
Asymmetric Growth of Tumor Spheroids in a Symmetric Environment

 ,
,  ,
,  and
and 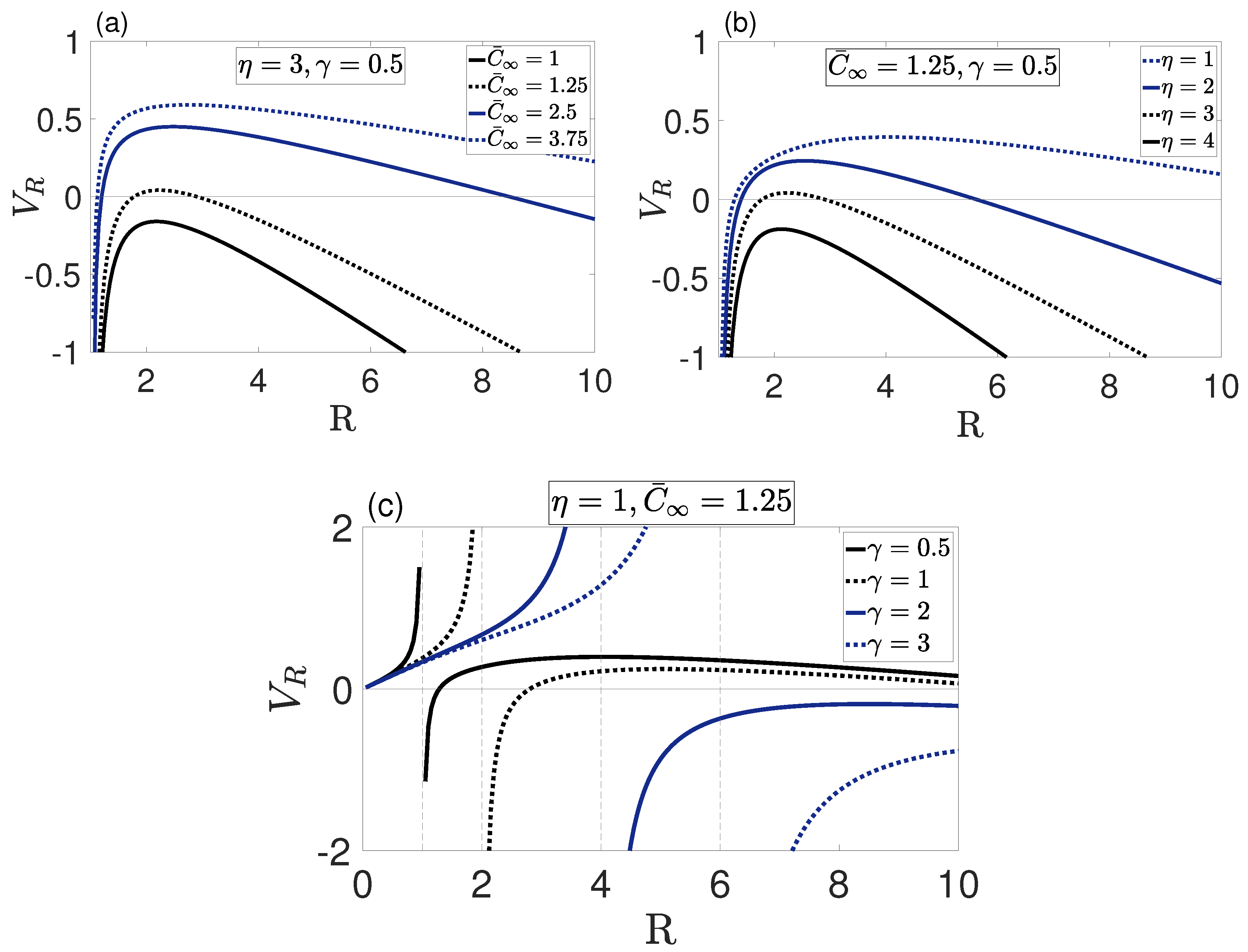{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Model Formulation
2.1. Analytical Solutions
Symmetric Steady-State Solutions
2.2. Instability to Asymmetric Perturbation
2.3. Growing Asymmetric Modes
3. Asymmetric Configuration
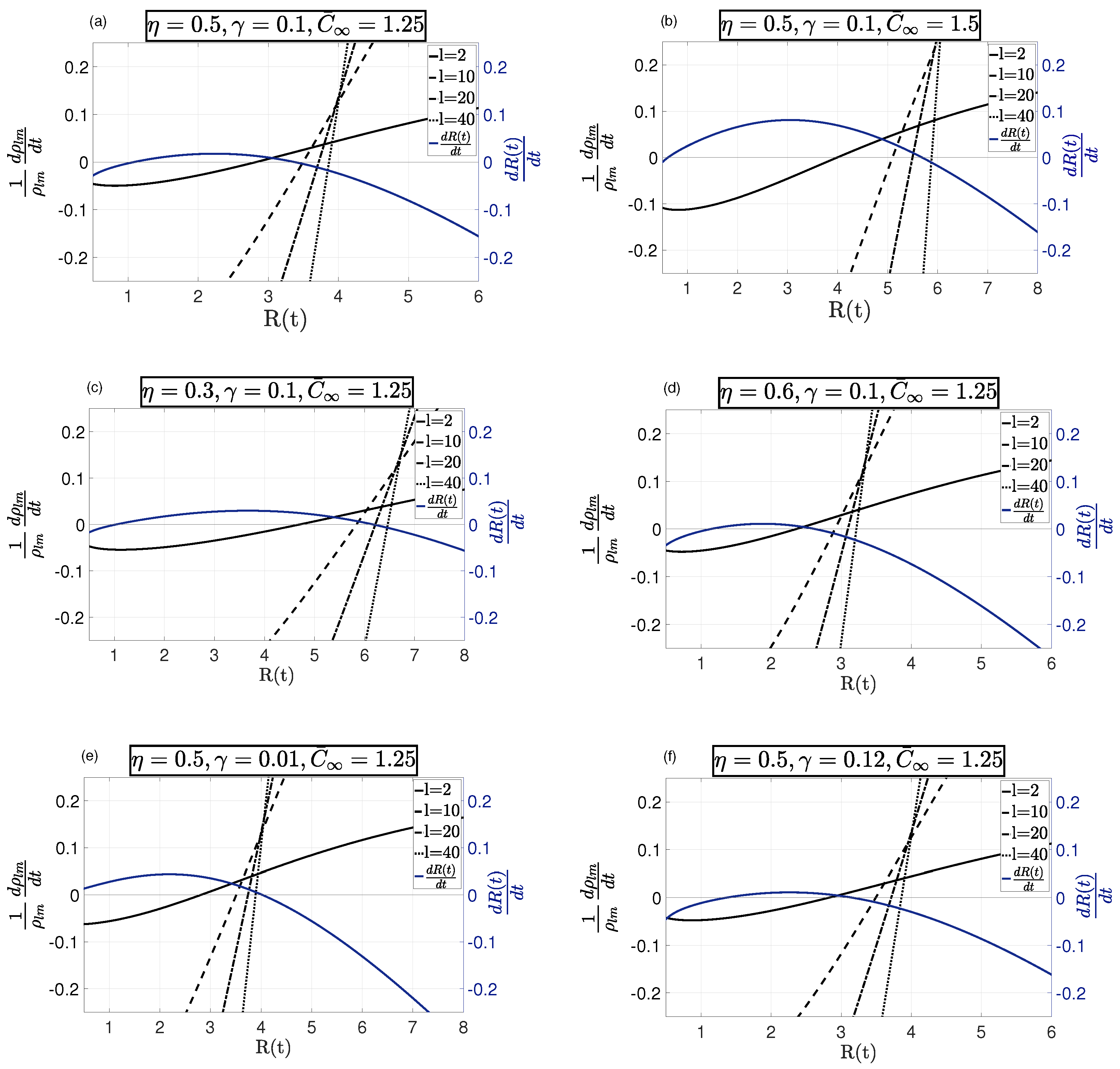
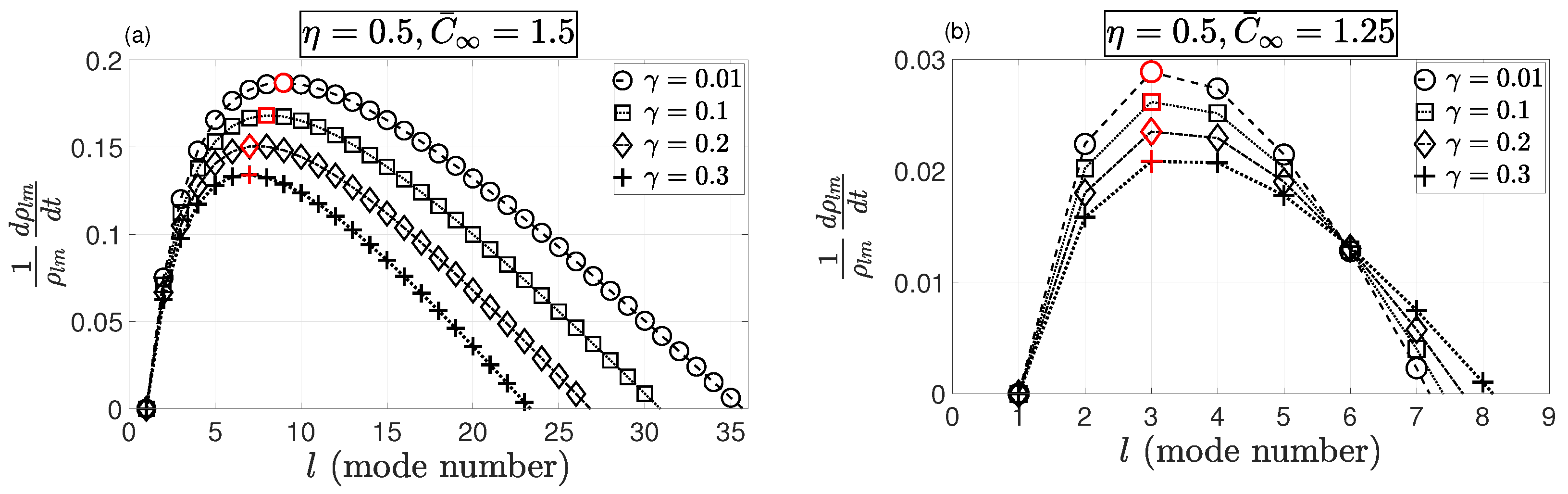
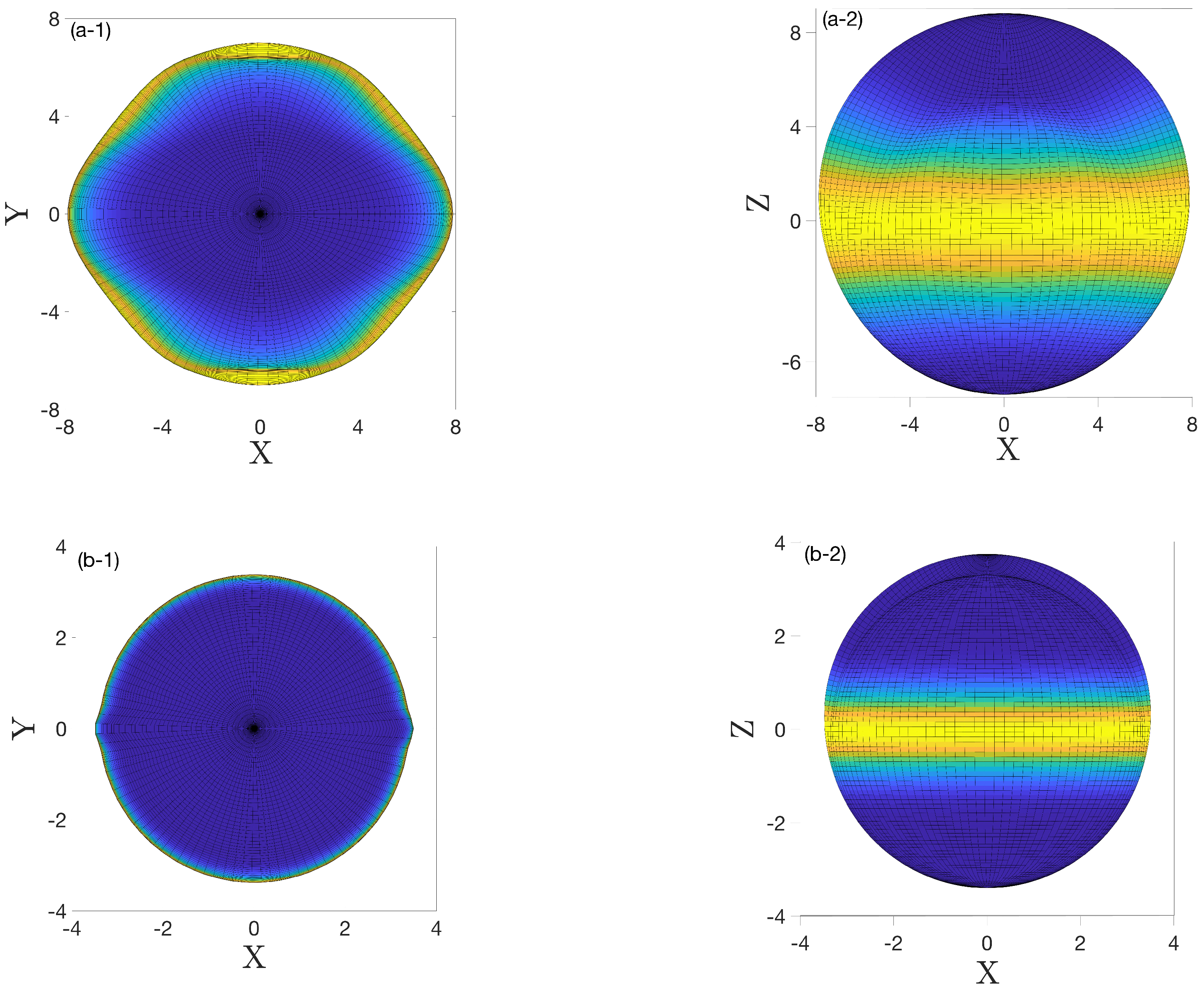
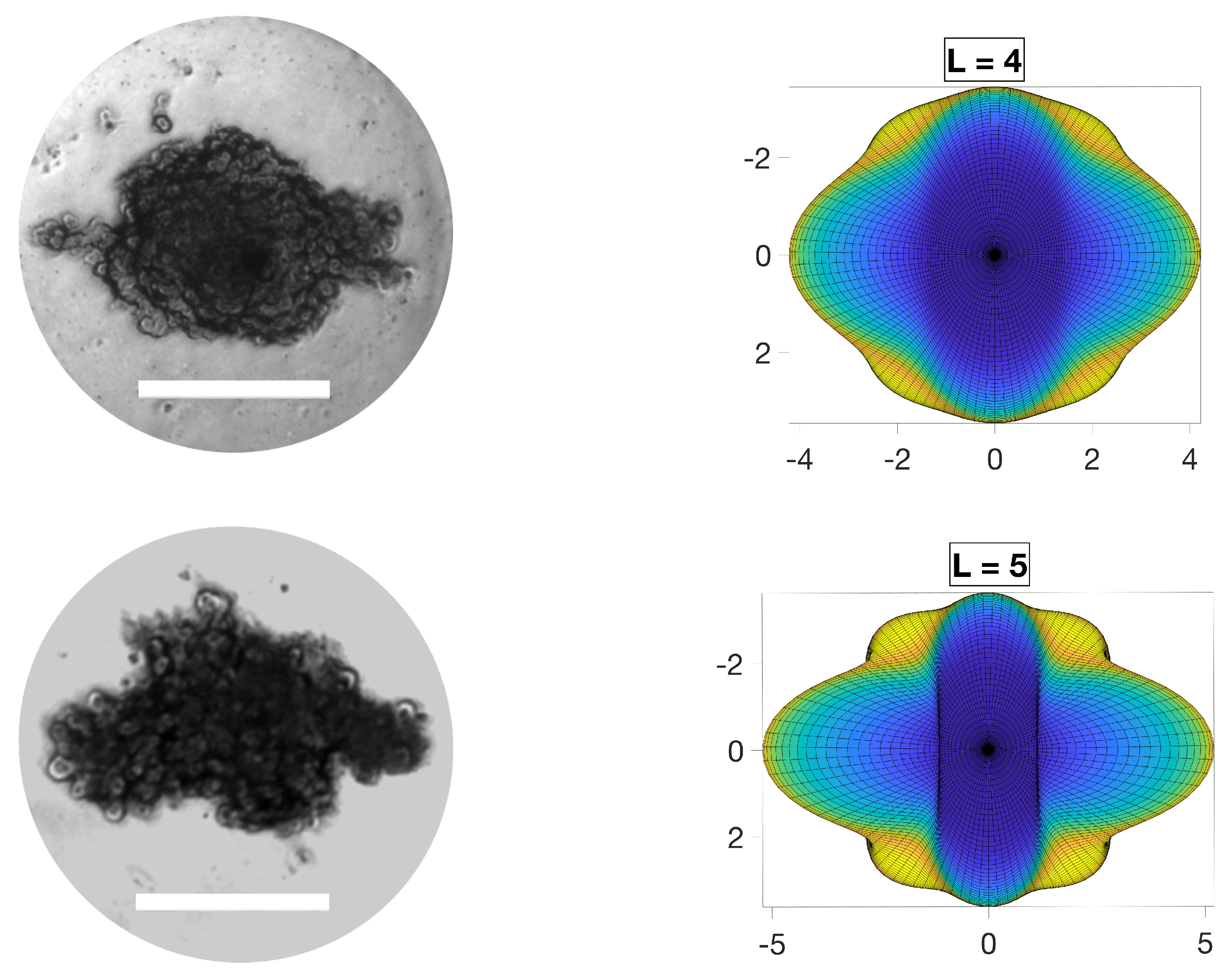
4. Results and Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Please, C.; Pettet, G.; McElwain, D. A new approach to modelling the formation of necrotic regions in tumours. Appl. Math. Lett. 1998, 11, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Altrock, P.M.; Liu, L.L.; Michor, F. The mathematics of cancer: Integrating quantitative models. Nat. Rev. Cancer 2015, 15, 730–745. [Google Scholar] [CrossRef]
- Araujo, R.P.; McElwain, D.L.S. A history of the study of solid tumour growth: The contribution of mathematical modelling. Bull. Math. Biol. 2004, 66, 1039. [Google Scholar] [CrossRef]
- Landman, K.A.; Please, C.P. Tumour dynamics and necrosis: Surface tension and stability. Math. Med. Biol. J. IMA 2001, 18, 131–158. [Google Scholar] [CrossRef]
- Ward, J.P.; King, J.R. Mathematical modelling of avascular-tumour growth. Math. Med. Biol. J. IMA 1997, 14, 39–69. [Google Scholar] [CrossRef]
- Cristini, V.; Lowengrub, J. Multiscale Modeling of Cancer: An Integrated Experimental and Mathematical Modeling Approach; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Chaplain, M.; Britton, N. On the concentration profile of a growth inhibitory factor in multicell spheroids. Math. Biosci. 1993, 115, 233–243. [Google Scholar] [CrossRef]
- Byrne, H.M.; Chaplain, M. Growth of necrotic tumors in the presence and absence of inhibitors. Math. Biosci. 1996, 135, 187–216. [Google Scholar] [CrossRef]
- Adam, J.A. A mathematical model of tumor growth. III. Comparison with experiment. Math. Biosci. 1987, 86, 213–227. [Google Scholar] [CrossRef]
- Casciari, J.; Sotirchos, S.; Sutherland, R. Mathematical modelling of microenvironment and growth in EMT6/Ro multicellular tumour spheroids. Cell Prolif. 1992, 25, 1–22. [Google Scholar] [CrossRef]
- Casciari, J.J.; Sotirchos, S.V.; Sutherland, R.M. Variations in tumor cell growth rates and metabolism with oxygen concentration, glucose concentration, and extracellular pH. J. Cell. Physiol. 1992, 151, 386–394. [Google Scholar] [CrossRef]
- Jiang, Y.; Pjesivac-Grbovic, J.; Cantrell, C.; Freyer, J.P. A multiscale model for avascular tumor growth. Biophys. J. 2005, 89, 3884–3894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amereh, M.; Edwards, R.; Akbari, M.; Nadler, B. In-Silico Modeling of Tumor Spheroid Formation and Growth. Micromachines 2021, 12, 749. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Bresch, D.; Colin, T.; Grenier, E.; Ribba, B.; Saut, O. Computational modeling of solid tumor growth: The avascular stage. SIAM J. Sci. Comput. 2010, 32, 2321–2344. [Google Scholar] [CrossRef]
- Yoo, Y.D.; Kwon, Y.T. Molecular mechanisms controlling asymmetric and symmetric self-renewal of cancer stem cells. J. Anal. Sci. Technol. 2015, 6, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benzekry, S.; Lamont, C.; Barbolosi, D.; Hlatky, L.; Hahnfeldt, P. Mathematical modeling of tumor–tumor distant interactions supports a systemic control of tumor growth. Cancer Res. 2017, 77, 5183–5193. [Google Scholar] [CrossRef] [Green Version]
- Mascheroni, P.; Carfagna, M.; Grillo, A.; Boso, D.; Schrefler, B.A. An avascular tumor growth model based on porous media mechanics and evolving natural states. Math. Mech. Solids 2018, 23, 686–712. [Google Scholar] [CrossRef]
- Byrne, H.; Chaplain, M. Modelling the role of cell-cell adhesion in the growth and development of carcinomas. Math. Comput. Model. 1996, 24, 1–17. [Google Scholar] [CrossRef]
- Li, C.K.N. The glucose distribution in 9l rat brain multicell tumor spheroids and its effect on cell necrosis. Cancer 1982, 50, 2066–2073. [Google Scholar] [CrossRef]
- Cui, S. Analysis of a mathematical model for the growth of tumors under the action of external inhibitors. J. Math. Biol. 2002, 44, 395–426. [Google Scholar] [CrossRef]
- Cui, S.; Friedman, A. Analysis of a mathematical model of the effect of inhibitors on the growth of tumors. Math. Biosci. 2000, 164, 103–137. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Cristini, V.; Nie, Q.; Lowengrub, J.S. Nonlinear three-dimensional simulation of solid tumor growth. Discret. Contin. Dyn. Syst. B 2007, 7, 581. [Google Scholar] [CrossRef]
- Cristini, V.; Lowengrub, J.; Nie, Q. Nonlinear simulation of tumor growth. J. Math. Biol. 2003, 46, 191–224. [Google Scholar] [CrossRef]
- Wu, J.; Cui, S. Asymptotic behaviour of solutions of a free boundary problem modelling the growth of tumours in the presence of inhibitors. Nonlinearity 2007, 20, 2389. [Google Scholar] [CrossRef]
- Langer, J.S. Instabilities and pattern formation in crystal growth. Rev. Mod. Phys. 1980, 52, 1. [Google Scholar] [CrossRef]
- Byrd, B.K.; Krishnaswamy, V.; Gui, J.; Rooney, T.; Zuurbier, R.; Rosenkranz, K.; Paulsen, K.; Barth, R.J. The shape of breast cancer. Breast Cancer Res. Treat. 2020, 183, 403–410. [Google Scholar] [CrossRef]
- Seyfoori, A.; Samiei, E.; Jalili, N.; Godau, B.; Rahmanian, M.; Farahmand, L.; Majidzadeh-A, K.; Akbari, M. Self-filling microwell arrays (SFMAs) for tumor spheroid formation. Lab Chip 2018, 18, 3516–3528. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amereh, M.; Bahri, Y.; Edwards, R.; Akbari, M.; Nadler, B. Asymmetric Growth of Tumor Spheroids in a Symmetric Environment. Mathematics 2022, 10, 1955. https://doi.org/10.3390/math10121955
Amereh M, Bahri Y, Edwards R, Akbari M, Nadler B. Asymmetric Growth of Tumor Spheroids in a Symmetric Environment. Mathematics. 2022; 10(12):1955. https://doi.org/10.3390/math10121955
Chicago/Turabian StyleAmereh, Meitham, Yakine Bahri, Roderick Edwards, Mohsen Akbari, and Ben Nadler. 2022. "Asymmetric Growth of Tumor Spheroids in a Symmetric Environment" Mathematics 10, no. 12: 1955. https://doi.org/10.3390/math10121955
APA StyleAmereh, M., Bahri, Y., Edwards, R., Akbari, M., & Nadler, B. (2022). Asymmetric Growth of Tumor Spheroids in a Symmetric Environment. Mathematics, 10(12), 1955. https://doi.org/10.3390/math10121955