
Bilateral Feedback in Oscillator Model Is Required to Explain the Coupling Dynamics of Hes1 with the Cell Cycle


Abstract
:1. Introduction
2. Mathematical Modelling Strategy
2.1. Phase Representation of Bcc Data
2.2. Specifying the Coupled Oscillator Model
- will be considered the peak of an oscillatory Hes1 wave, and is the trough.
- will denote the beginning and the end of a cell cycle, i.e., a cell division event.
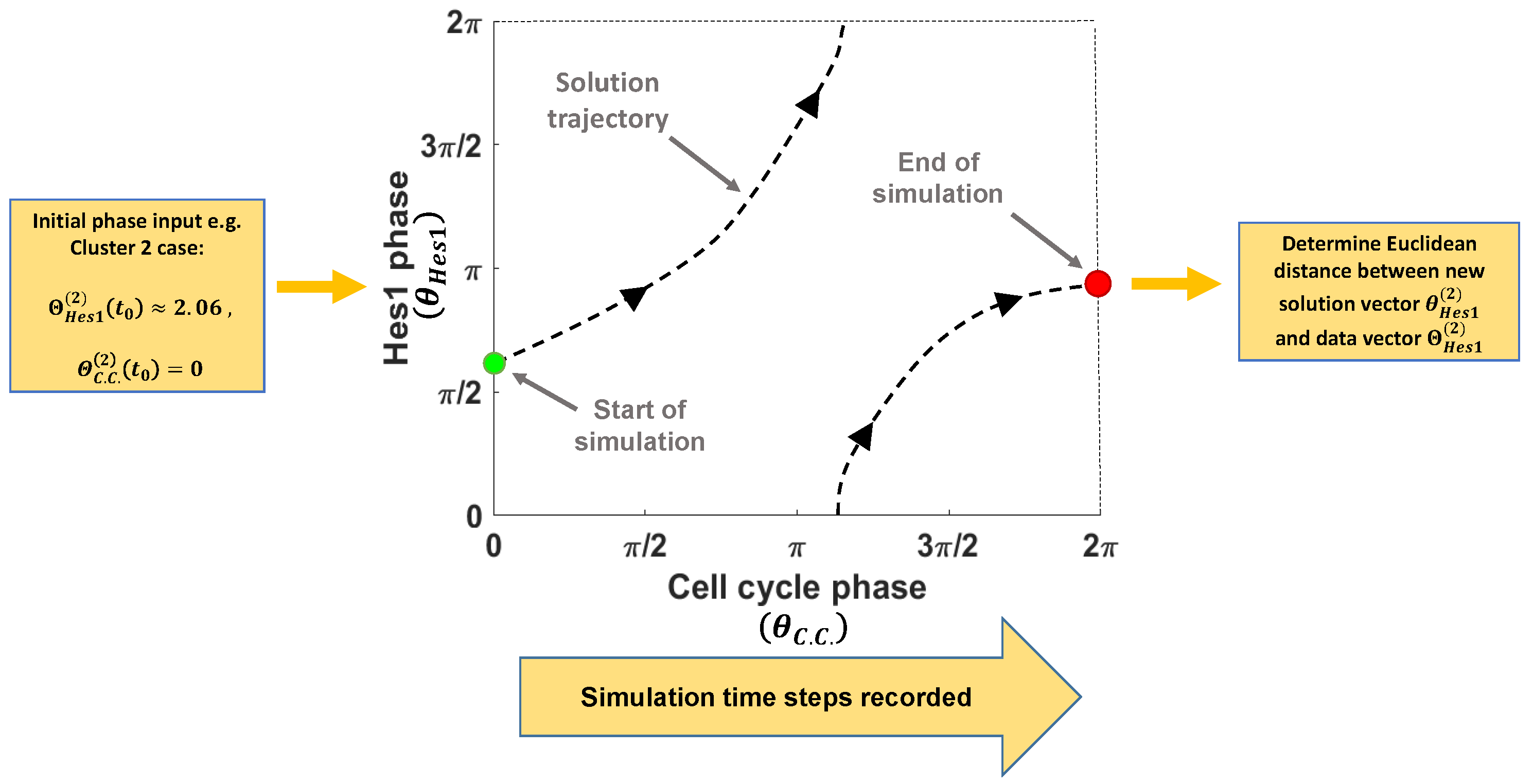
2.3. Numerical Implementation
2.4. Model Optimisation Strategy
3. Results of Model Simulations and Comparisons to Biological Data
3.1. The Uncoupled Scenario
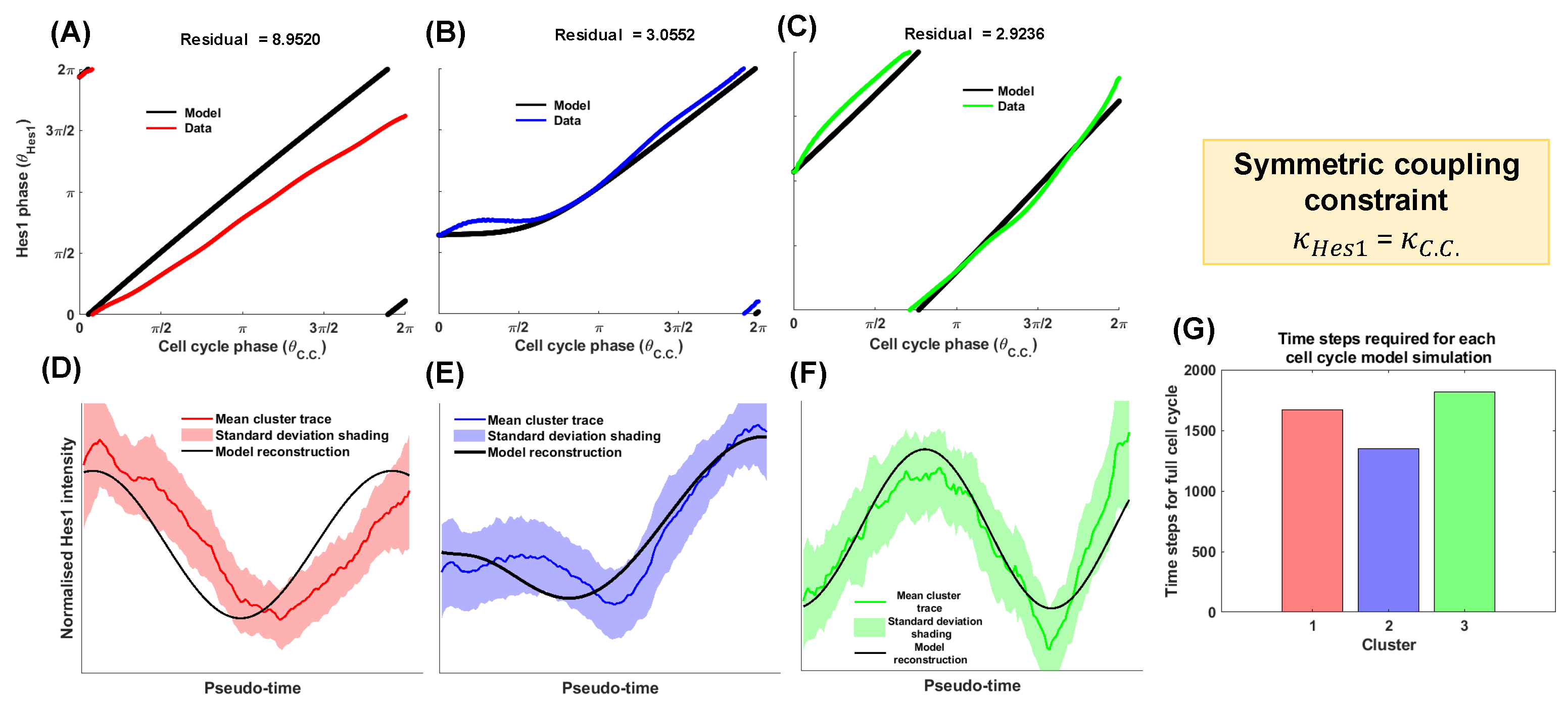
3.2. The Symmetric Interaction Strength Scenario
3.3. The Unconstrained Interaction Strength Scenario
3.4. Asymmetry in Interaction Strength Is Predictive of Elongation in Bcsc Data
3.5. Cluster-Dependent Coupling Strength Points to Gene Expression and Cell Cycle Duration Differences
3.6. Mathematical Analysis and Long-Term Behaviour of Our Model
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Unidirectional Interaction Constraints


References
- Purvis, J.E.; Karhohs, K.W.; Mock, C.; Batchelor, E.; Loewer, A.; Lahav, G. p53 dynamics control cell fate. Science 2012, 336, 1440–1444. [Google Scholar] [CrossRef] [Green Version]
- Imayoshi, I.; Isomura, A.; Harima, Y.; Kawaguchi, K.; Kori, H.; Miyachi, H.; Kageyama, R. Oscillatory control of factors determining multipotency and fate in mouse neural progenitors. Science 2013, 342, 1203–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, C.S.; Biga, V.; Boyd, J.; Kursawe, J.; Ymisson, B.; Spiller, D.G.; Papalopulu, N. Quantitative single-cell live imaging links HES5 dynamics with cell-state and fate in murine neurogenesis. Nat. Commun. 2019, 10, 2835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, D.; Hasty, J. Dynamics of single-cell gene expression. Mol. Syst. Biol. 2006, 2, 64. [Google Scholar] [CrossRef] [Green Version]
- Sonnen, K.F.; Janda, C.Y. Signalling dynamics in embryonic development. Biochem. J. 2021, 478, 4045–4070. [Google Scholar] [CrossRef]
- Purvis, J.E.; Lahav, G. Encoding and decoding cellular information through signaling dynamics. Cells 2013, 152, 945–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, E.; Sung, M.H. Challenges of Decoding Transcription Factor Dynamics in Terms of Gene Regulation. Cells 2018, 7, 132. [Google Scholar] [CrossRef] [Green Version]
- Behar, M.; Dohlman, H.G.; Elston, T.C. Kinetic insulation as an effective mechanism for achieving pathway specificity in intracellular signaling networks. Proc. Natl. Acad. Sci. USA 2007, 104, 16146–16151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, K.; Alberts, B.; Johnson, A.; Walter, P.; Hunt, T. Molecular Biology of the Cell, 6th ed.; Garland Science: New York, NY, USA, 2015; pp. 963–967. [Google Scholar]
- Tyson, J.J.; Csikasz-Nagy, A.; Novak, B. The dynamics of cell cycle regulation. Bioessays 2002, 24, 1095–1109. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.; MacLellan, W.R.; Weiss, J.N. Dynamics of the cell cycle: Checkpoints, sizers, and timers. Biophys. J. 2003, 85, 3600–3611. [Google Scholar] [CrossRef] [Green Version]
- Dalton, S. Linking the cell cycle to cell fate decisions. Trends Cell Biol. 2015, 25, 592–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauklin, S.; Vallier, L. The cell-cycle state of stem cells determines cell fate propensity. Cell 2013, 155, 135–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roccio, M.; Schmitter, D.; Knobloch, M.; Okawa, Y.; Sage, D.; Lutolf, M.P. Predicting stem cell fate changes by differential cell cycle progression patterns. Development 2013, 140, 459–470. [Google Scholar] [CrossRef] [Green Version]
- de Lichtenberg, K.H.; Seymour, P.A.; Jørgensen, M.C.; Kim, Y.H.; Grapin-Botton, A.; Magnuson, M.A.; Serup, P. Notch controls multiple pancreatic cell fate regulators through direct Hes1-mediated repression. BioRxiv 2019, 336305. [Google Scholar]
- Kageyama, R.; Ohtsuka, T.; Tomita, K. The bHLH gene Hes1 regulates differentiation of multiple cell types. Mol. Cells 2000, 10, 1–7. [Google Scholar] [CrossRef]
- Kageyama, R.; Ohtsuka, T.; Kobayashi, T. Roles of Hes genes in neural development. Dev. Growth Differ. 2008, 50, S97–S103. [Google Scholar] [CrossRef]
- Isomura, A.; Kageyama, R. Ultradian oscillations and pulses: Coordinating cellular responses and cell fate decisions. Development 2014, 141, 3627–3636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinopoulou, E.; Biga, V.; Sabherwal, N.; Miller, A.; Desai, J.; Adamson, A.D.; Papalopulu, N. HES1 protein oscillations are necessary for neural stem cells to exit from quiescence. Iscience 2021, 24, 103198. [Google Scholar] [CrossRef]
- Noda, N.; Honma, S.; Ohmiya, Y. Hes1 is required for contact inhibition of cell proliferation in 3T3-L1 preadipocytes. Genes Cells 2011, 16, 704–713. [Google Scholar] [CrossRef]
- Murata, K.; Hattori, M.; Hirai, N.; Shinozuka, Y.; Hirata, H.; Kageyama, R.; Minato, N. Hes1 directly controls cell proliferation through the transcriptional repression of p27Kip1. Mol. Cell. Biol. 2005, 25, 4262–4271. [Google Scholar] [CrossRef] [Green Version]
- Ochi, S.; Imaizumi, Y.; Shimojo, H.; Miyachi, H.; Kageyama, R. Oscillatory expression of Hes1 regulates cell proliferation and neuronal differentiation in the embryonic brain. Development 2020, 147, dev182204. [Google Scholar] [CrossRef]
- Cenciarelli, C.; Marei, H.E.; Zonfrillo, M.; Casalbore, P.; Felsani, A.; Giannetti, S.; Mangiola, A. The interference of Notch1 target Hes1 affects cell growth, differentiation and invasiveness of glioblastoma stem cells through modulation of multiple oncogenic targets. Oncotarget 2017, 8, 17873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sang, L.; Roberts, J.M.; Coller, H.A. Hijacking HES1: How tumors co-opt the anti-differentiation strategies of quiescent cells. Trends Mol. Med. 2010, 16, 17–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.H.; Dai, X.M.; Du, B. Hes1: A Key Role in Stemness, Metastasis and Multidrug Resistance. Cancer Biol. Ther. 2015, 16, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabherwal, N.; Rowntree, A.; Marinopoulou, E.; Pettini, T.; Hourihane, S.; Thomas, R.; Papalopulu, N. Differential phase register of Hes1 oscillations with mitoses underlies cell-cycle heterogeneity in ER+ breast cancer cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2113527118. [Google Scholar] [CrossRef] [PubMed]
- Comşa, Ş.; Cimpean, A.M.; Raica, M. The story of MCF-7 breast cancer cell line: 40 years of experience in research. Anticancer Res. 2015, 35, 3147–3154. [Google Scholar]
- Lee, A.V.; Oesterreich, S.; Davidson, N.E. MCF-7 cells—Changing the course of breast cancer research and care for 45 years. J. Natl. Cancer Inst. 2015, 107, djv073. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Cong, Y.; Wang, D.; Sun, Y.; Deng, L.; Liu, Y.; Wicha, M.S. Breast cancer stem cells transition between epithelial and mesenchymal states reflective of their normal counterparts. Stem Cell Rep. 2014, 2, 78–91. [Google Scholar] [CrossRef]
- Hirata, H.; Yoshiura, S.; Ohtsuka, T.; Bessho, Y.; Harada, T.; Yoshikawa, K.; Kageyama, R. Oscillatory expression of the bHLH factor Hes1 regulated by a negative feedback loop. Science 2002, 298, 840–843. [Google Scholar] [CrossRef] [Green Version]
- Monk, N.A. Oscillatory expression of Hes1, p53, and NF-kB driven by transcriptional time delays. Curr. Biol. 2003, 13, 1409–1413. [Google Scholar] [CrossRef] [Green Version]
- Momiji, H.; Monk, N.A. Dissecting the dynamics of the Hes1 genetic oscillator. J. Theor. Biol. 2008, 254, 784–798. [Google Scholar] [CrossRef]
- Ay, A.; Arnosti, D.N. Mathematical modeling of gene expression: A guide for the perplexed biologist. Crit. Rev. Biochem. Mol. Biol. 2011, 46, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Robert, P. Mathematical models of gene expression. Probab. Surv. 2019, 16, 277–332. [Google Scholar] [CrossRef]
- Smolen, P.; Baxter, D.A.; Byrne, J.H. Mathematical modeling of gene networks. Neuron 2000, 26, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, M.; Phillips, N.E.; Manning, C.; Galla, T.; Papalopulu, N. microRNA input into a neural ultradian oscillator controls emergence and timing of alternative cell states. Nat. Commun. 2014, 5, 3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, N.E.; Manning, C.S.; Pettini, T.; Biga, V.; Marinopoulou, E.; Stanley, P.; Papalopulu, N. Stochasticity in the miR-9/Hes1 oscillatory network can account for clonal heterogeneity in the timing of differentiation. eLife 2016, 5, e16118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrio, M.; Burrage, K.; Leier, A.; Tian, T. Oscillatory regulation of Hes1: Discrete stochastic delay modelling and simulation. PLoS Comput. Biol. 2006, 2, e117. [Google Scholar] [CrossRef] [Green Version]
- Pfeuty, B. Multistability and transitions between spatiotemporal patterns through versatile Notch-Hes signaling. J. Theor. Biol. 2022, 539, 111060. [Google Scholar] [CrossRef]
- Bertuzzi, A.; Gandolfi, A.; Giovenco, M.A. Mathematical models of the cell cycle with a view to tumor studies. Math. Biosci. 1981, 53, 159–188. [Google Scholar] [CrossRef]
- Basse, B.; Baguley, B.C.; Marshall, E.S.; Joseph, W.R.; van Brunt, B.; Wake, G.; Wall, D.J. A mathematical model for analysis of the cell cycle in cell lines derived from human tumors. J. Math. Biol. 2003, 47, 295–312. [Google Scholar] [CrossRef]
- Sible, J.C.; Tyson, J.J. Mathematical modeling as a tool for investigating cell cycle control networks. Methods 2007, 41, 238–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gérard, C.; Goldbeter, A. Temporal self-organization of the cyclin/Cdk network driving the mammalian cell cycle. Proc. Natl. Acad. Sci. USA 2009, 106, 21643–21648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weis, M.C.; Avva, J.; Jacobberger, J.W.; Sreenath, S.N. A data-driven, mathematical model of mammalian cell cycle regulation. PloS ONE 2014, 9, e97130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyson, J.J.; Novák, B. Models in biology: Lessons from modeling regulation of the eukaryotic cell cycle. BMC Biol. 2015, 13, 46. [Google Scholar] [CrossRef] [Green Version]
- Shao, Q.; Cortes, M.G.; Trinh, J.T.; Guan, J.; Balázsi, G.; Zeng, L. Coupling of DNA replication and negative feedback controls gene expression for cell-fate decisions. Iscience 2018, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tyson, J.J.; Baumann, W.T.; Chen, C.; Verdugo, A.; Tavassoly, I.; Wang, Y.; Clarke, R. Dynamic modelling of oestrogen signalling and cell fate in breast cancer cells. Nat. Rev. Cancer 2011, 11, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Pfeuty, B.; David-Pfeuty, T.; Kaneko, K. Underlying principles of cell fate determination during G1 phase of the mammalian cell cycle. Cell Cycle 2008, 7, 3246–3257. [Google Scholar] [CrossRef]
- Pfeuty, B.A. A computational model for the coordination of neural progenitor self-renewal and differentiation through Hes1 dynamics. Development 2015, 142, 3. [Google Scholar] [CrossRef] [Green Version]
- Strogatz, S.H. The Mathematical Structure of the Human Sleep-Wake Cycle; Springer: New York, NY, USA, 1986; Volume 69. [Google Scholar]
- Strogatz, S.H. Human sleep and circadian rhythms: A simple model based on two coupled oscillators. J. Math. Biol. 1987, 25, 327–347. [Google Scholar] [CrossRef]
- Burton, J.; Manning, C.S.; Rattray, M.; Papalopulu, N.; Kursawe, J. Inferring kinetic parameters of oscillatory gene regulation from single cell time-series data. J. R. Soc. Interface 2021, 18, 20210393. [Google Scholar] [CrossRef]
- Paulsson, J. Models of stochastic gene expression. Phys. Life Rev. 2005, 2, 157–175. [Google Scholar] [CrossRef]
- Fan, M.; Kuwahara, H.; Wang, X.; Wang, S.; Gao, X. Parameter estimation methods for gene circuit modeling from time-series mRNA data: A comparative study. Brief. Bioinform. 2015, 16, 987–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gérard, C.; Goldbeter, A. Entrainment of the mammalian cell cycle by the circadian clock: Modeling two coupled cellular rhythms. PLoS Comput. Biol. 2012, 8, e1002516. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Pando, B.F.; Dong, G.; Golden, S.S.; van Oudenaarden, A. Circadian gating of the cell cycle revealed in single cyanobacterial cells. Science 2010, 327, 1522–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feillet, C.; Krusche, P.; Tamanini, F.; Janssens, R.C.; Downey, M.J.; Martin, P.; Rand, D.A. Phase locking and multiple oscillating attractors for the coupled mammalian clock and cell cycle. Proc. Natl. Acad. Sci. USA 2014, 111, 9828–9833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Droin, C.; Paquet, E.R.; Naef, F. Low-dimensional dynamics of two coupled biological oscillators. Nat. Phys. 2019, 15, 1086–1094. [Google Scholar] [CrossRef]
- Tyson, J.J.; Novák, B. Irreversible transitions, bistability and checkpoint controls in the eukaryotic cell cycle: A systems-level understanding. In Handbook of Systems Biology; Elsevier: San Diego, CA, USA, 2012; pp. 265–285. [Google Scholar]
- Pomerening, J.R.; Kim, S.Y.; Ferrell, J.E., Jr. Systems-level dissection of the cell-cycle oscillator: Bypassing positive feedback produces damped oscillations. Cell 2005, 122, 565–578. [Google Scholar] [CrossRef] [Green Version]
- Ferrell, J.E., Jr.; Pomerening, J.R.; Kim, S.Y.; Trunnell, N.B.; Xiong, W.; Huang, C.Y.F.; Machleder, E.M. Simple, realistic models of complex biological processes: Positive feedback and bistability in a cell fate switch and a cell cycle oscillator. FEBS Lett. 2009, 583, 3999–4005. [Google Scholar] [CrossRef] [Green Version]
- Tyson, J.J.; Novak, B. Temporal organization of the cell cycle. Curr. Biol. 2008, 18, R759–R768. [Google Scholar] [CrossRef] [Green Version]
- Pikovsky, A.; Rosenblum, M.; Kurths, J. Synchronization: A Universal Concept in Nonlinear Science; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Strogatz, S.H. Nonlinear Dynamics and Chaos: With Applications to Physics, Biology, Chemistry, and Engineering, 2nd ed.; Westview press: Boulder, CO, USA, 2015; pp. 276–281. [Google Scholar]
- Kuramoto, Y. Self-entrainment of a population of coupled non-linear oscillators International symposium on mathematical problems in theoretical physics. Lect. Notes Phys. 1975, 30, 420. [Google Scholar]
- Morelli, L.G.; Ares, S.; Herrgen, L.; Schröter, C.; Jülicher, F.; Oates, A.C. Delayed coupling theory of vertebrate segmentation. HFSP J. 2009, 3, 55–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biga, V.; Hawley, J.; Soto, X.; Johns, E.; Han, D.; Bennett, H.; Papalopulu, N. A dynamic, spatially periodic, micro-pattern of HES5 underlies neurogenesis in the mouse spinal cord. Mol. Syst. Biol. 2021, 17, e9902. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, X.; Yang, X.; Liu, S.; Jiang, L.; Qu, Y.; Tang, C. Reliable cell cycle commitment in budding yeast is ensured by signal integration. eLife 2015, 4, e03977. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, F.A.; Murray, P.J.; Ditsova, D.; Ferris, M.A.; Davies, P.; Dale, J.K. CDK 1 and CDK 2 regulate NICD 1 turnover and the periodicity of the segmentation clock. EMBO Rep. 2019, 20, e46436. [Google Scholar] [CrossRef]
- Woo, J.H.; Honey, C.J.; Moon, J.Y. Phase and amplitude dynamics of coupled oscillator systems on complex networks. Chaos Interdiscip. J. Nonlinear Sci. 2020, 30, 121102. [Google Scholar] [CrossRef] [PubMed]
- Pietras, B.; Daffertshofer, A. Network dynamics of coupled oscillators and phase reduction techniques. Phys. Rep. 2019, 819, 1–105. [Google Scholar] [CrossRef]
- Ashwin, P.; Bick, C.; Poignard, C. Dead zones and phase reduction of coupled oscillators. Chaos Interdiscip. J. Nonlinear Sci. 2021, 31, 093132. [Google Scholar] [CrossRef]
- Gupta, P.B.; Fillmore, C.M.; Jiang, G.; Shapira, S.D.; Tao, K.; Kuperwasser, C.; Lander, E.S. Stochastic state transitions give rise to phenotypic equilibrium in populations of cancer cells. Cell 2011, 146, 633–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oklejewicz, M.; Destici, E.; Tamanini, F.; Hut, R.A.; Janssens, R.; van der Horst, G.T. Phase resetting of the mammalian circadian clock by DNA damage. Curr. Biol. 2008, 18, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Hong, C.I.; Zámborszky, J.; Csikász-Nagy, A. Minimum criteria for DNA damage-induced phase advances in circadian rhythms. PLoS Comput. Biol. 2009, 5, e1000384. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Modelling Strategy | Coupling Strength Parameter Constraint | Cluster 1 | Cluster 2 | Cluster 3 | Sum |
|---|---|---|---|---|---|
| 1 | 12.1199 | 23.5206 | 6.8118 | 42.4523 | |
| 2 | 8.9520 | 3.0552 | 2.9236 | 14.9308 | |
| 3 | 9.9229 | 1.7894 | 3.0774 | 14.7897 | |
| 4 | 8.8340 | 7.3753 | 2.8757 | 19.085 | |
| 5 | Optimal (no constraints) | 8.8352 | 1.7967 | 2.8702 | 13.5021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rowntree, A.; Sabherwal, N.; Papalopulu, N. Bilateral Feedback in Oscillator Model Is Required to Explain the Coupling Dynamics of Hes1 with the Cell Cycle. Mathematics 2022, 10, 2323. https://doi.org/10.3390/math10132323
Rowntree A, Sabherwal N, Papalopulu N. Bilateral Feedback in Oscillator Model Is Required to Explain the Coupling Dynamics of Hes1 with the Cell Cycle. Mathematics. 2022; 10(13):2323. https://doi.org/10.3390/math10132323
Chicago/Turabian StyleRowntree, Andrew, Nitin Sabherwal, and Nancy Papalopulu. 2022. "Bilateral Feedback in Oscillator Model Is Required to Explain the Coupling Dynamics of Hes1 with the Cell Cycle" Mathematics 10, no. 13: 2323. https://doi.org/10.3390/math10132323
APA StyleRowntree, A., Sabherwal, N., & Papalopulu, N. (2022). Bilateral Feedback in Oscillator Model Is Required to Explain the Coupling Dynamics of Hes1 with the Cell Cycle. Mathematics, 10(13), 2323. https://doi.org/10.3390/math10132323