossRef]
- Biscarini, F.; Cozzi, P.; Casella, L.; Riccardi, P.; Vattari, A.; Orasen, G.; Perrini, R.; Tacconi, G.; Tondelli, A.; Biselli, C.; et al. Genome-wide association study for traits related to plant and grain morphology, and root architecture in temperate rice accessions. PLoS ONE 2016, 11, e0155425. [Google Scholar] [CrossRef]
- Hwang, Y.S.; Thomas, B.R.; Rodriguez, R.L. Differential expression of rice α-amylase genes during seedling development under anoxia. Plant Mol. Biol. 1999, 40, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Chen, P.; Lu, C.; Chen, S.; Ho, T.D.; Yu, S. Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding. Sci. Signal. 2009, 2, ra61. [Google Scholar] [CrossRef] [PubMed]
- Weinl, S.; Kudla, J. The CBL-CIPK Ca2+-decoding signaling network: Function and perspectives. New Phytol. 2009, 184, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Ho, V.T.; Tran, A.N.; Cardarelli, F.; Perata, P.; Pucciariello, C. A calcineurin B-like protein participates in low oxygen signalling in rice. Funct. Plant Biol. 2017, 44, 917–928. [Google Scholar] [CrossRef]
- Ye, N.H.; Wang, F.Z.; Shi, L.; Chen, M.X.; Cao, Y.Y.; Zhu, F.Y.; Wu, Y.Z.; Xie, L.J.; Liu, T.Y.; Su, Z.Z.; et al. Natural variation in the promoter of rice calcineurin B-like protein10 (OsCBL10) affects flooding tolerance during seed germination among rice subspecies. Plant J. 2018, 94, 612–625. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Lim, E.; Yu, S. Sugar response sequence in the promoter of a rice a-Amylase gene. J. Biolo. Chem. 1998, 273, 10120–10131. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.A.; Ho, T.H.; Ho, S.L.; Yu, S.M. Three novel MYB proteins with one DNA binding repeat mediate sugar and hormone regulation of alpha-amylase gene expression. Plant Cell 2002, 14, 1963–1980. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.-A.; Lin, C.-C.; Lee, K.-W.; Chen, J.-L.; Huang, L.-F.; Ho, S.-L.; Liu, H.-J.; Hsing, Y.-I.; Yu, S.-M. The SnRK1A protein kinase plays a key role in sugar signaling during germination and seedling growth of rice. Plant Cell 2007, 19, 2484–2499. [Google Scholar] [CrossRef] [Green Version]
- Magneschi, L.; Kudahettige, R.L.; Alpi, A.; Perata, P. Comparative analysis of anoxic coleoptile elongation in rice varieties: Relationship between coleoptile length and carbohydrate levels, fermentative metabolism and anaerobic gene expression. Plant Biol. 2009, 11, 561–573. [Google Scholar] [CrossRef]
- Takahashi, H.; Greenway, H.; Matsumura, H.; Tsutsumi, N.; Nakazono, M. Rice alcohol dehydrogenase 1 promotes survival and has a major impact on carbohydrate metabolism in the embryo and endosperm when seeds are germinated in partially oxygenated water. Ann. Bot. 2014, 113, 851–859. [Google Scholar] [CrossRef] [Green Version]
- Nagao, M.; Ohwaki, Y. The action of trans-cinnamic and 2,3,5-triiodobenzoic acids in the rice seedling. Sci. Rep. Tohoku Univ. 1955, 21, 96–108. [Google Scholar]
- Kefford, N.P. Auxin-gibberellin interaction in rice coleoptile elongation. Plant Physiol. 1962, 37, 380–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegoraro, R.; Mapelli, S.; Torti, G.; Bertani, A. Indole-3-acetic acid and rice coleoptile elongation under anoxia. J. Plant Growth Regul. 1988, 7, 85–94. [Google Scholar] [CrossRef]
- Horton, R.F. The effect of ethylene and other regulators on coleoptile growth of rice under anoxia. Plant Sci. 1991, 79, 57–62. [Google Scholar] [CrossRef]
- Breviario, D.; Giani, S.; Di Vietri, P.; Coraggio, I. Auxin and growth regulation of rice coleoptile segments: Molecular analysis. Plant Physiol. 1992, 98, 488–495. [Google Scholar] [CrossRef]
- Hoson, T.; Masuda, Y.; Pilet, P.E. Auxin content in air and water grown rice coleoptiles. J. Plant Physiol. 1992, 139, 685–689. [Google Scholar] [CrossRef]
- Guo, F.; Han, N.; Xie, Y.; Fang, K.; Yang, Y.; Zhu, M.; Wang, J.; Bian, H. The miR393a/target module regulates seed germination and seedling establishment under submergence in rice (Oryza sativa L.). Plant Cell Environ. 2016, 39, 2288–2302. [Google Scholar] [CrossRef]
- Ismail, A.M.; Ella, E.S.; Vergara, G.V.; Mackill, D.J. Mechanisms associated with tolerance to flooding during germination and early seedling growth in rice (Oryza sativa). Ann. Bot. 2009, 103, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Hartman, S.; Liu, Z.; van Veen, H.; Vicente, J.; Reinen, E.; Martopawiro, S.; Zhang, H.; van Dongen, N.; Bosman, F.; Bassel, G.W.; et al. Ethylene-mediated nitric oxide depletion pre-adapts plants to hypoxia stress. Nat. Commun. 2019, 10, 4020. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Ma, B.; He, S.J.; Xiong, Q.; Duan, K.X.; Yin, C.C.; Chen, H.; Lu, X.; Chen, S.Y.; Zhang, J.S. MAOHUZI6/ETHYLENE INSENSITIVE3-LIKE1 and ETHYLENE INSENSITIVE3-LIKE2 regulate ethylene response of roots and coleoptiles and negatively affect salt tolerance in rice. Plant Physiol. 2015, 169, 148–165. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, D.J.; Lee, S.C.; Md Isa, N.; Gramuglia, S.; Fukao, T.; Bassel, G.W.; Correia, C.S.; Corbineau, F.; Theodoulou, F.L.; Bailey-Serres, J.; et al. Homeostatic response to hypoxia is regulated by the N-end rule pathway in plants. Nature 2011, 479, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Chao, Y.T.; Chen, W.C.; Ho, H.Y.; Chou, M.Y.; Li, Y.R.; Wu, Y.L.; Yang, H.A.; Hsieh, H.; Lin, C.S.; et al. Regulatory cascade involving transcriptional and N-end rule pathways in rice under submergence. Proc. Natl. Acad. Sci. USA 2019, 116, 3300–3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, H.; Zhang, Z.; Wang, J.; Chen, X.; Wei, P.; Huang, R. The activation of OsEIL1 on YUC8M transcription and auxin biosynthesis is required for ethylene-inhibited root elongation in rice early seedling development. PLoS Genet. 2017, 13, e1006955. [Google Scholar] [CrossRef]
- Ishizawa, K.; Esashi, Y. Osmoregulation in rice coleoptile elongation as promoted by cooperation between IAA and ethylene. Plant Cell Physiol. 1984, 25, 495–504. [Google Scholar] [CrossRef]


{kind=link}
| Trait of Study | Genotypes | Major Chromosomal Region | Reference |
|---|---|---|---|
| Tolerance to flooding during germination | Khao Hlan On backcross population with IR64 recurrent parent | Chr 9 qAG-9-1 and qAG-9-2 QTLs | Angaji et al., 2010 [26] |
| Tolerance to anaerobic conditions during germination | Population derived from a cross between Ma-Zhan Red and IR42 | Chr 7 qAG-7-1 QTL | Septiningsih et al., 2013 [2] |
| Coleoptile elongation under anaerobic germination | Recombinant inbred line population derived from a cross between japonica and indica varieties | Chr1 QTL | Hsu and Tung, 2015 [24] |
| Coleoptile length during germination under flooding | Panel of 432 indica rice varieties | Chr 6 MTAs | Zhang et al., 2017 [27] |
| Coleoptile length under dark submergence | Panel of 273 japonica rice accessions | Chr1, Chr5 MTAs | Nghi et al., 2019 [20] |
| Coleoptile length in anaerobic solution | 39 chromosome segment substitution lines derived from a cross between Koshihikari and IR64 rice varieties | Chr3 qACE3.1 | Noshimura et al., 2020 [28] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pucciariello, C. Molecular Mechanisms Supporting Rice Germination and Coleoptile Elongation under Low Oxygen. Plants 2020, 9, 1037. https://doi.org/10.3390/plants9081037
Pucciariello C. Molecular Mechanisms Supporting Rice Germination and Coleoptile Elongation under Low Oxygen. Plants. 2020; 9(8):1037. https://doi.org/10.3390/plants9081037
Chicago/Turabian StylePucciariello, Chiara. 2020. "Molecular Mechanisms Supporting Rice Germination and Coleoptile Elongation under Low Oxygen" Plants 9, no. 8: 1037. https://doi.org/10.3390/plants9081037
APA StylePucciariello, C. (2020). Molecular Mechanisms Supporting Rice Germination and Coleoptile Elongation under Low Oxygen. Plants, 9(8), 1037. https://doi.org/10.3390/plants9081037