
Acclimation Changes of Flavonoids in Needles of Conifers during Heat and Drought Stress 2015


Abstract
:1. Introduction
2. Materials and Methods
2.1. Climate Conditions of 2015
2.2. Plant Material
2.3. Histology
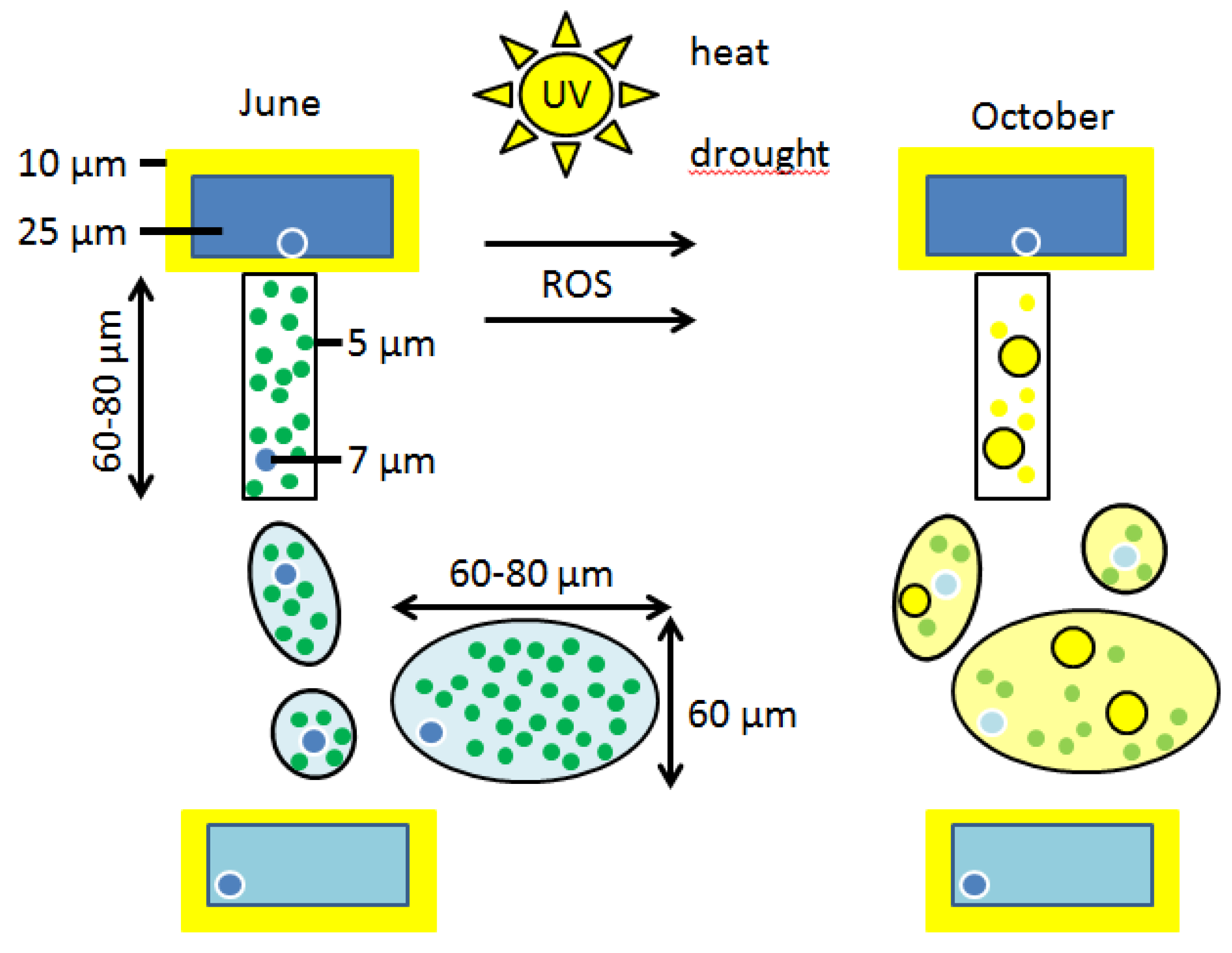
3. Results
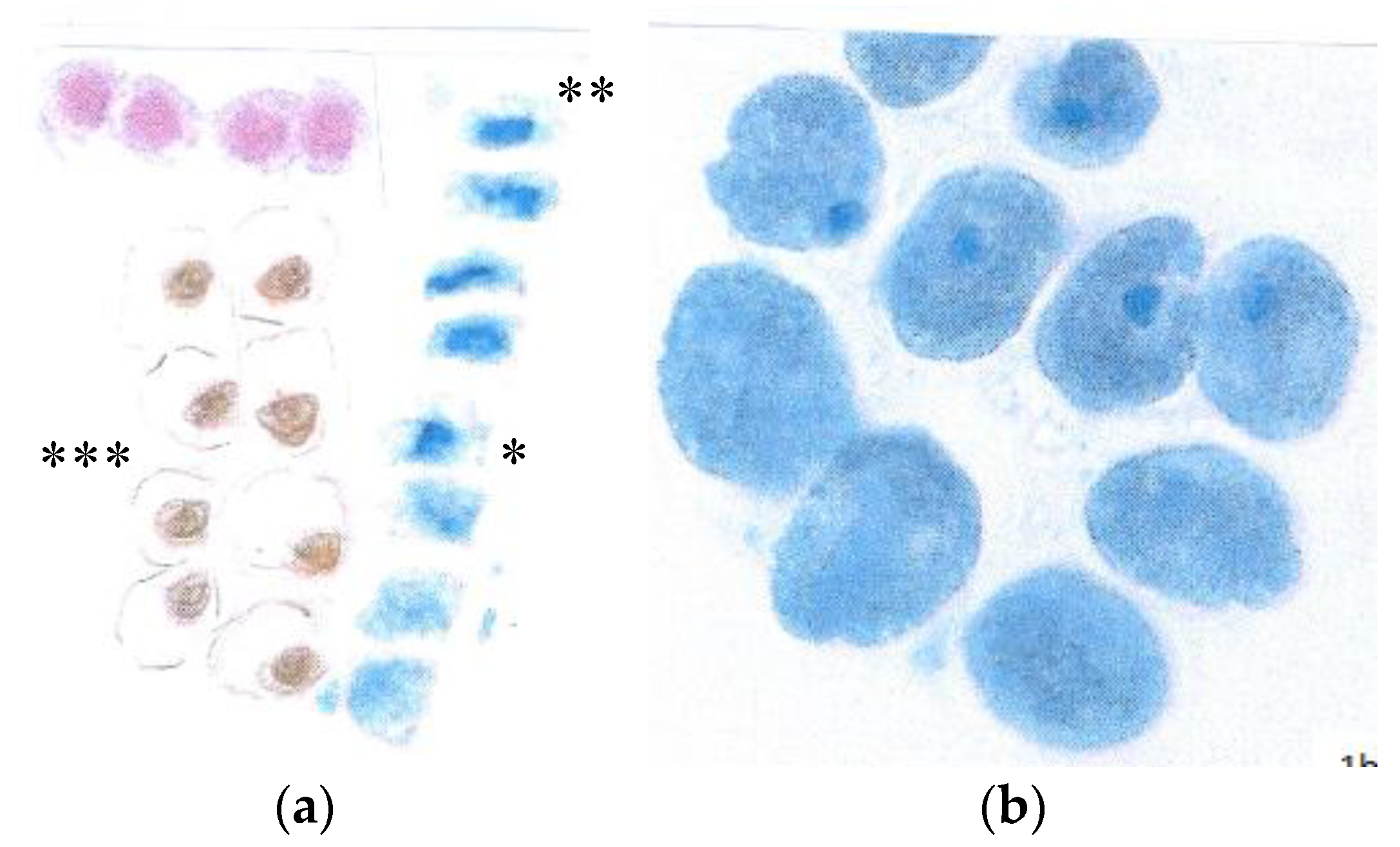
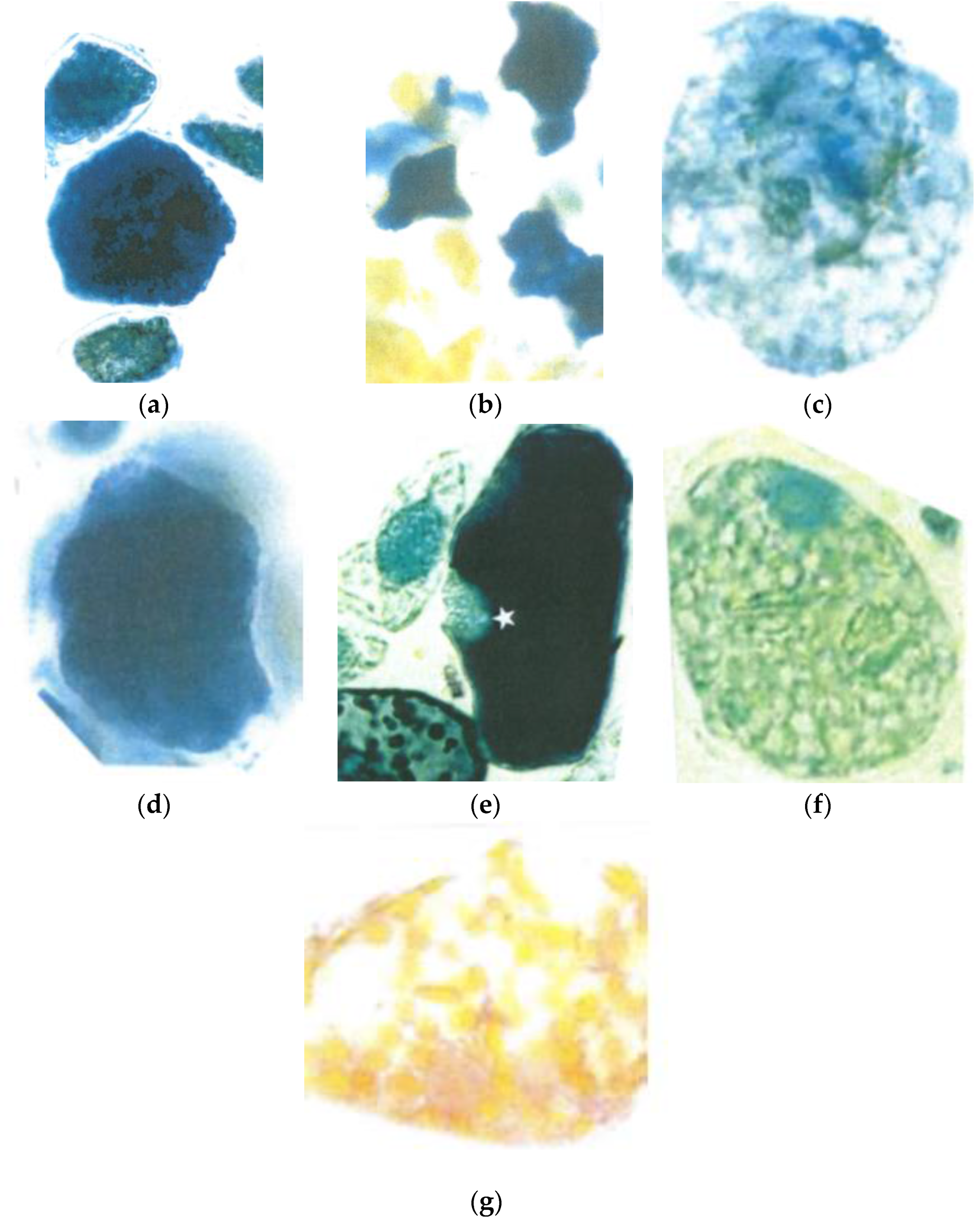
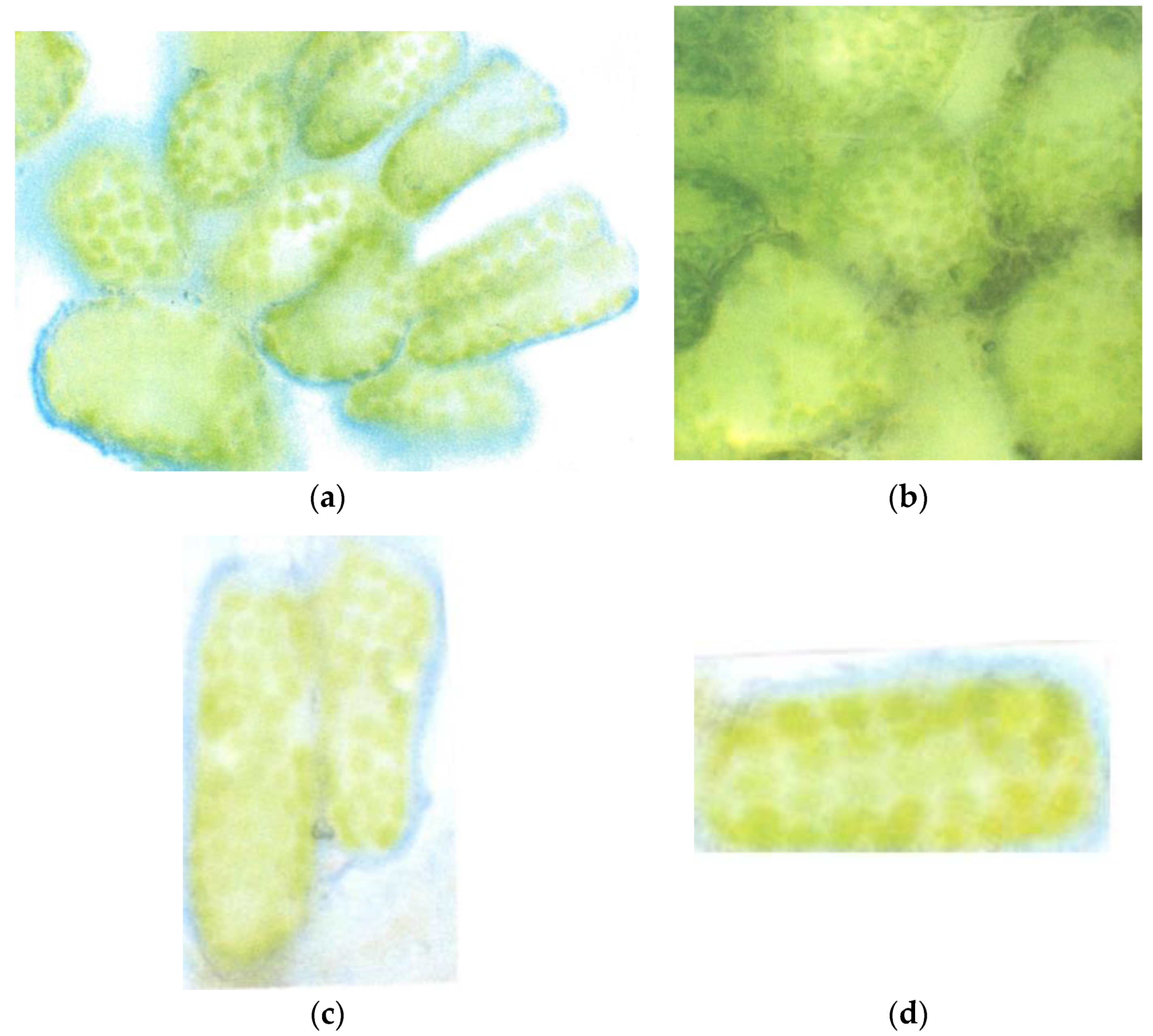
3.1. Loss of the Optimal Nuclear Assembly from May to August
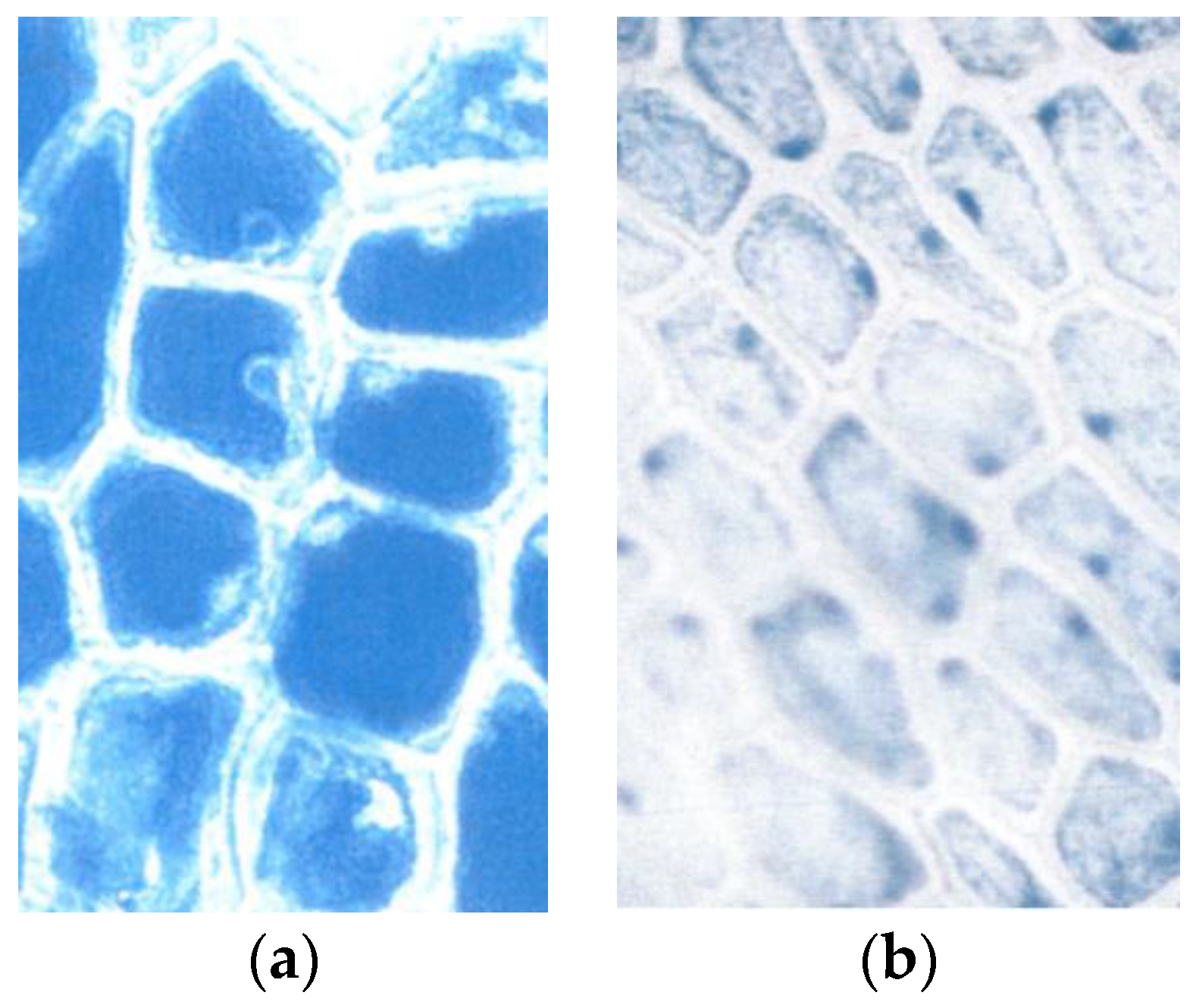
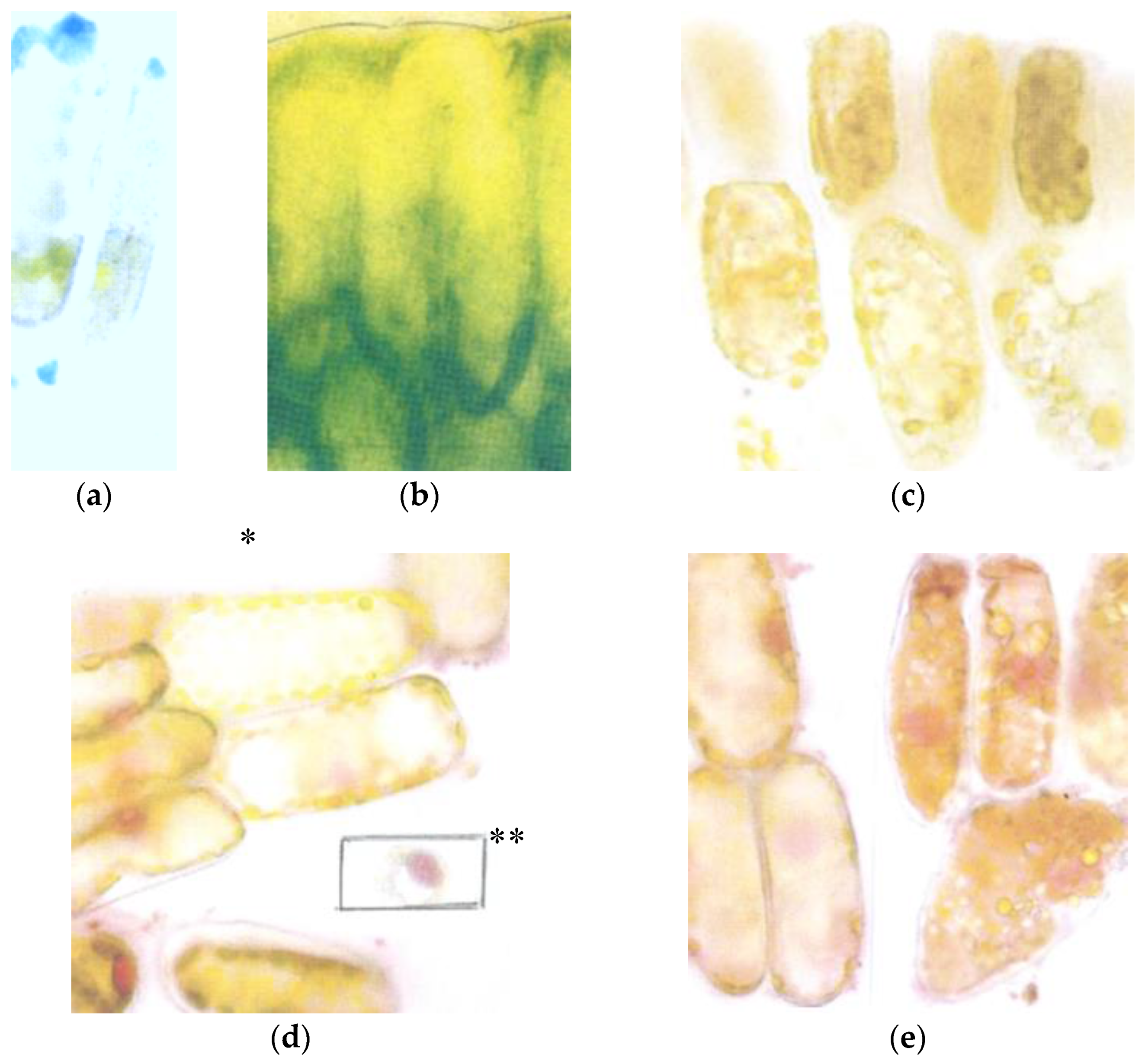
3.2. Early Breakdown of the Epidermal Flavanol Barrier against UV-Radiation
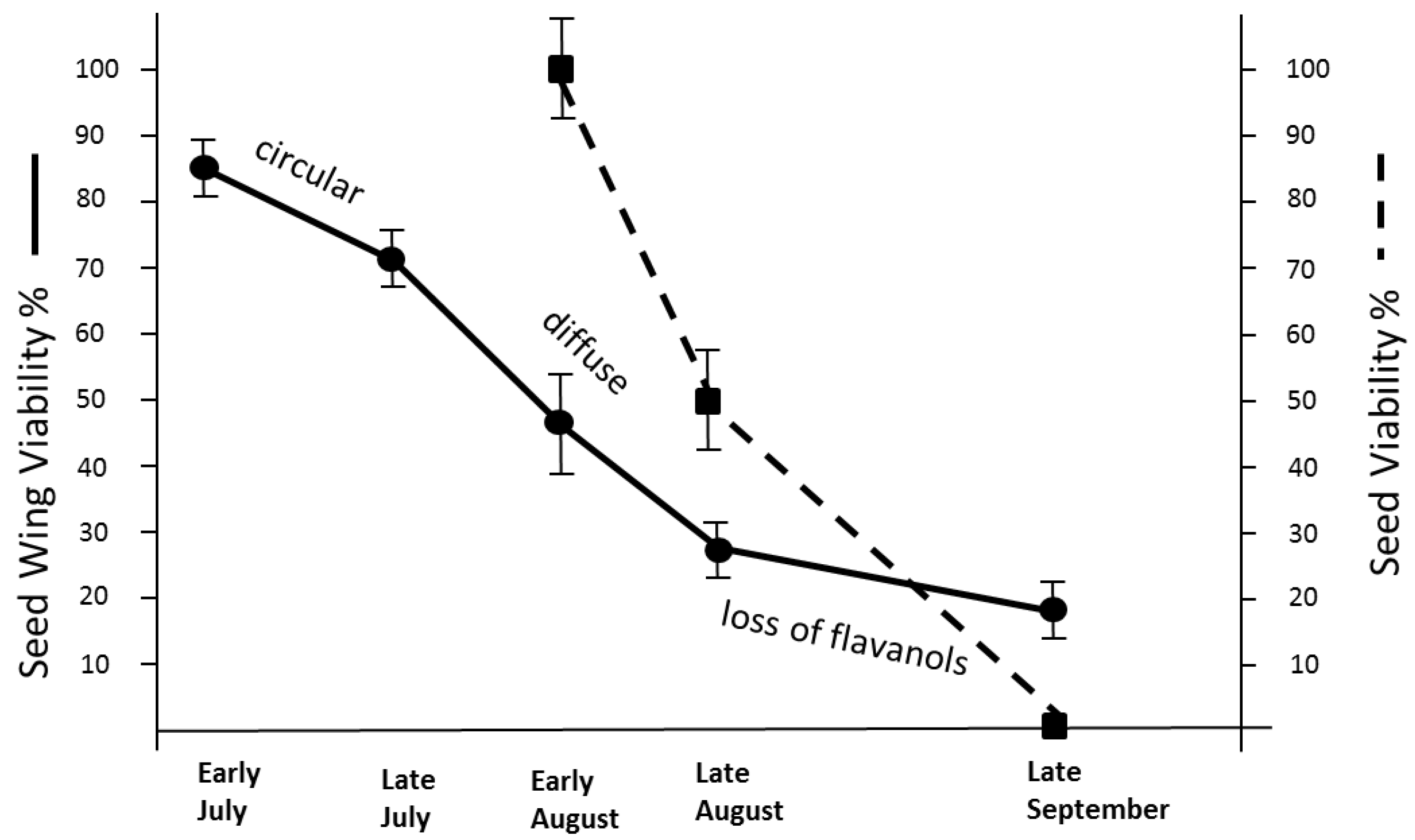
3.3. Early Breakdown in July of the Young Seed Wings from Tsuga
3.4. Loss of the Flavanols Promotes the Onset of Intracellular Structural Mismatch
3.5. First Signs of Breakdown of the Epidermal Flavanols by Mid-July
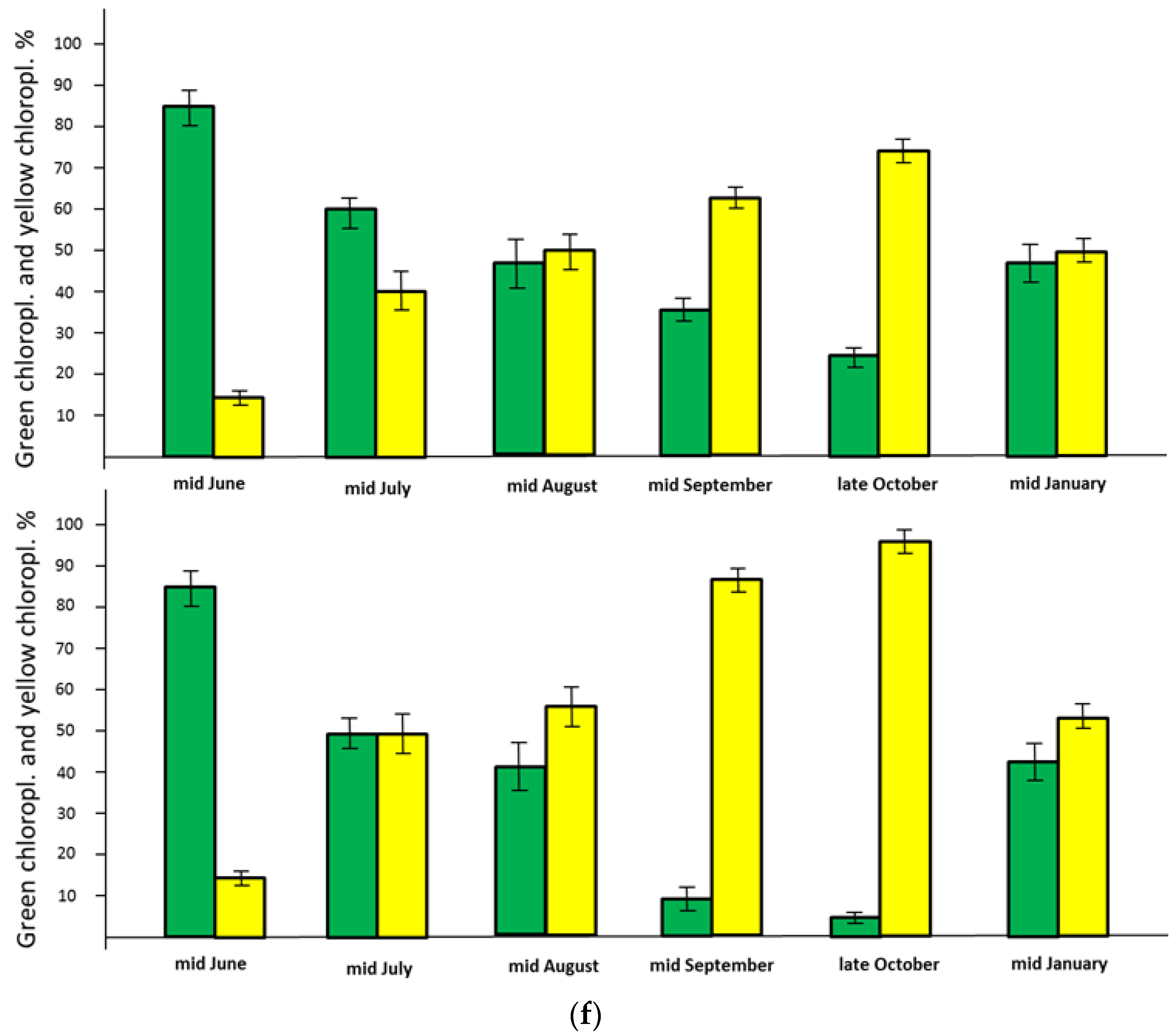
3.6. Very Early Decrease of Green Chloroplasts of the Palisade Cells in Late July
3.7. Retarded Breakdown of the Flavanols from the Spongy Mesophyll in Late August
3.8. The Curious Restoration of Green Chloroplasts in January 2016
4. Discussion
4.1. Early Beginning of Cellular Disorganization in June 2015
4.2. Flavanols Guarantee an Antioxidative Protection of the Overstresses Mesophyll Chloroplasts
4.3. Flavanoid Deposition in Severely Stressed Needles
4.4. Restoration of the Chloroplasts in Mid-Winter 2015/2016
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Feucht, W.; Polster, J. Nuclei of plants as a sink for flavanols. Z. Naturforsch. C 2001, 56, 479–481. [Google Scholar] [PubMed]
- Fey, V.; Wagner, R.; Bräutigam, K.; Pfannschmidt, T. Photosynthetic redox control of nuclear gene expression. J. Exp. Bot. 2005, 56, 1491–1498. [Google Scholar] [CrossRef] [PubMed]
- Mueller-Harvey, I.; Feucht, W.; Polster, J.; Trnkova, L.; Burgos, P.; Parker, A.W.; Botchway, S.W. Two-photon excitation with pico-second fluorescence lifetime imaging to detect nuclear association of flavanols. Anal. Chim. Acta 2012, 719, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Bors, W.; Michel, C. Antioxidant capacity of flavanols and gallate esters: Pulse radiolysis studies. Free Radic. Biol. Med. 1999, 27, 1413–1426. [Google Scholar] [CrossRef]
- Leshem, Y.A.Y.; Halevy, A.H.; Frenkel, C. Processes and Control of Plant Senescence; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Gunthardt-Goerg, M.S.; Vollenweider, P. Linking stress with macroscopic and microscopic leaf response in trees: New diagnostic perspectives. Environ. Pollut. 2007, 147, 467–488. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Matteini, P.; Goti, A.; Tattini, M. Chloroplast-located flavonoids can scavenge singlet oxygen. New Phytol. 2007, 174, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Stefano, G.; Biricolti, S.; Tattini, M. Mesophyll distribution of “antioxidant“ flavonoid glycosides in ligustrum vulgare leaves under contrasting sunlight irradiance. Ann. Bot. 2009, 104, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Rohn, S.; Rawel, H.M.; Kroll, J. Antioxidant activity of protein-bound quercetin. J. Agric. Food Chem. 2004, 52, 4725–4729. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, C.; Di Ferdinando, M.; Fini, A.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants and developmental regulators: Relative significance in plants and humans. Int. J. Mol. Sci. 2013, 14, 3540–3555. [Google Scholar] [CrossRef] [PubMed]
- Feucht, W.; Treutter, D.; Dithmar, H.; Polster, J. Loss of nuclear flavanols during drought periods in taxus baccata. Plant Biol. 2013, 15, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Feucht, W.; Schmid, M.; Treutter, D. Nuclei of Tsuga canadensis: Role of flavanols in chromatin organization. Int. J. Mol. Sci. 2011, 12, 6834–6855. [Google Scholar] [CrossRef] [PubMed]
- Treutter, D. Chemical reaction detection of catechins and proanthocyanidins with 4-dimethylaminocinnamaldehyde. J. Chromatogr. A 1989, 467, 185–193. [Google Scholar] [CrossRef]
- Feucht, W.; Treutter, D.; Dithmar, H.; Polster, J. Microspore development of three coniferous species: Affinity of nuclei for flavonoids. Tree Physiol. 2008, 28, 1783–1791. [Google Scholar] [CrossRef] [PubMed]
- Krauze-Baranowska, M. Flavonoids from the genus taxus. Z. Naturforsch. C 2004, 59, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Feucht, W.; Treutter, D.; Polster, J. Flavanols in nuclei of tree species: Facts and possible functions. Trees-Struct. Funct. 2012, 26, 1413–1425. [Google Scholar] [CrossRef]
- Feucht, W.; Treutter, D.; SantosBuelga, C.; Christ, E. Catechin as a radical scavenger in paraquat-treated prunus avium. J. Appl. Bot.-Angew. Bot. 1996, 70, 119–123. [Google Scholar]
- Ries, G.; Heller, W.; Puchta, H.; Sandermann, H.; Seidlitz, H.K.; Hohn, B. Elevated UV-B radiation reduces genome stability in plants. Nature 2000, 406, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Solovchenko, A.; Schmitz-Eiberger, M. Significance of skin flavonoids for uv-b-protection in apple fruits. J. Exp. Bot. 2003, 54, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Feucht, W.; Treutter, D. Effects of abscisic acid and (+)-catechin on growth and leaching properties of callus from four fruit tree species. Gartenbauwissenschaft (Germany) 1996, 61, 174–178. [Google Scholar]
- D'Angelo, M.A.; Raices, M.; Panowski, S.H.; Hetzer, M.W. Age-dependent deterioration of nuclear pore complexes causes a loss of nuclear integrity in postmitotic cells. Cell 2009, 136, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Vigh, L.; Huitema, H.; Woltjes, J.; van Hasselt, P.R. Drought stress-induced changes in the composition and physical state of phospholipids in wheat. Physiol. Plant. 1986, 67, 92–96. [Google Scholar] [CrossRef]
- Dubas, E.; Janowiak, F.; Krzewska, M.; Hura, T.; Żur, I. Endogenous aba concentration and cytoplasmic membrane fluidity in microspores of oilseed rape (brassica napus L.) genotypes differing in responsiveness to androgenesis induction. Plant Cell Rep. 2013, 32, 1465–1475. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Plant secondary metabolism: Diversity, function and its evolution. Nat. Prod. Commun. 2008, 3, 1205–1216. [Google Scholar]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. Ros signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Jordan, D.; Smith, W. Simulated influence of leaf geometry on sunlight interception and photosynthesis in conifer needles. Tree Physiol. 1993, 13, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Voltas, J.; Chambel, M.R.; Prada, M.A.; Ferrio, J.P. Climate-related variability in carbon and oxygen stable isotopes among populations of aleppo pine grown in common-garden tests. Trees 2008, 22, 759–769. [Google Scholar] [CrossRef]
- Strand, Å. Plastid-to-nucleus signalling. Curr.Opin. Plant Biol. 2004, 7, 621–625. [Google Scholar]
- Beck, C.F. Signaling pathways from the chloroplast to the nucleus. Planta 2005, 222, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R. Role of plant polyphenols in genomic stability. Mutat. Res. 2001, 475, 89–111. [Google Scholar] [CrossRef]
- Kefeli, V.; Kutacek, M. Phenolic substances and their possible role in plant growth regulation. In Plant Growth Regulation; Springer: Berlin, Germany, 1977; pp. 181–188. [Google Scholar]
- Ackland, M.L.; Van de Warsenburg, S.; Jones, R. Synergistic antiproliferative action of the flavonols quercetin and kaempferol in cultured human cancer cell lines. In Vivo 2005, 19, 69–76. [Google Scholar] [PubMed]
- Peer, W.; Murphy, A.S. Flavonoids as signal molecules: Targets of flavonoid action. In The Science of Flavonoids; Springer: Berlin, Germany, 2006; pp. 239–268. [Google Scholar]
- Khan, M.A.M.; Ulrichs, C.; Mewis, I. Effect of water stress and aphid herbivory on flavonoids in broccoli (brassica oleracea var. Italica plenck). J. Appl. Bot. Food Qual. Angew. Bot. 2011, 84, 178–182. [Google Scholar]
- Niinemets, Ü.; Lukjanova, A.; Turnbull, M.H.; Sparrow, A.D. Plasticity in mesophyll volume fraction modulates light-acclimation in needle photosynthesis in two pines. Tree Physiol. 2007, 27, 1137–1151. [Google Scholar] [CrossRef] [PubMed]

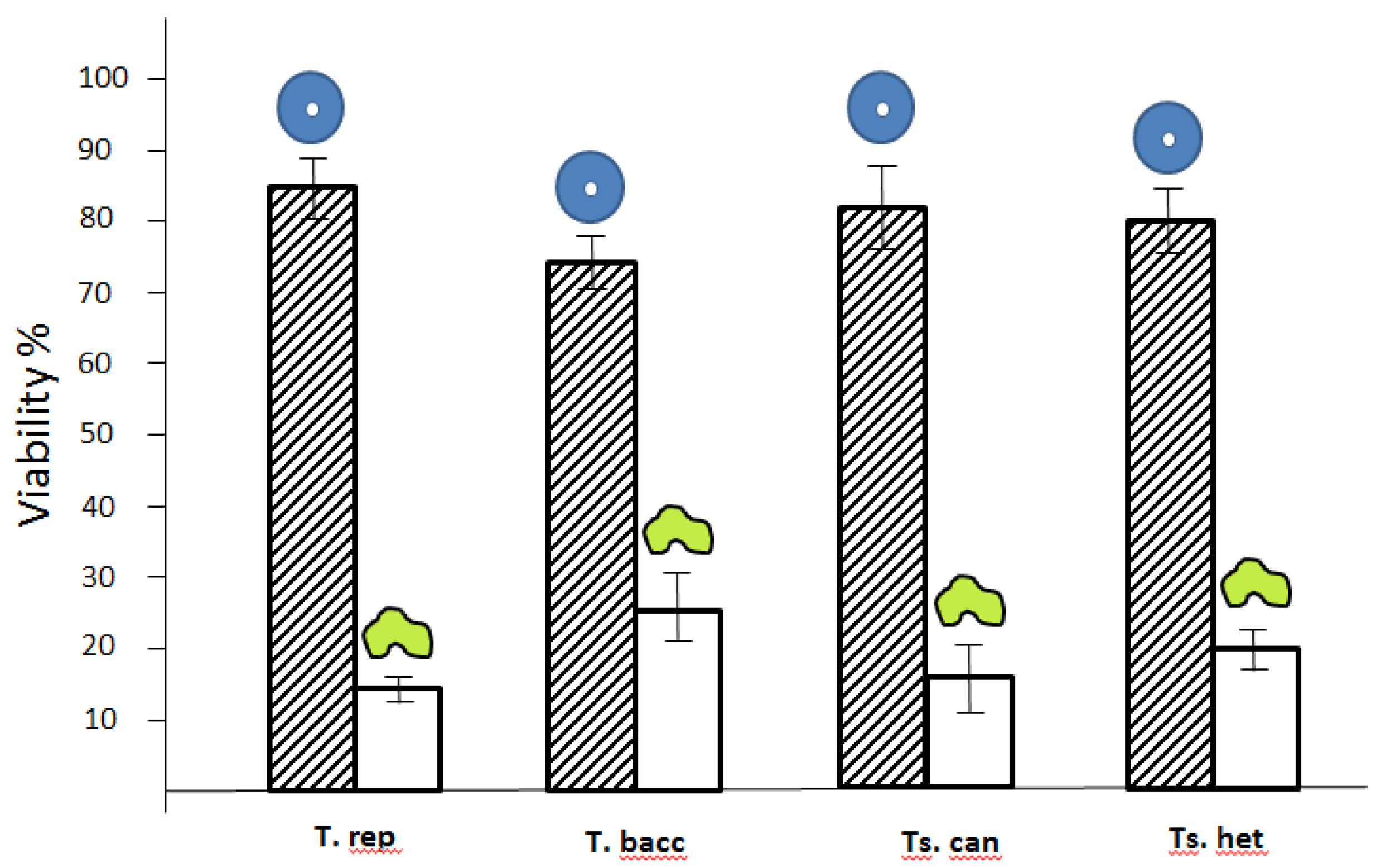
 = high viability
= high viability  = low viability with T. rep (Taxus repens), T. bacc (Taxus baccata), Ts. can (Tsuga cannadensis) and Ts. het (Tsuga heterophilla).
= high viability = low viability with T. rep (Taxus repens), T. bacc (Taxus baccata), Ts. can (Tsuga cannadensis) and Ts. het (Tsuga heterophilla).
= low viability with T. rep (Taxus repens), T. bacc (Taxus baccata), Ts. can (Tsuga cannadensis) and Ts. het (Tsuga heterophilla).
= high viability = low viability with T. rep (Taxus repens), T. bacc (Taxus baccata), Ts. can (Tsuga cannadensis) and Ts. het (Tsuga heterophilla).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| aT °C | Max T °C | aR mm | Max R mm | |
|---|---|---|---|---|
| March | 10 | 1 × 17 | 20 | 1 × 8 |
| April | 15 | 3 × 22 | 49 | 1 × 24 |
| May | 19 | 2 × 27 | 102 | 2 × 24 |
| June | 21 | 3 × 29 | 38 | 2 × 21 |
| July | 27 | 7 × 34 | 20 | 2 × 5 |
| August | 24 | 12 × 32 | 44 | 1 × 21 |
| September | 19 | 1 × 28 | 33 | 1 × 8 |
| October | 11 | 2 × 21 | 61 | 3 × 13 |
| November | 11 | 15 × 17 | 57 | 1 × 53 |
| December | 8 | 3 × 12 | 15 | 1 × 5 |
| Start of Decline Tissue 1 | Start of Decline Month 2 | Velocity of Decline Tissue 3 | Intensity of Decline Genotype 4 |
|---|---|---|---|
| 1. Upper epidermis | early July | medium | Tsu can. strong |
| 2. Palisade cells | late July | very high | Tsu het. medium |
| 3. Mesophyll cells | late August | high | Tax back. medium |
| 4. Lower epidermis | late August | medium | Tax rep. low |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feucht, W.; Schmid, M.; Treutter, D. Acclimation Changes of Flavonoids in Needles of Conifers during Heat and Drought Stress 2015. Climate 2016, 4, 35. https://doi.org/10.3390/cli4030035
Feucht W, Schmid M, Treutter D. Acclimation Changes of Flavonoids in Needles of Conifers during Heat and Drought Stress 2015. Climate. 2016; 4(3):35. https://doi.org/10.3390/cli4030035
Chicago/Turabian StyleFeucht, Walter, Markus Schmid, and Dieter Treutter. 2016. "Acclimation Changes of Flavonoids in Needles of Conifers during Heat and Drought Stress 2015" Climate 4, no. 3: 35. https://doi.org/10.3390/cli4030035
APA StyleFeucht, W., Schmid, M., & Treutter, D. (2016). Acclimation Changes of Flavonoids in Needles of Conifers during Heat and Drought Stress 2015. Climate, 4(3), 35. https://doi.org/10.3390/cli4030035