



Proteoform Analysis of the Human Olfactory System: A Window into Neurodegenerative Diseases

 , ,
, , 
Abstract
:1. Introduction
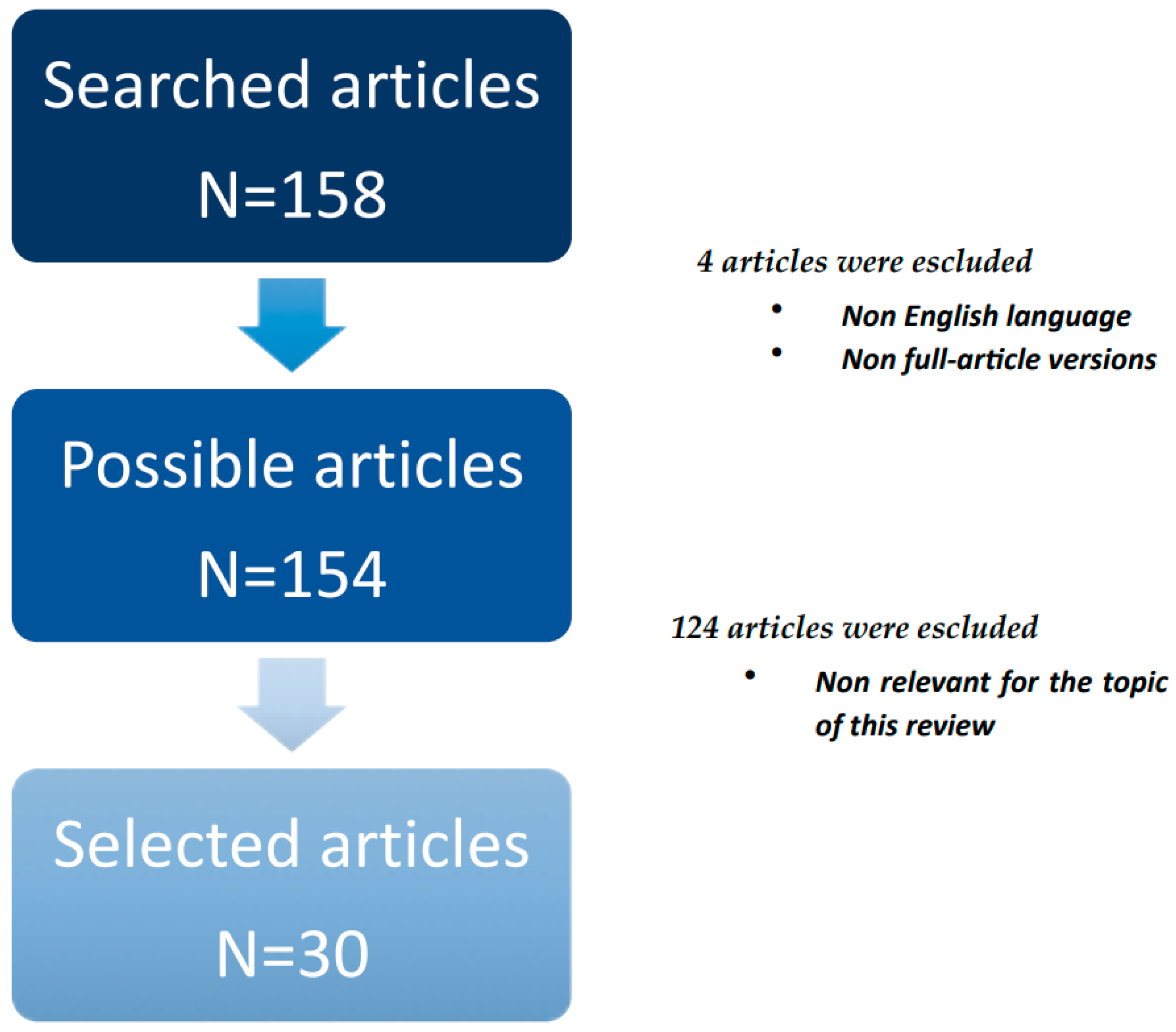
2. Materials and Methods
3. Results
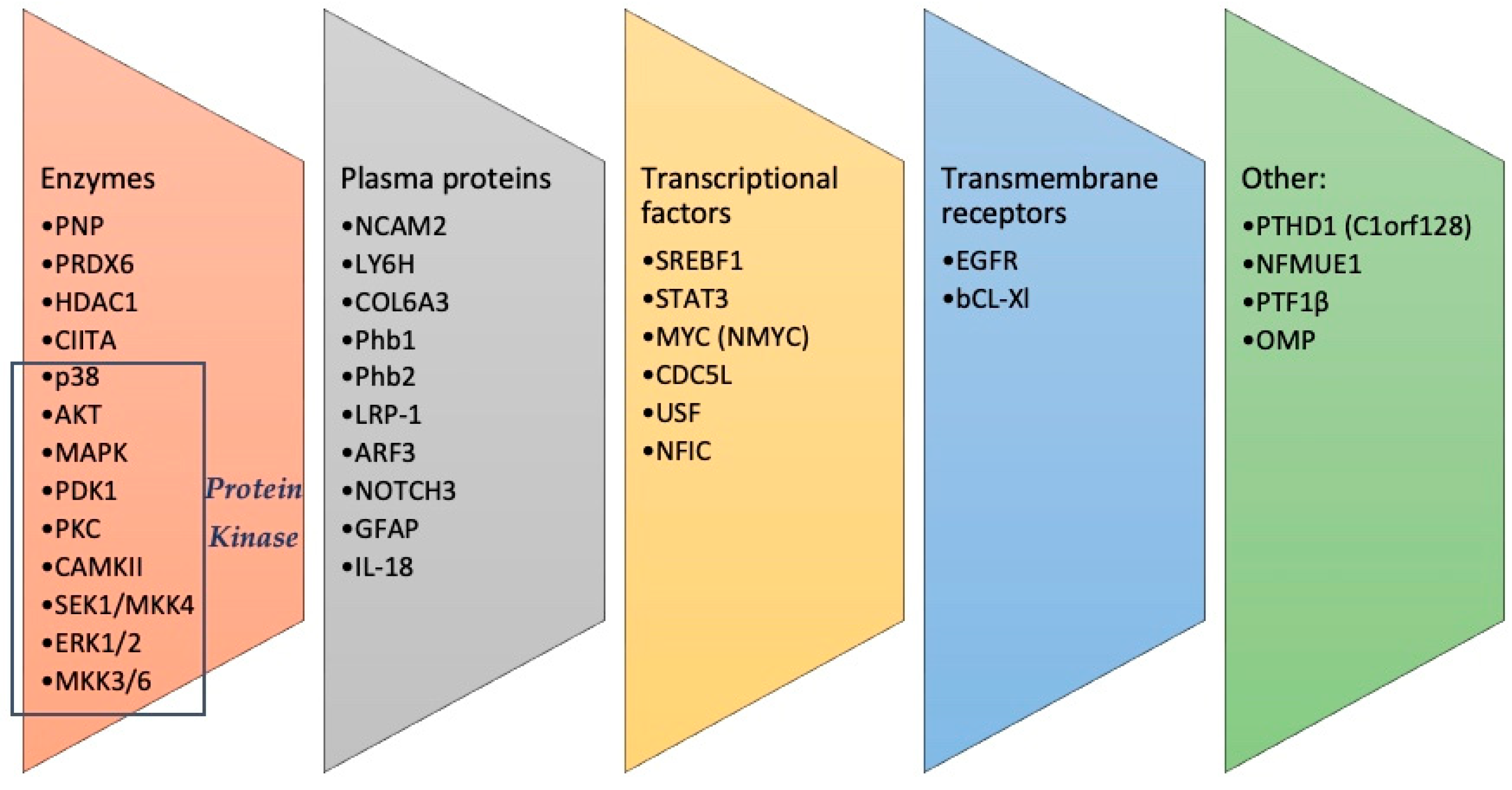
3.1. Olfactory Proteomic
3.2. Proteomic Studies on SARS-CoV-2 Associated Olfactory Dysfunctions
3.3. Proteomic Studies on Neurodegenerative Diseases Associated with Olfactory Dysfunction
3.3.1. Alzheimer’s Disease
3.3.2. Frontotemporal Dementias
3.3.3. Lewy-Type α-Synucleinopathies
3.3.4. Mixed Dementia
3.3.5. Amyotrophic Lateral Sclerosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fatuzzo, I.; Niccolini, G.F.; Zoccali, F.; Cavalcanti, L.; Bellizzi, M.G.; Riccardi, G.; de Vincentiis, M.; Fiore, M.; Petrella, C.; Minni, A.; et al. Neurons, Nose, and Neurodegenerative Diseases: Olfactory Function and Cognitive Impairment. Int. J. Mol. Sci. 2023, 24, 2117. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Irigoyen, J.; Corrales, F.J.; Santamaría, E. Proteomic atlas of the human olfactory bulb. J. Proteom. 2012, 75, 4005–4016. [Google Scholar] [CrossRef] [PubMed]
- Sarnat, H.B.; Yu, W. Maturation and Dysgenesis of the Human Olfactory Bulb. Brain Pathol. 2016, 26, 301–318. [Google Scholar] [CrossRef]
- Sarnat, H.B.; Flores-Sarnat, L. Survey on Olfactory Testing by Pediatric Neurologists: Is the Olfactory a “True” Cranial Nerve? J. Child Neurol. 2020, 35, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Kobayakawa, K.; Kobayakawa, R.; Matsumoto, H.; Oka, Y.; Imai, T.; Ikawa, M.; Okabe, M.; Ikeda, T.; Itohara, S.; Kikusui, T.; et al. Innate versus learned odour processing in the mouse olfactory bulb. Nature 2007, 450, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Kobayakawa, R.; Kobayakawa, K.; Emura, M.; Itohara, S.; Kizumi, M.; Hamana, H.; Tsuboi, A.; Hirono, J. Supersensitive detection and discrimination of enantiomers by dorsal olfactory receptors: Evidence for hierarchical odour coding. Sci. Rep. 2015, 5, 14073. [Google Scholar] [CrossRef] [PubMed]
- Schweyer, K.; Rüschoff-Steiner, C.; Arias-Carrión, O.; Oertel, W.H.; Rösler, T.W.; Höglinger, G.U. Neuronal precursor cells with dopaminergic commitment in the rostral migratory stream of the mouse. Sci. Rep. 2019, 9, 13359. [Google Scholar] [CrossRef] [PubMed]
- Paß, T.; Aßfalg, M.; Tolve, M.; Blaess, S.; Rothermel, M.; Wiesner, R.J.; Ricke, K.M. The impact of mitochondrial dysfunction on dopa-minergic neurons in the olfactory bulb and odor detection. Mol. Neurobiol. 2020, 57, 3646–3657. [Google Scholar] [CrossRef]
- Blennow, K.; Hampel, H.; Weiner, M.; Zetterberg, H. Cerebrospinal fluid and plasma biomarkers in Alzheimer disease. Nat. Rev. Neurol. 2010, 6, 131–144. [Google Scholar] [CrossRef]
- Rochín-Hernández, L.J.; Jiménez-Acosta, M.A.; Ramírez-Reyes, L.; Figueroa-Corona, M.d.P.; Sánchez-González, V.J.; Orozco-Barajas, M.; Meraz-Ríos, M.A. The Proteome Profile of Olfactory Ecto-Mesenchymal Stem Cells-Derived from Patients with Familial Alzheimer’s Disease Reveals New Insights for AD Study. Int. J. Mol. Sci. 2023, 24, 12606. [Google Scholar] [CrossRef]
- Mastrangelo, A.; Bonato, M.; Cinque, P. Smell and taste disorders in COVID-19: From pathogenesis to clinical features and outcomes. Neurosci. Lett. 2021, 748, 135694. [Google Scholar] [CrossRef] [PubMed]
- Riccardi, G.; Niccolini, G.F.; Bellizzi, M.G.; Fiore, M.; Minni, A.; Barbato, C. Post-COVID-19 Anosmia and Therapies: Stay Tuned for New Drugs to Sniff Out. Diseases 2023, 11, 79. [Google Scholar] [CrossRef]
- von Bartheld, C.S.; Wang, L. Prevalence of Olfactory Dysfunction with the Omicron Variant of SARS-CoV-2: A Systematic Review and Meta-Analysis. Cells 2023, 12, 430. [Google Scholar] [CrossRef]
- Machado-Curbelo, C.; Gutierrez-Gil, J.; González-Quevedo, A. A Shift in SARS-CoV-2 Omicron Variant’s Entry Pathway Might Explain Different Clinical Outcomes. Medicc Rev. 2022, 24, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Grozdanić, M.; Vidmar, R.; Vizovišek, M.; Fonović, M. Degradomics in Biomarker Discovery. Proteom. Clin. Appl. 2019, 13, e1800138. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, I.; Tripathi, P.; Biswas, S. Mass spectrometry based protein biomarkers and drug target discovery and clinical diagnosis in Age-Related progressing neurodegenerative diseases. Drug Metab. Rev. 2022, 54, 22–36. [Google Scholar] [CrossRef]
- Lachen-Montes, M.; González-Morales, A.; Zelaya, M.V.; Pérez-Valderrama, E.; Ausín, K.; Ferrer, I.; Fernández-Irigoyen, J.; Santamaría, E. Olfactory bulb neuroproteomics reveals a chronological perturbation of survival routes and a disruption of prohibitin complex during Alzheimer’s disease progression. Sci. Rep. 2017, 7, 9115. [Google Scholar] [CrossRef]
- Dammalli, M.; Dey, G.; Kumar, M.; Madugundu, A.K.; Gopalakrishnan, L.; Gowrishankar, B.S.; Mahadevan, A.; Shankar, S.K.; Prasad, T.S.K. Proteomics of the Human Olfactory Tract. OMICS A J. Integr. Biol. 2018, 22, 77–87. [Google Scholar] [CrossRef]
- Keydar, I.; Ben-Asher, E.; Feldmesser, E.; Nativ, N.; Oshimoto, A.; Restrepo, D.; Matsunami, H.; Chien, M.-S.; Pinto, J.M.; Gilad, Y.; et al. General Olfactory Sensitivity Database (GOSdb): Candidate Genes and their Genomic Variations. Hum. Mutat. 2013, 34, 32–41. [Google Scholar] [CrossRef]
- Dammalli, M.; Dey, G.; Madugundu, A.K.; Kumar, M.; Rodrigues, B.; Gowda, H.; Siddaiah, B.G.; Mahadevan, A.; Shankar, S.K.; Prasad, T.S.K. Proteomic Analysis of the Human Olfactory Bulb. OMICS A J. Integr. Biol. 2017, 21, 440–453. [Google Scholar] [CrossRef]
- Zeng, H.-L.; Rao, X.; Zhang, L.-K.; Zhao, X.; Zhang, W.-P.; Wang, J.; Xu, F.; Guo, L. Quantitative proteomics reveals olfactory input-dependent alterations in the mouse olfactory bulb proteome. J. Proteom. 2014, 109, 125–142. [Google Scholar] [CrossRef] [PubMed]
- Lachén-Montes, M.; Mendizuri, N.; Ausín, K.; Pérez-Mediavilla, A.; Azkargorta, M.; Iloro, I.; Elortza, F.; Kondo, H.; Ohigashi, I.; Ferrer, I.; et al. Smelling the Dark Proteome: Functional Characterization of PITH Domain-Containing Protein 1 (C1orf128) in Olfactory Metabolism. J. Proteome Res. 2020, 19, 4826–4843. [Google Scholar] [CrossRef]
- Simões, T.; Charro, N.; Blonder, J.; Faria, D.; Couto, F.M.; Chan, K.C.; Waybright, T.; Isaaq, H.J.; Veenstra, T.D.; Penque, D. Molecular profiling of the human nasal epithelium: A proteomics approach. J. Proteom. 2011, 75, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.T.; A Coulombe, P.; Reed, R.R. Contribution of olfactory neural stem cells to tissue maintenance and regeneration. Nat. Neurosci. 2007, 10, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Virgilio, L.; Luarte, A.; Ponce, D.P.; Bruna, B.A.; Behrens, M.I. Analyzing Olfactory Neuron Precursors Non-Invasively Isolated through NADH FLIM as a Potential Tool to Study Oxidative Stress in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 6311. [Google Scholar] [CrossRef] [PubMed]
- Macron, C.; Lane, L.; Galindo, A.N.; Dayon, L. Deep Dive on the Proteome of Human Cerebrospinal Fluid: A Valuable Data Resource for Biomarker Discovery and Missing Protein Identification. J. Proteome Res. 2018, 17, 4113–4126. [Google Scholar] [CrossRef] [PubMed]
- Lachén-Montes, M.; Ausín, K.; Cartas-Cejudo, P.; Fernández-Irigoyen, J.; Santamaría, E. Combination of Antibody Arrays to Functionally Characterize Dark Proteins in Human Olfactory Neuroepithelial Cells. Methods Mol. Biol. 2021, 2344, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Taga, T.; Fukuda, S. Role of IL-6 in the Neural Stem Cell Differentiation. Clin. Rev. Allergy Immunol. 2005, 28, 249–256. [Google Scholar] [CrossRef]
- Corps, K.N.; Islam, Z.; Pestka, J.J.; Harkema, J.R. Neurotoxic, Inflammatory, and Mucosecretory Responses in the Nasal Airways of Mice Repeatedly Exposed to the Macrocyclic Trichothecene Mycotoxin Roridin A. Toxicol. Pathol. 2010, 38, 429–451. [Google Scholar] [CrossRef]
- Lachén-Montes, M.; Corrales, F.J.; Fernández-Irigoyen, J.; Santamaría, E. Proteomics Insights Into the Molecular Basis of SARS-CoV-2 Infection: What We Can Learn From the Human Olfactory Axis. Front. Microbiol. 2020, 11, 569052. [Google Scholar] [CrossRef]
- Zhou, J.; Guan, M.-Y.; Li, R.-T.; Qi, Y.-N.; Yang, G.; Deng, Y.-Q.; Li, X.-F.; Li, L.; Yang, X.; Liu, J.-F.; et al. Zika virus leads to olfactory disorders in mice by targeting olfactory ensheathing cells. EBioMedicine 2023, 89, 104457. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Zhou, J.; He, Q.; Li, R.-T.; Yang, G.; Zhang, Y.; Wu, S.-J.; Chen, Q.; Shi, J.-H.; Zhang, R.-R.; et al. SARS-CoV-2 infection in the mouse olfactory system. Cell Discov. 2021, 7, 49. [Google Scholar] [CrossRef]
- Lachén-Montes, M.; Mendizuri, N.; Ausín, K.; Echaide, M.; Blanco, E.; Chocarro, L.; de Toro, M.; Escors, D.; Fernández-Irigoyen, J.; Kochan, G.; et al. Metabolic dyshomeostasis induced by SARS-CoV-2 structural proteins reveals immunological insights into viral olfactory interactions. Front. Immunol. 2022, 13, 866564. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Wang, S.; Zhang, W.; Li, C.; Guo, L.; Wang, Z.; Li, H.; Zhang, H.; Zhou, Y.; Liang, W.; et al. Probing long COVID through a proteomic lens: A comprehensive two-year longitudinal cohort study of hospitalised survivors. EBioMedicine 2023, 98, 104851. [Google Scholar] [CrossRef]
- Li, H.; McLaurin, K.A.; Mactutus, C.F.; Rappaport, J.; Datta, P.K.; Booze, R.M. SARS-CoV-2 RNA persists in the central nervous system of non-human primates despite clinical recovery. Mol. Biomed. 2023, 4, 39. [Google Scholar] [CrossRef] [PubMed]
- Biswas, D.; Shenoy, S.V.; Chetanya, C.; Lachén-Montes, M.; Barpanda, A.; Athithyan, A.P.; Ghosh, S.; Ausín, K.; Zelaya, M.V.; Fernández-Irigoyen, J.; et al. Deciphering the Interregional and Interhemisphere Proteome of the Human Brain in the Context of the Human Proteome Project. J. Proteome Res. 2021, 20, 5280–5293. [Google Scholar] [CrossRef] [PubMed]
- Astillero-Lopez, V.; Gonzalez-Rodriguez, M.; Villar-Conde, S.; Flores-Cuadrado, A.; Martinez-Marcos, A.; Ubeda-Banon, I.; Saiz-Sanchez, D. Neurodegeneration and astrogliosis in the entorhinal cortex in Alzheimer’s disease: Stereological layer-specific assessment and proteomic analysis. Alzheimer’s Dement. 2022, 18, 2468–2480. [Google Scholar] [CrossRef]
- Salimi, M.; Tabasi, F.; Nazari, M.; Ghazvineh, S.; Salimi, A.; Jamaati, H.; Raoufy, M.R. The olfactory bulb modulates entorhinal cortex oscillations during spatial working memory. J. Physiol. Sci. 2021, 71, 21. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, X.; Chen, Y.; Qiu, W.; Ge, W.; Ma, C. Proteomic and Transcriptomic Analyses Reveal Pathological Changes in the Entorhinal Cortex Region that Correlate Well with Dysregulation of Ion Transport in Patients with Alzheimer’s Disease. Mol. Neurobiol. 2021, 58, 4007–4027. [Google Scholar] [CrossRef]
- Rajkumar, R.; Dawe, G.S. OBscure but not OBsolete: Perturbations of the frontal cortex in common between rodent olfactory bulbectomy model and major depression. J. Chem. Neuroanat. 2018, 91, 63–100. [Google Scholar] [CrossRef]
- Cartas-Cejudo, P.; Lachén-Montes, M.; Fernández-Irigoyen, J.; Santamaría, E. Tackling the Biological Meaning of the Human Olfactory Bulb Dyshomeostatic Proteome across Neurological Disorders: An Integrative Bioinformatic Approach. Int. J. Mol. Sci. 2021, 22, 11340. [Google Scholar] [CrossRef] [PubMed]
- Romano, R.; Bucci, C. Role of EGFR in the Nervous System. Cells 2020, 9, 1887. [Google Scholar] [CrossRef] [PubMed]
- Cartas-Cejudo, P.; Lachén-Montes, M.; Ferrer, I.; Fernández-Irigoyen, J.; Santamaría, E. Sex-divergent effects on the NAD+-dependent deacetylase sirtuin signaling across the olfactory–entorhinal–amygdaloid axis in Alzheimer’s and Parkinson’s diseases. Biol. Sex Differ. 2023, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Bubak, A.N.; Merle, L.; Niemeyer, C.S.; Baxter, B.D.; Polese, A.G.; Ramakrishnan, V.; Gomez, J.; Madrigal, L.; Villegas-Lanau, A.; Lopera, F.; et al. Signatures for viral infection and inflammation in the proximal olfactory system in familial Alzheimer’s disease. Neurobiol. Aging 2023, 123, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Zelaya, M.V.; Pérez-Valderrama, E.; de Morentin, X.M.; Tuñon, T.; Ferrer, I.; Luquin, M.R.; Fernandez-Irigoyen, J.; Santamaría, E. Olfactory bulb proteome dynamics during the progression of sporadic Alzheimer’s disease: Identification of common and distinct olfactory targets across Alzheimer-related co-pathologies. Oncotarget 2015, 6, 39437–39456. [Google Scholar] [CrossRef] [PubMed]
- Lachen-Montes, M.; Zelaya, M.V.; Segura, V.; Fernández-Irigoyen, J.; Santamaría, E. Progressive modulation of the human olfactory bulb transcriptome during Alzheimer’s disease evolution: Novel insights into the olfactory signaling across proteinopathies. Oncotarget 2017, 8, 69663–69679. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.-J.; Son, G.; Bae, J.; Kim, S.Y.; Yoo, Y.K.; Park, D.; Baek, S.Y.; Chang, K.-A.; Suh, Y.-H.; Lee, Y.-B.; et al. Longitudinal profiling of oligomeric Aβ in human nasal discharge reflecting cognitive decline in probable Alzheimer’s disease. Sci. Rep. 2020, 10, 11234. [Google Scholar] [CrossRef] [PubMed]
- Lachén-Montes, M.; González-Morales, A.; Schvartz, D.; Zelaya, M.V.; Ausin, K.; Fernández-Irigoyen, J.; Sánchez, J.C.; Santamaría, E. The olfactory bulb proteotype differs across frontotemporal dementia spectrum. J. Proteom. 2019, 201, 37–47. [Google Scholar] [CrossRef]
- Martin-Lopez, E.; Vidyadhara, D.J.; Liberia, T.; Meller, S.J.; Harmon, L.E.; Hsu, R.M.; Spence, N.; Brennan, B.; Han, K.; Yücel, B.; et al. α-Synuclein Pathology and Reduced Neurogenesis in the Olfactory System Affect Olfaction in a Mouse Model of Parkinson’s Disease. J. Neurosci. 2023, 43, 1051–1071. [Google Scholar] [CrossRef]
- Lachén-Montes, M.; González-Morales, A.; Iloro, I.; Elortza, F.; Ferrer, I.; Gveric, D.; Fernández-Irigoyen, J.; Santamaría, E. Unveiling the olfactory proteostatic disarrangement in Parkinson’s disease by proteome-wide profiling. Neurobiol. Aging 2019, 73, 123–134. [Google Scholar] [CrossRef]
- Lachén-Montes, M.; Mendizuri, N.; Schvartz, D.; Fernández-Irigoyen, J.; Sánchez, J.C.; Santamaría, E. Proteomic Characterization of the Olfactory Molecular Imbalance in Dementia with Lewy Bodies. Int. J. Mol. Sci. 2020, 21, 6371. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, L.; Altera, A.; Barone, V.; Bonente, D.; Bacci, T.; De Benedetto, E.; Bini, L.; Tosi, G.M.; Galvagni, F.; Bertelli, E. Untangling the Extracellular Matrix of Idiopathic Epiretinal Membrane: A Path Winding among Structure, Interactomics and Translational Medicine. Cells 2022, 11, 2531. [Google Scholar] [CrossRef] [PubMed]
- Lachén-Montes, M.; Íñigo-Marco, I.; Cartas-Cejudo, P.; Fernández-Irigoyen, J.; Santamaría, E. Olfactory Bulb Proteomics Reveals Widespread Proteostatic Disturbances in Mixed Dementia and Guides for Potential Serum Biomarkers to Discriminate Alzheimer Disease and Mixed Dementia Phenotypes. J. Pers. Med. 2021, 11, 503. [Google Scholar] [CrossRef] [PubMed]
- Lachén-Montes, M.; Mendizuri, N.; Ausin, K.; Andrés-Benito, P.; Ferrer, I.; Fernández-Irigoyen, J.; Santamaría, E. Amyotrophic Lateral Sclerosis Is Accompanied by Protein Derangements in the Olfactory Bulb-Tract Axis. Int. J. Mol. Sci. 2020, 21, 8311. [Google Scholar] [CrossRef] [PubMed]
- Cartas-Cejudo, P.; Cortes, A.; Lachén-Montes, M.; Cubero, E.A.; Peral, E.; Ausin, K.; Diaz-Peña, R.; Fernández-Irigoyen, J.; Santamaria, E. Mapping the human brain proteome: Opportunities, challenges and clinical potential. Expert Rev. Proteom. 2024; accepted. [Google Scholar] [CrossRef] [PubMed]
- Basak, I.; Harfoot, R.; Palmer, J.E.; Kumar, A.; Quiñones-Mateu, M.E.; Schweitzer, L.; Hughes, S.M. Neuroproteomic Analysis after SARS-CoV-2 Infection Reveals Overrepresented Neurodegeneration Pathways and Disrupted Metabolic Pathways. Biomolecules 2023, 13, 1597. [Google Scholar] [CrossRef] [PubMed]
- Adilbay, D.; Gonzales, J.; Zazhytska, M.; Demetrio de Souza Franca, P.; Roberts, S.; Viray, T.; Artschwager, R.; Patel, S.; Kodra, A.; Overdevest, J.B.; et al. Non-invasive diagnostic method to objectively measure olfaction and diagnose smell disorders by molecularly targeted fluorescent imaging agent. bioRxiv 2022. [Google Scholar] [CrossRef]
- Iravani, B.; Arshamian, A.; Schaefer, M.; Svenningsson, P.; Lundström, J.N. A non-invasive olfactory bulb measure dissociates Parkinson’s patients from healthy controls and discloses disease duration. NPJ Park. Dis. 2021, 7, 75. [Google Scholar] [CrossRef]
- Duan, S.; Yang, J.; Cui, Z.; Li, J.; Zheng, H.; Zhao, T.; Yuan, Y.; Liu, Y.; Zhao, L.; Wang, Y.; et al. Seed amplification assay of nasal swab extracts for accurate and non-invasive molecular diagnosis of neurodegenerative diseases. Transl. Neurodegener. 2023, 12, 13. [Google Scholar] [CrossRef]
- Lachén-Montes, M.; Fernández-Irigoyen, J.; Santamaría, E. Deconstructing the molecular architecture of olfactory areas using proteomics. Proteom. Clin. Appl. 2016, 10, 1178–1190. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Neurodegenerative Diseases | Single Proteins/Pathway Alterations |
|---|---|
| Alzheimer disease | PNP, NCAM2, LY6H, COL6A3, PRDX6, EGFR, SREBF1, NFMUE1, STAT3, p38, MAPK, PDK1, PKC, Phb2 |
| Fronto-Temporal Dementia | MAPK, CAMKII, PKC |
| PSP | SEK1/MKK4, SAPK |
| FTLD-TDP43 | PDK1, NMYC, Phb1 |
| Lewy-type α-synucleinopathies | |
| PD | PNP, NCAM2, LY6H, COL6A3, PRDX6, ERK1/2, MKK3/6, PDK1/PKC, HDAC1, CIITA, MYC, CDC5L, USF, PTF1β |
| LBD | AKT/MAPK/SEK1/p38 MAPK |
| Mixed dementia | PNP, NCAM2, LY6H, COL6A3, PRDX6, EGFR, Phb2, LRP-1, NFIC, ARF3, NOTCH3 |
| Amyotrophic lateral sclerosis | PNP, NCAM2, LY6H, COL6A3, PRDX6, OMP, GFAP, Bcl-xL, IL-18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusi, E.; Pennacchia, F.; Ruqa, W.A.; Talarico, G.; Bruno, G.; Minni, A.; Barbato, C. Proteoform Analysis of the Human Olfactory System: A Window into Neurodegenerative Diseases. Proteomes 2024, 12, 9. https://doi.org/10.3390/proteomes12010009
Rusi E, Pennacchia F, Ruqa WA, Talarico G, Bruno G, Minni A, Barbato C. Proteoform Analysis of the Human Olfactory System: A Window into Neurodegenerative Diseases. Proteomes. 2024; 12(1):9. https://doi.org/10.3390/proteomes12010009
Chicago/Turabian StyleRusi, Eqrem, Fiorenza Pennacchia, Wael Abu Ruqa, Giuseppina Talarico, Giuseppe Bruno, Antonio Minni, and Christian Barbato. 2024. "Proteoform Analysis of the Human Olfactory System: A Window into Neurodegenerative Diseases" Proteomes 12, no. 1: 9. https://doi.org/10.3390/proteomes12010009
APA StyleRusi, E., Pennacchia, F., Ruqa, W. A., Talarico, G., Bruno, G., Minni, A., & Barbato, C. (2024). Proteoform Analysis of the Human Olfactory System: A Window into Neurodegenerative Diseases. Proteomes, 12(1), 9. https://doi.org/10.3390/proteomes12010009