
Why Is Aedes aegypti Moving South in South America?


Abstract
:1. Introduction
2. Methods
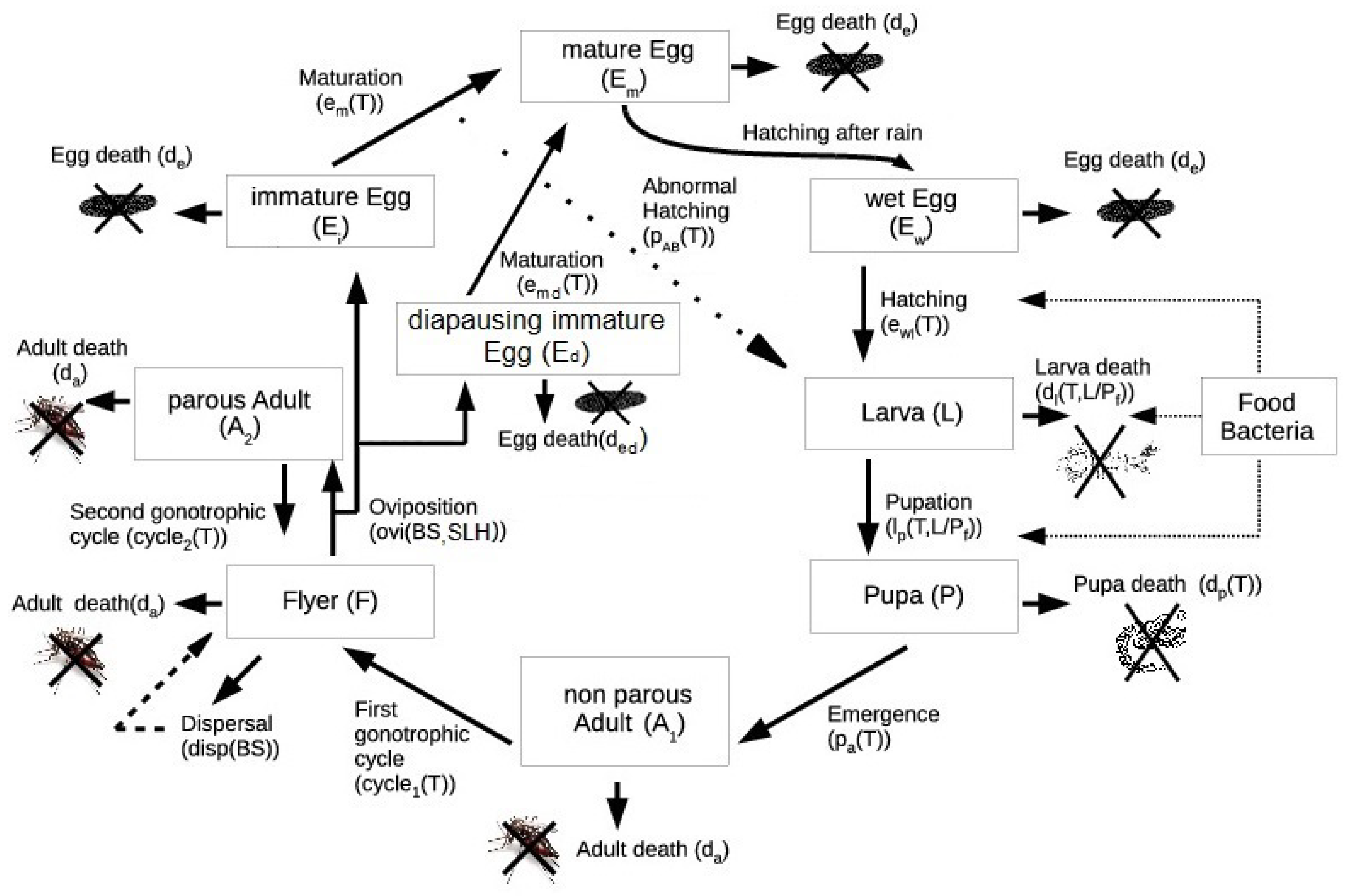
2.1. The Model
2.1.1. Theoretical Biology?
Understanding cannot intuit, and the sensuous faculty cannot think. In no other way than from the united operation of both, can knowledge arise [32].
[Fundamental dialectic] In all human KNOWLEDGE both Thoughts and Things are concerned. In every part of my knowledge there must be some thing about which I know, and an internal act of me who know... Man is interpreting the phenomena which he sees. He often interprets without being aware that he does so [33].
[Speaking of fundamental volition in perception] It is the sense that something has hit me or that I am hitting something; it might be called the sense of collision or clash. It has an outward and an inward variety, corresponding to Kant’s outer and inner sense, to will and self-control, to nerve-action and inhibition, to the two logical types A:B and A:A …
[About empiricists] They often deny this and say they rest entirely on experience. This is because they so overlook the Outward Clash, that they do not know what experience is ([34] CP 8.41) (emphasis added).
Un hecho es, siempre, el producto de la composición entre una parte provista por los objetos y otra construida por el sujeto ([35] Original version).
A fact is always the product of the composition between one part provided by the objects and another constructed by the subject [Our translation].
A man does not need to have seen or experienced everything himself. But if he is to commit himself to another’s experiences and his way of putting them, let him consider that he has to do with three things – the object in question and two subjects ([36] #556).
2.1.2. The Timeline of AedesBA
2.1.3. Current Update
2.2. Input Data
- Daily light hours were calculated using standard methods based on latitude (see [49], Ch 1, §6, eq. 1 and 10) for each city.
- Food dynamics were assumed to depend on temperature in a similar fashion compared to yeast (see [42]), with an optimal temperature of 27 °C and a minimal temperature of 11 °C.
- All other developmental parameters correspond to those reported in [50] for the strain collected in Córdoba, Argentina (31°25 S 64°11 W).
- A “colonized urbanization” of 6 by 10 (6 × 10) blocks was arbitrarily chosen for the simulations. Each block was attributed a food productivity value under optimal weather conditions (27 °C), that was able to sustain the development of 60 larvae in their fourth instar, being the larvae of the largest possible size. All the larvae in a block are assumed to be co-inhabiting a single breeding site.
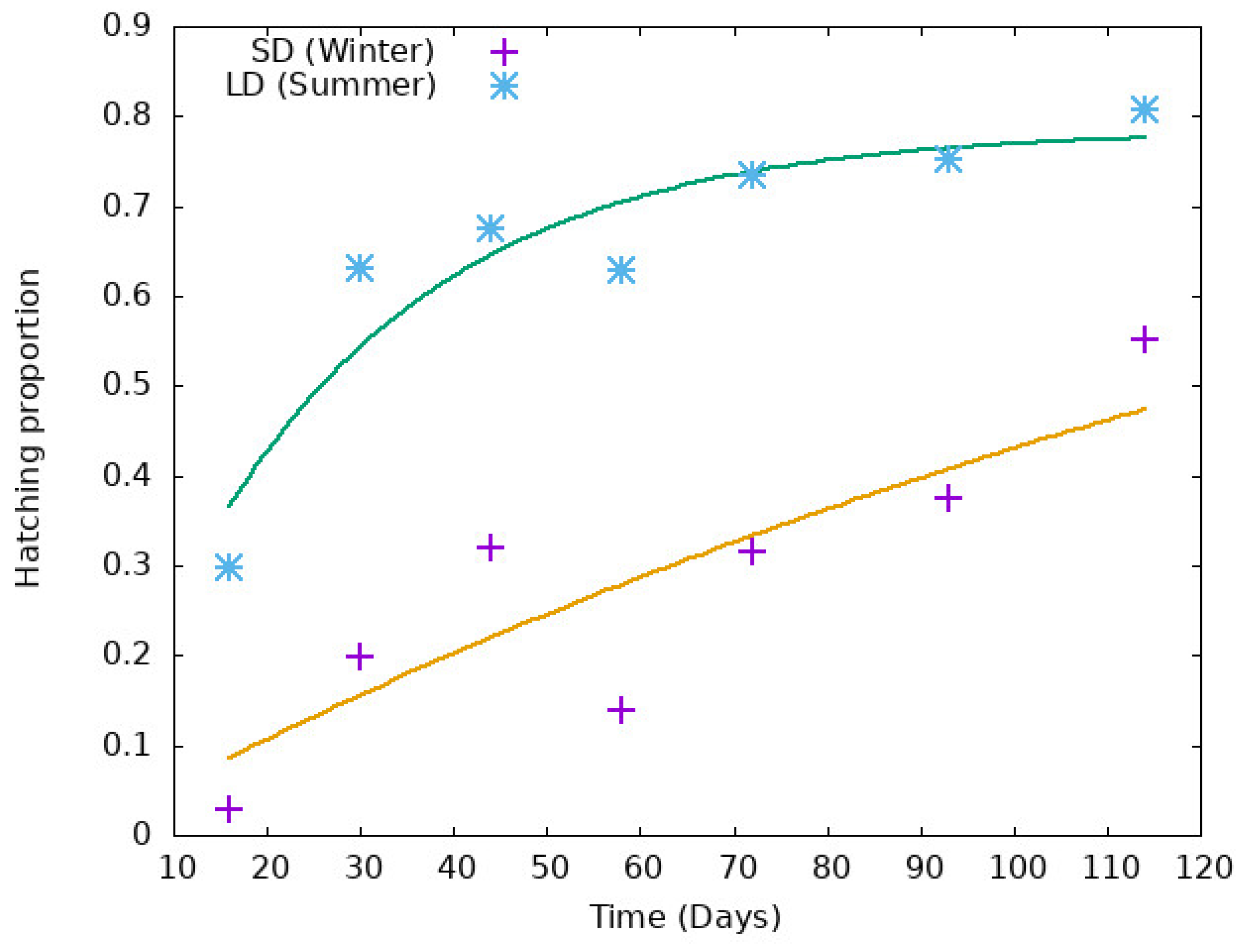
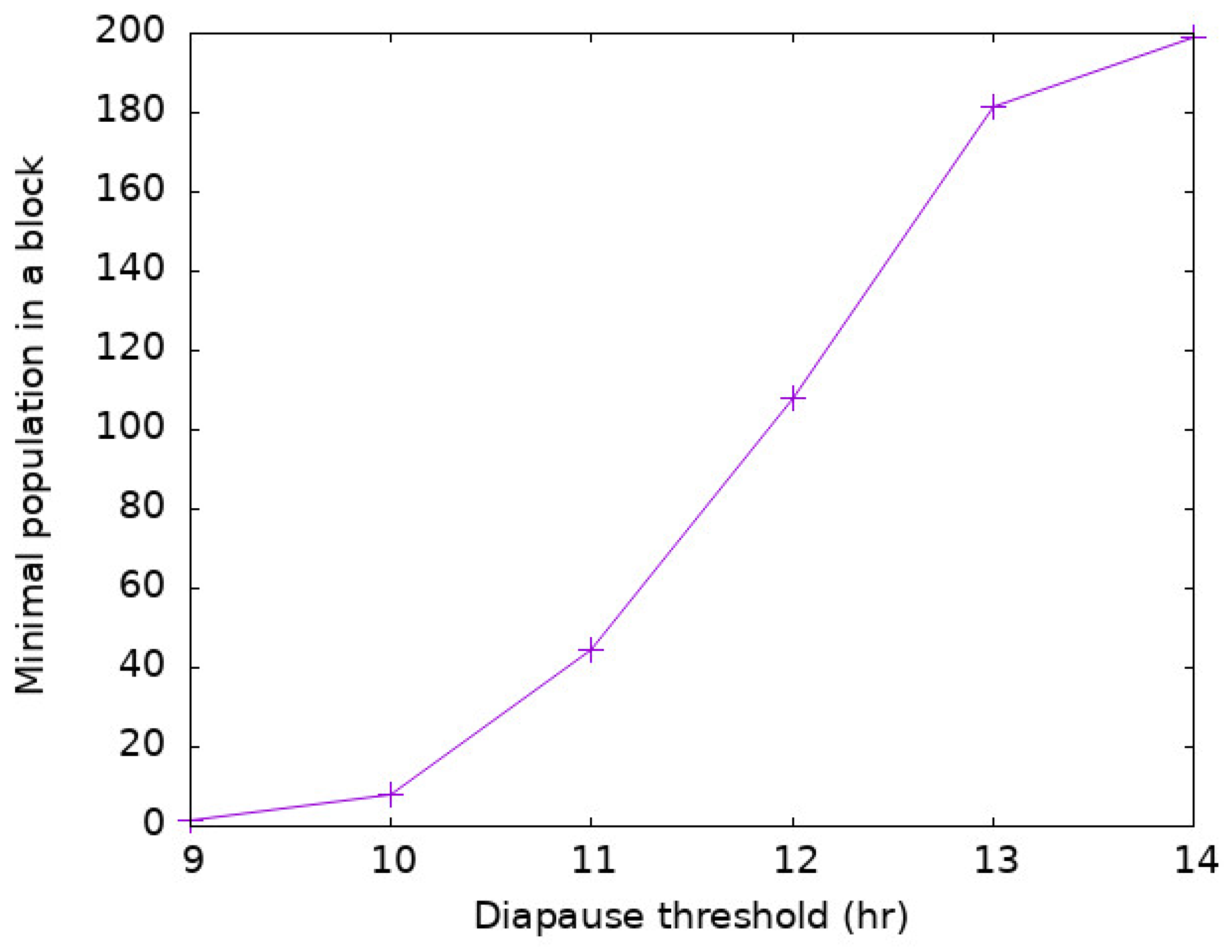
2.3. Modeling Diapause
2.4. Direct and Indirect Model Outputs
3. Results
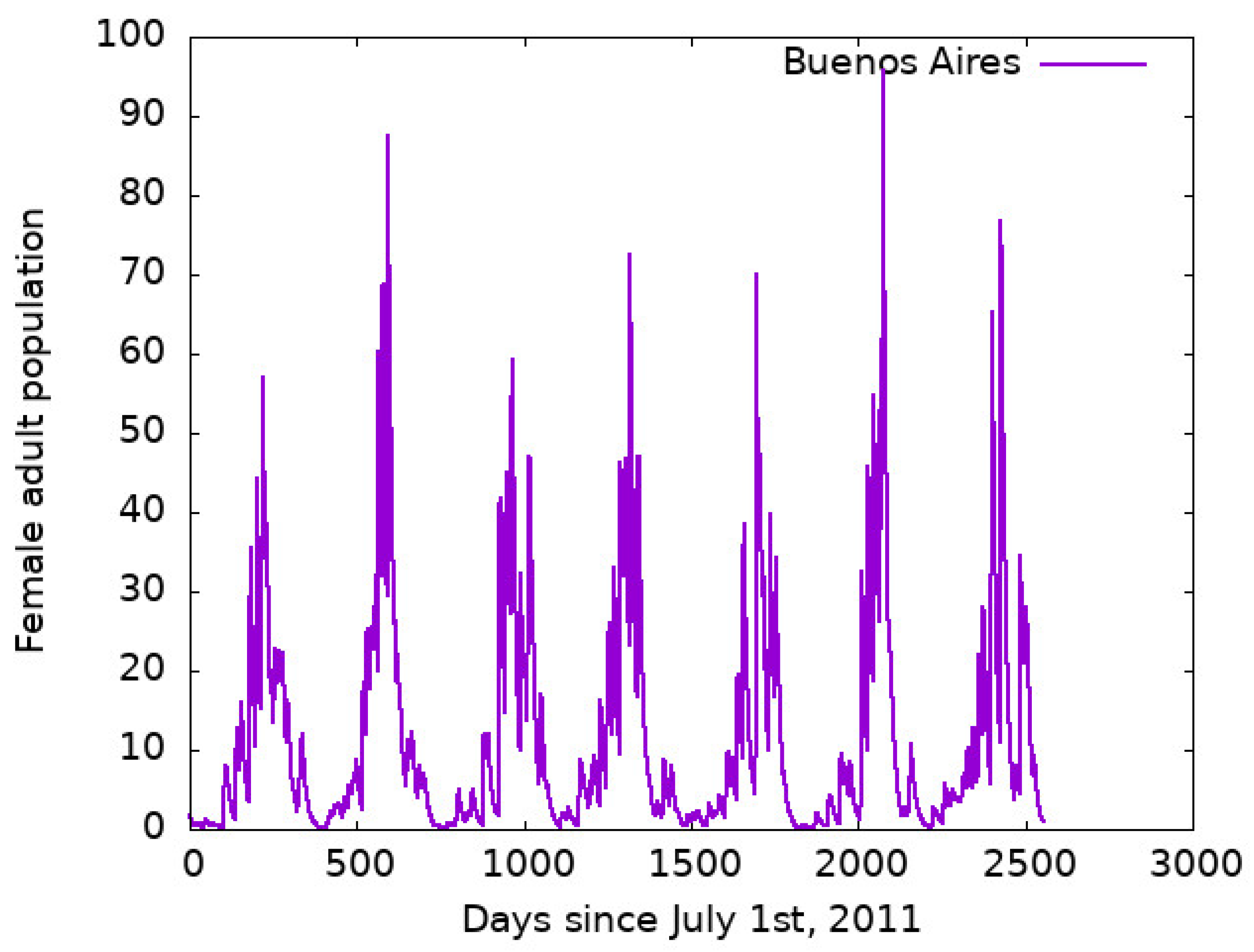
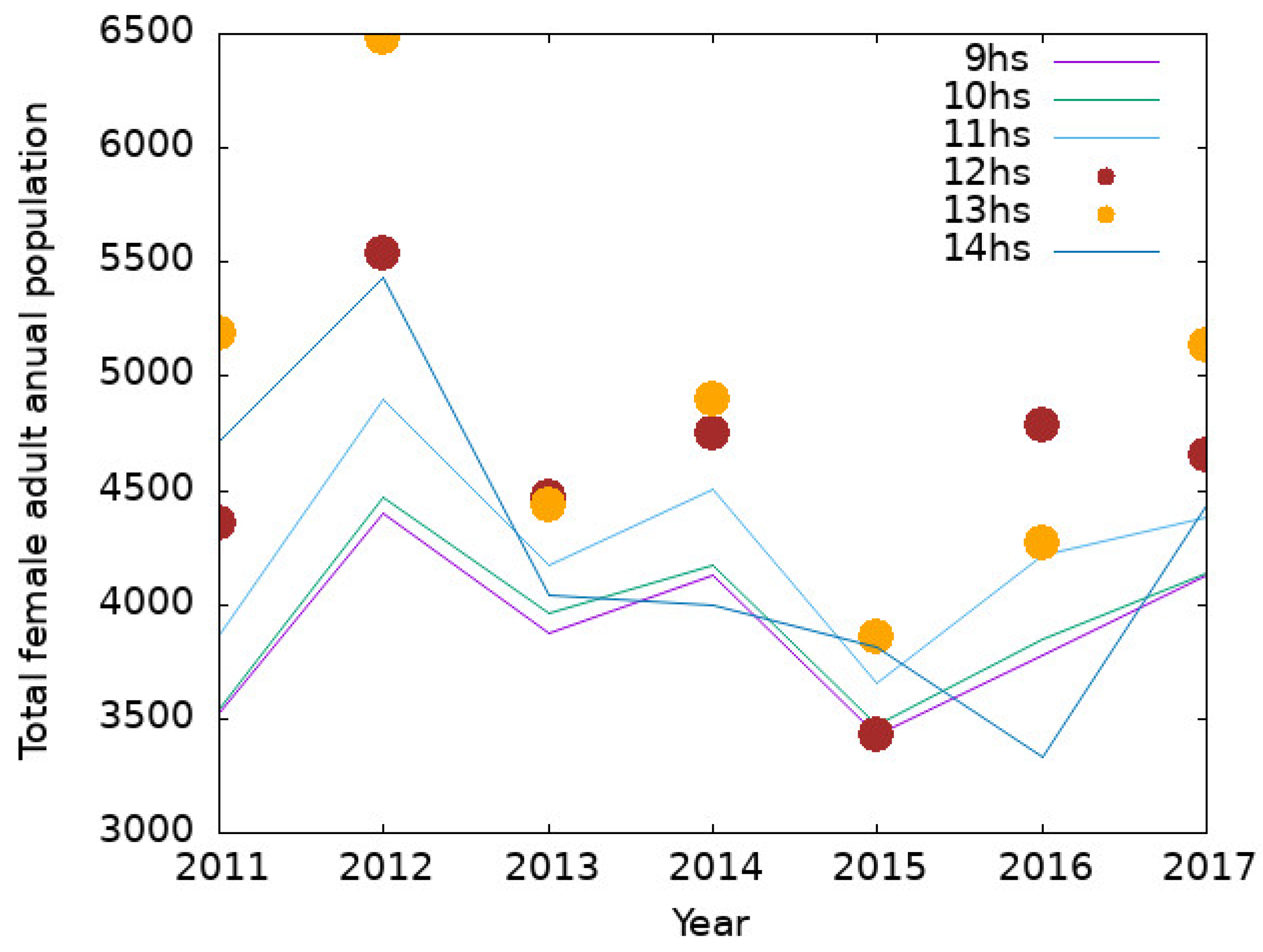
3.1. Outcomes of the Model for Buenos Aires City
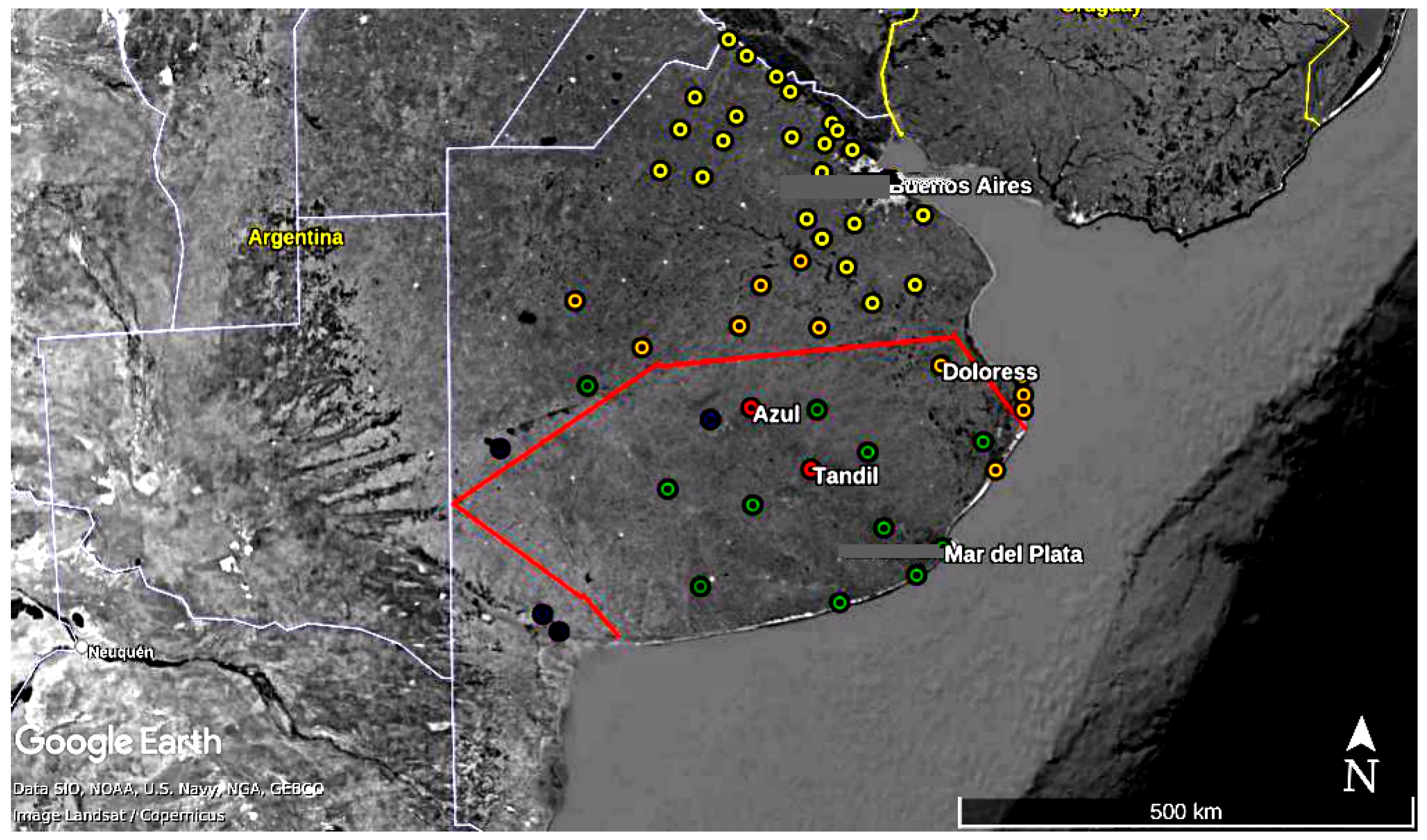
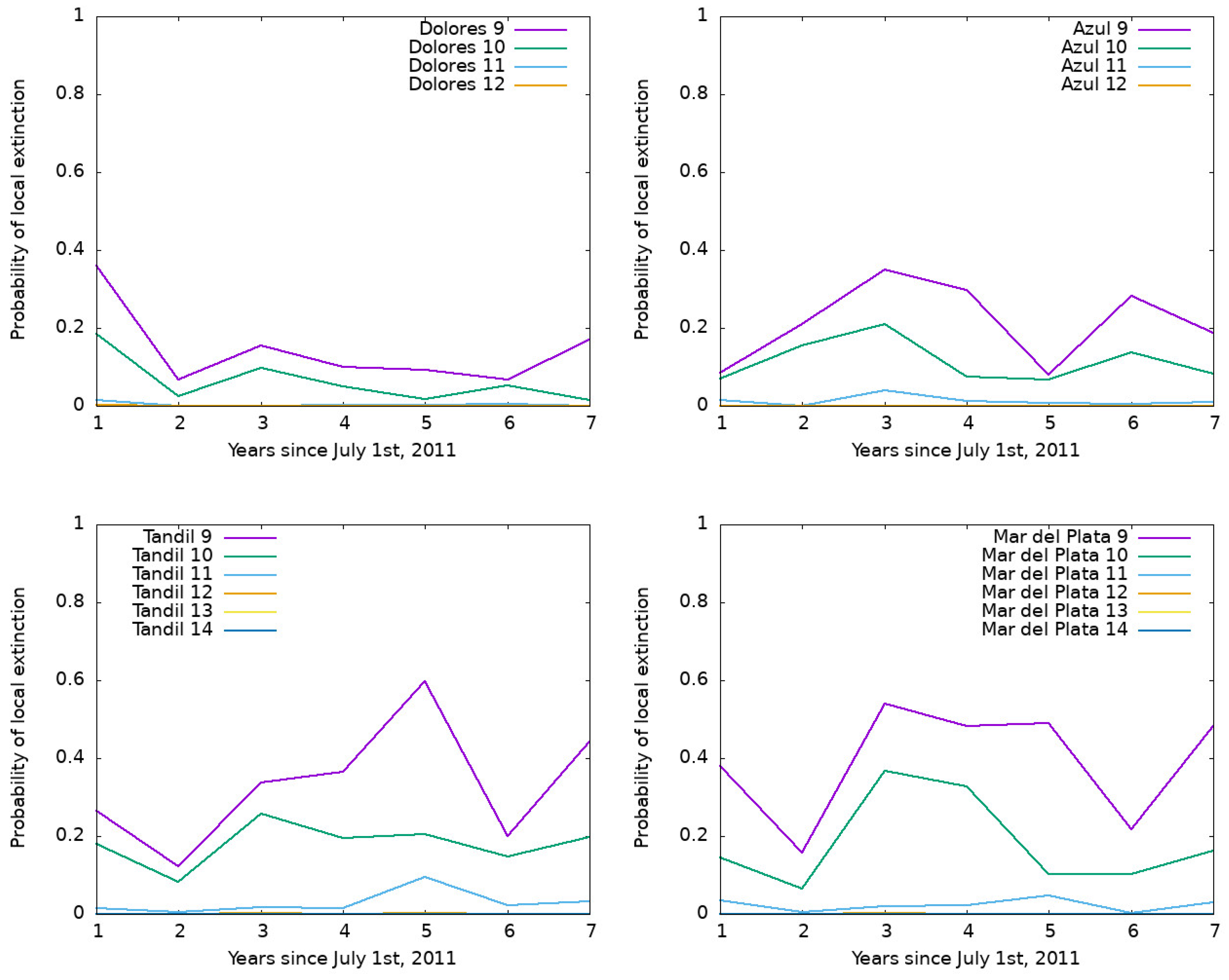
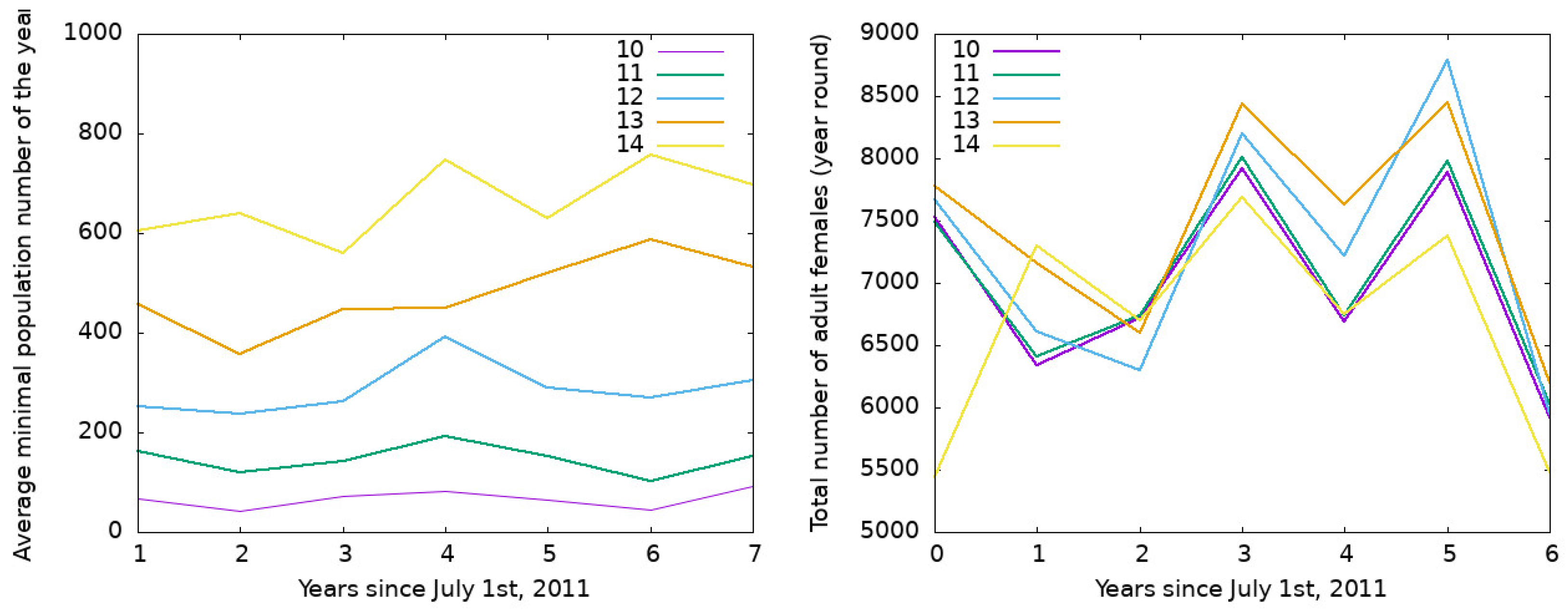
3.2. Output of the Model for Southern Cities in Buenos Aires Province
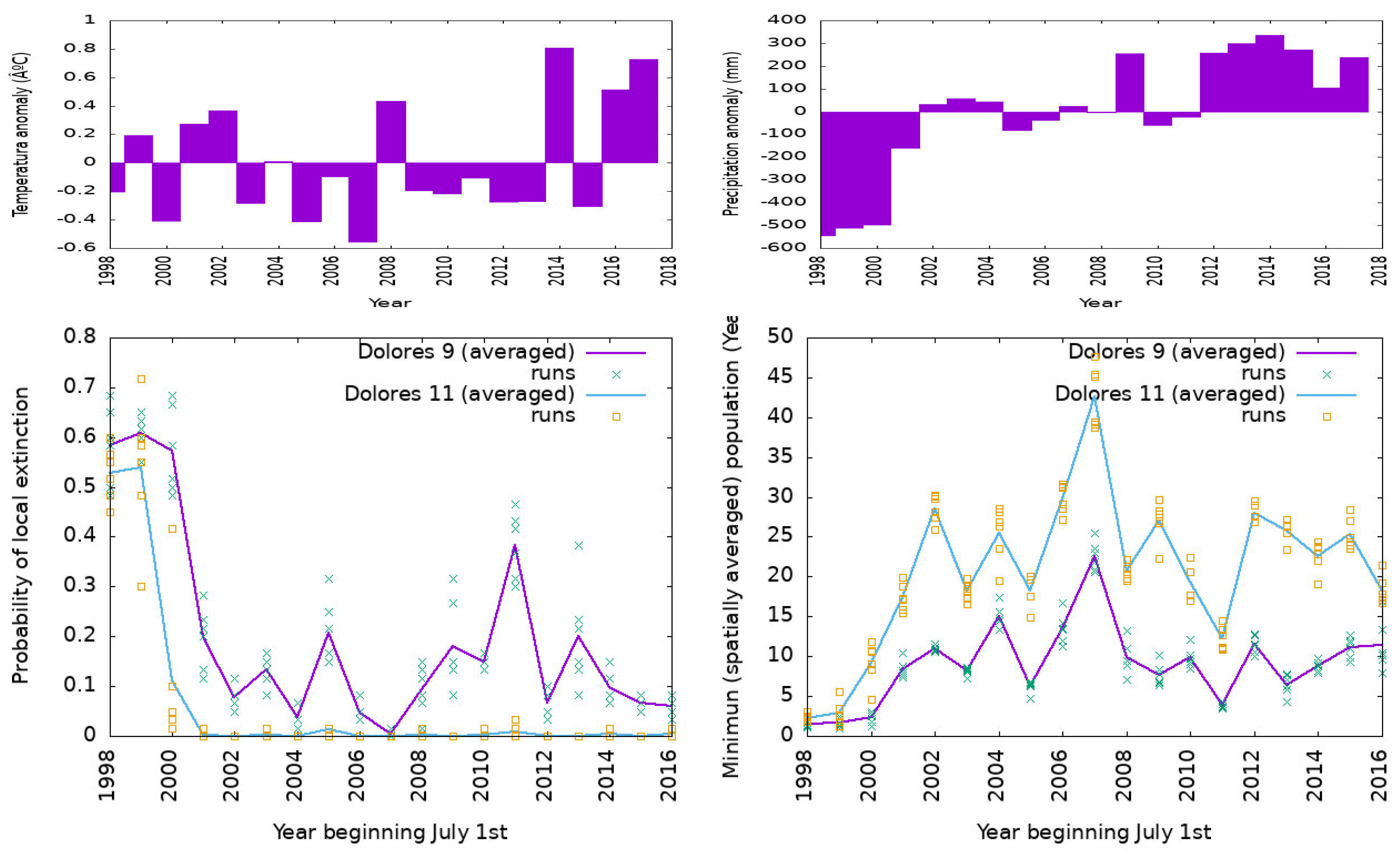
3.3. The 1998–2018 Simulation for Dolores (Buenos Aires)
3.4. The Simulation for Resistencia (Chaco)
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Generalities of the Model

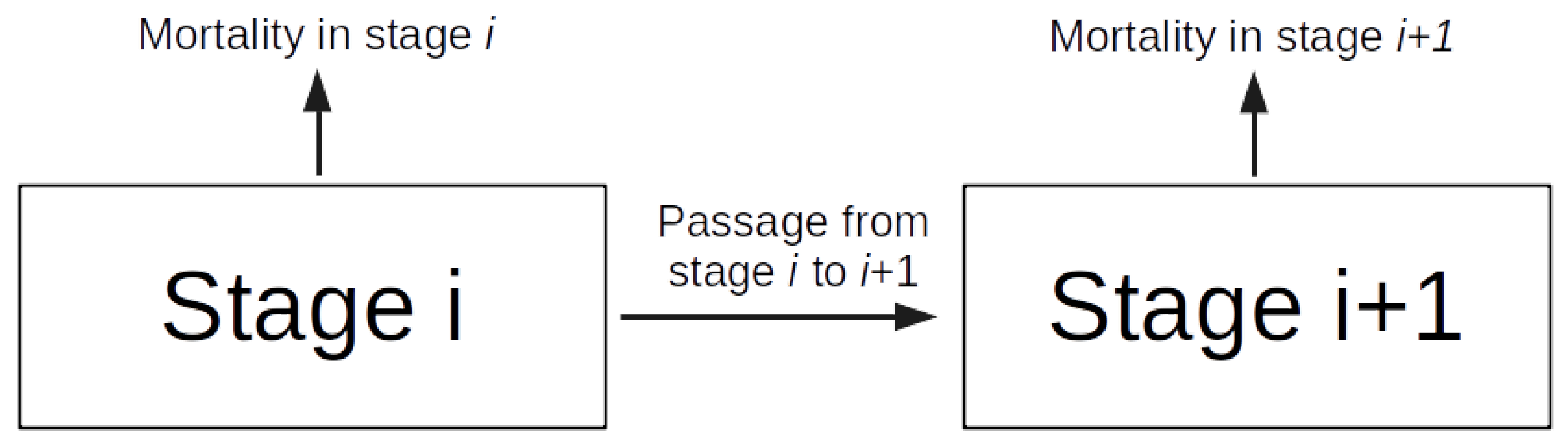
Phases within the Main Ae. aegypti Stages and Rates
- Female adults not having laid eggs, (longer gonadotropic cycle).
- Female adults having laid eggs, (shorter gonadotropic cycle).
- Flyers, F. Females are able to deposit their eggs; they fly in order to find oviposition sites. This stage is responsible for the connection between blocks. The flyers can only relocate to neighboring units.
- Immature eggs, (eggs laid by females),
- Mature eggs, ,
- Wet eggs, .
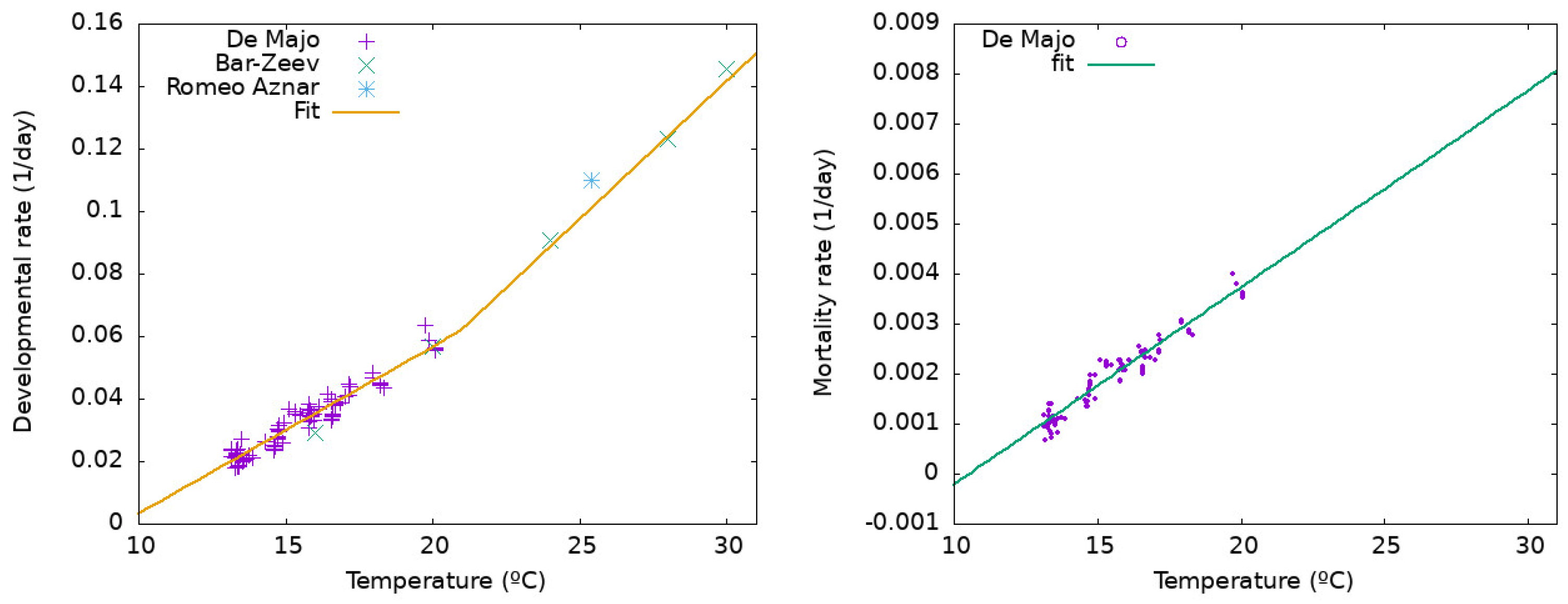
Appendix B. Temperature Dependence of Development and Mortality as a Function of Temperature

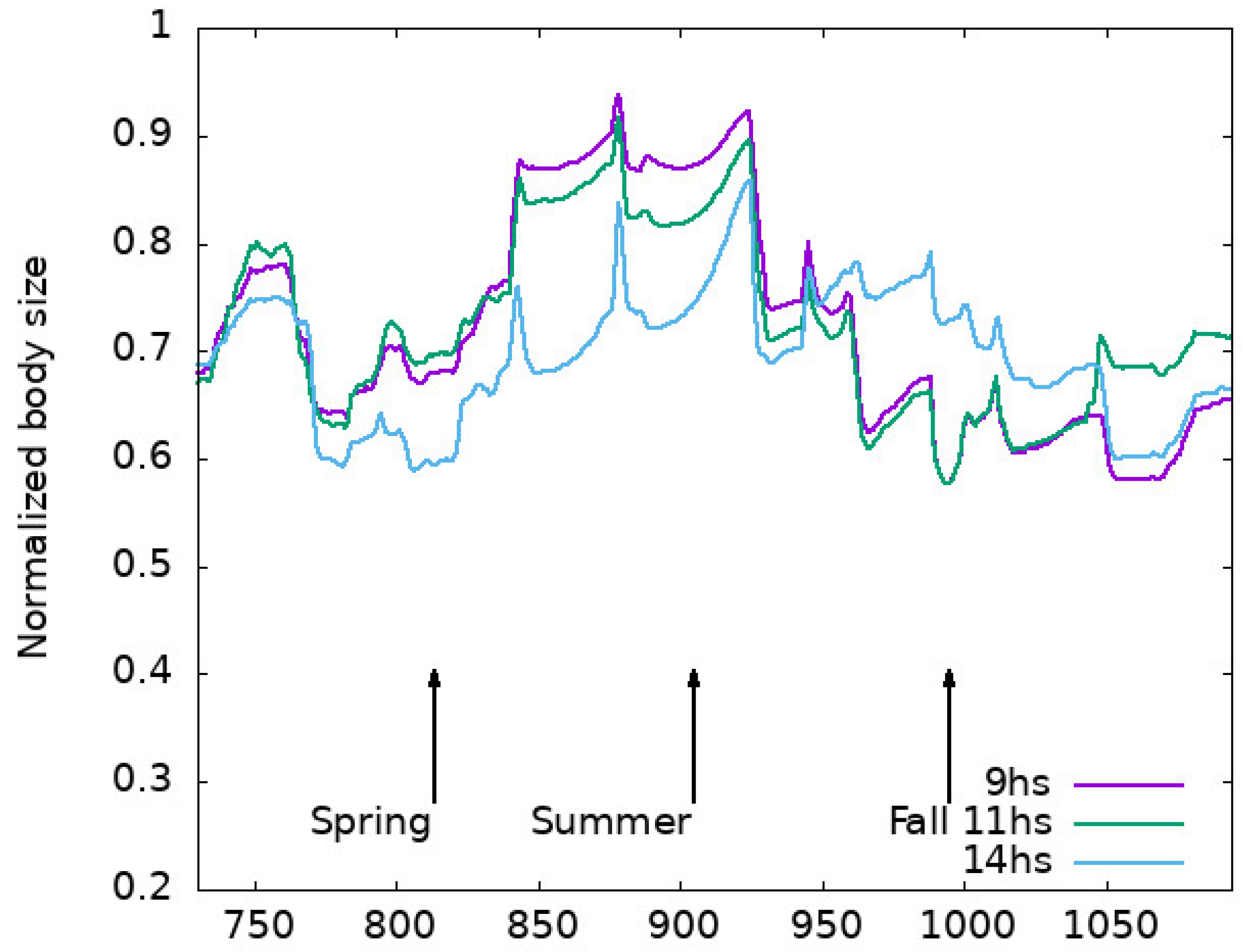
Appendix C. Body Size Evolution

References
- Agramonte, A. An Account of Dr. Louis-Daniel Beauperthuy: A Pioneer in Yellow Fever Research. Boston Med. Surg. J. 1908, CLV111, 928–929. [Google Scholar] [CrossRef] [Green Version]
- Finlay, C. Yellow Fever: Its Transmission By Means of the Culex Mosquito. Am. J. Med. Sci. 1886, 184, 395–408. [Google Scholar] [CrossRef]
- Finlay, C.J. Mosquitoes considered as transmitters of yellow fever and malaria. Psyche 1899, 8, 379–384. [Google Scholar] [CrossRef]
- Reed, W.; Carroll, J.; Agramonte, A.; Lazear, J.W. The Etiology of Yellow Fever—A Preliminary Note. Public Health Pap. Rep. 1900, 26, 37–53. [Google Scholar]
- Carter, H.R. Yellow Fever: An Epidemiological and Historical Study of Its Place of Origin; The Williams & Wilkins Company: Baltimore, MD, USA, 1931. [Google Scholar]
- Christophers, R. Aedes aegypti (L.), the Yellow Fever Mosquito; Cambridge University Press: Cambridge, UK, 1960. [Google Scholar]
- WHO. Dengue Hemorrhagic Fever. Diagnosis, Treatment, Prevention and Control, 2nd ed.; World Health Organization: Geneva, Switzerland, 1998.
- FUNCEI. Dengue enfermedad emergente. Fund. Estud. Infectológicos 1999, 2, 1–12. [Google Scholar]
- Otero, M.; Solari, H.G.; Schweigmann, N. A Stochastic Population Dynamic Model for Aedes aegypti: Formulation and Application to a City with Temperate Climate. Bull. Math. Biol. 2006, 68, 1945–1974. [Google Scholar] [CrossRef]
- Curto, S.; Boffi, R.; Carbajo, A.E.; Plastina, R.; Schweigmann, N.; Salomón, O. Reinfestación del territorio argentino por Aedes aegypti. Distribución geográfica (1994–1999). In Actualizaciones en Artropodología Sanitaria Argentina; Salomón, O., Ed.; Fundación Mundo Sano: Buenos Aires, Argentina, 2002; pp. 127–137. [Google Scholar]
- Zanotti, G.; Majo, D.; Sol, M.; Alem, I.; Schweigmann, N.; Campos, R.E.; Fischer, S. New records of Aedes aegypti at the southern limit of its distribution in Buenos Aires province, Argentina. J. Vector Ecol. 2015, 40, 408–411. [Google Scholar] [CrossRef]
- Carbajo, A.; Cardo, M.; Vezzani, D. Past, present and future of Aedes aegypti in its South American southern distribution fringe: What do temperature and population tell us? Acta Trop. 2019, 190, 149–156. [Google Scholar] [CrossRef]
- Rubio, A.; Cardo, M.V.; Vezzani, D.; Carbajo, A.E. Aedes aegypti spreading in South America: New coldest and southernmost records. Mem. Inst. Oswaldo Cruz 2020, 115, e190496. [Google Scholar] [CrossRef]
- Byttebier, B.; De Majo, M.S.; Fischer, S. Hatching response of Aedes aegypti (Diptera: Culicidae) eggs at low temperatures: Effects of hatching media and storage conditions. J. Med. Entomol. 2014, 51, 97–103. [Google Scholar] [CrossRef] [Green Version]
- De Majo, M.S.; Montini, P.; Fischer, S. Egg hatching and survival of immature stages of Aedes aegypti (Diptera: Culicidae) under natural temperature conditions during the cold season in Buenos Aires, Argentina. J. Med. Entomol. 2017, 54, 106–113. [Google Scholar] [CrossRef]
- Fischer, S.; De Majo, M.S.; Di Battista, C.M.; Montini, P.; Loetti, V.; Campos, R.E. Adaptation to temperate climates: Evidence of photoperiod-induced embryonic dormancy in Aedes aegypti in South America. J. Insect Physiol. 2019, 117, 103887. [Google Scholar] [CrossRef]
- Mensch, J.; Di Battista, C.; De Majo, M.S.; Campos, R.E.; Fischer, S. Increased size and energy reserves in diapausing eggs of temperate Aedes aegypti populations. J. Insect Physiol. 2021, 131, 104232. [Google Scholar] [CrossRef]
- Gillett, J.D. Control of Hatching in Prediapause Eggs of Aedes Mosquitoes. Nature 1959, 104, 1621–1623. [Google Scholar] [CrossRef]
- Hanson, S.M.; Craig, G.B., Jr. Cold acclimation, diapause, and geographic origin affect cold hardiness in eggs of Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 1994, 31, 192–201. [Google Scholar] [CrossRef]
- Zwietering, M.H.; de Koos, J.T.; Hasenack, B.E.; de Witt, J.C.; van’t Riet, K. Modeling of bacterial growth as a function of temperature. Appl. Environ. Microbiol. 1991, 57, 1094–1101. [Google Scholar] [CrossRef] [Green Version]
- Gillett, J.D. Variation in the Hatching-Response of Aedes Eggs (Diptera: Culicidae). Bull. Entomol. Res. 1955, 46, 241–254. [Google Scholar] [CrossRef]
- Gillett, J.D. The Inherited Basis of Variation in the Hatching Response of Aedes Eggs (Diptera: Culicidae). Bull. Entomol. Res. 1955, 46, 255–265. [Google Scholar] [CrossRef]
- Gillett, J.D.; Roman, E.A.; Phillips, V. Erratic Hatching in Aedes Eggs: A New Interpretation. Proc. R. Soc. Lond. B 1977, 196, 223–232. [Google Scholar]
- Carbajo, A.E.; Gomez, S.M.; Curto, S.I.; Schweigmann, N. Variación Espacio Temporal del Riesgo de Transmisión de Dengue en la Ciudad de Buenos Aires. Medicina 2004, 64, 231–234. [Google Scholar]
- Otero, M.; Schweigmann, N.; Solari, H.G. A stochastic spatial dynamical model for Aedes aegypti. Bull. Math. Biol. 2008, 70, 1297–1325. [Google Scholar] [CrossRef] [PubMed]
- Legros, M.; Otero, M.; Romeo Aznar, V.; Solari, H.; Gould, F.; Lloyd, A.L. Comparison of two detailed models of Aedes aegypti population dynamics. Ecosphere 2016, 7, e01515. [Google Scholar] [CrossRef] [PubMed]
- Romeo Aznar, V. Biología teórica, modelo y experimentos aplicados al entendimiento de la dinámica poblacional del mosquito Aedes aegypti. Ph.D. Thesis, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina, 2015. [Google Scholar]
- Romeo Aznar, V.; De Majo, M.S.; Fischer, S.; Natiello, M.A.; Solari, H.G. A model for the development of Aedes (Stegomyia) aegypti (and other insects) as a function of the available food. J. Theor. Biol. 2015, 365, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Romeo Aznar, V.; Alem, I.; De Majo, M.S.; Byttebier, B.; Solari, H.G.; Fischer, S. Effects of scarcity and excess of larval food on life history traits of Aedes aegypti (Diptera: Culicidae). J. Vector Ecol. 2018, 43, 117–124. [Google Scholar] [CrossRef]
- Focks, D.A.; Haile, D.C.; Daniels, E.; Moun, G.A. Dynamics life table model for Aedes aegypti: Analysis of the literature and model development. J. Med. Entomol. 1993, 30, 1003–1018. [Google Scholar] [CrossRef] [PubMed]
- Focks, D.A.; Haile, D.C.; Daniels, E.; Mount, G.A. Dynamic life table model for Aedes aegypti: Simulations results. J. Med. Entomol. 1993, 30, 1019–1029. [Google Scholar]
- Kant, I. The Critique of Pure Reason; Meiklejohn, J.M.D., Translator; An Electronic Classics Series Publication; Manis, J., Ed.; PSU-Hazleton: Hazleton, PA, USA, 2010. [Google Scholar]
- Whewell, W. The History of Scientific Ideas, 3rd ed.; JW Parker: London, UK, 1858; Volume 1. [Google Scholar]
- Peirce, C. Collected Papers of Charles Sanders Peirce; InteLex Corporation: Charlottesville, VA, USA, 1994. [Google Scholar]
- Piaget, J.; García, R. Psicogénesis e Historia de la Ciencia; Siglo XXI: Madrid, Spain, 1982. [Google Scholar]
- von Goethe, J. The Maxims and Reflections of Goethe; The MacMillan Company: London, UK, 1906. [Google Scholar]
- Solari, H.G.; Natiello, M. Science, Dualities and the Fenomenological Map. Found. Sci. 2022. [Google Scholar] [CrossRef]
- Durrett, R. Essentials of Stochastic Processes; Springer: New York, NY, USA, 2001. [Google Scholar]
- Solari, H.G.; Natiello, M.A. Stochastic Population Dynamics: The Poisson Approximation. Phys. Rev. E 2003, 67, 031918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergero, P.; Ruggerio, C.; Lombardo, R.; Schweigmann, N.; Solari, H. Dispersal of Aedes aegypti: Field study in temperate areas and statistical approach. J. Vector Borne Dis. 2013, 50, 163–170. [Google Scholar] [CrossRef]
- Romeo Aznar, V. El Efecto de la Lluvia sobre la Eclosión de Huevos de Aedes aegypti. Estudio e Incorporación a un Modelo. Master’s Thesis, Departamento de Física, Universidad de Buenos Aires, Buenos Aires, Argentina, 2010. [Google Scholar]
- Romeo Aznar, V.; Otero, M.J.; de Majo, M.S.; Fischer, S.; Solari, H.G. Modelling the Complex Hatching and Development of Aedes aegypti in Temperated Climates. Ecol. Model. 2013, 253, 44–55. [Google Scholar] [CrossRef]
- Powell, J. Genetic Variation in Insect Vectors: Death of Typology? Insects 2018, 9, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otero, M.; Solari, H.G. Mathematical model of dengue disease transmission by Aedes aegypti mosquito. Math. Biosci. 2010, 223, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Focks, D.A.; Haile, D.C.; Daniels, E.; Keesling, D. A simulation model of the epidemiology of urban dengue fever: Literature analysis, model development, preliminary validation and samples of simulation results. Am. J. Trop. Med. Hyg. 1995, 53, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Seijo, A.; Romer, Y.; Espinosa, M.; Monroig, J.; Giamperetti, S.; Ameri, D.; Antonelli, L. Brote de Dengue Autóctono en el Area Metropolitana Buenos Aires. Experiencia del Hospital de Enfermedades Infecciosas F. J. Muñiz. Medicina 2009, 69, 593–600. [Google Scholar]
- Fernández, M.L.; Otero, M.; Schweigmann, N.; Solari, H.G. A mathematically assisted reconstruction of the initial focus of the yellow fever outbreak in Buenos Aires (1871). Pap. Phys. 2013, 5, 050002. [Google Scholar] [CrossRef] [Green Version]
- Fischer, S.; Departamento de Ecología, Genética y Evolución, FCEN-UBA. Private Communication, 2018.
- Duffie, J.A.; Beckman, W.A. Solar Engineering of Thermal Processes, 4th ed.; Wiley: Hoboken, NJ, USA, 2013. [Google Scholar]
- Grech, M.G.; Ludueña-Almeida, F.; Almirón, W.R. Bionomics of Aedes aegypti subpopulations (Diptera: Culicidae) from Argentina. J. Vector Ecol. 2010, 35, 277–285. [Google Scholar] [CrossRef]
- Fischer, S.; Alem, I.S.; De Majo, M.S.; Campos, R.E.; Schweigmann, N. Cold season mortality and hatching behavior of Aedes aegypti L.(Diptera: Culicidae) eggs in Buenos Aires City, Argentina. J. Vector Ecol. 2011, 36, 94–99. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.M.; Vera, N.S.; Chiappero, M.B.; Almirón, W.R.; Gardenal, C.N. Urban Populations of Aedes aegypti (Diptera: Culicidae) From Central Argentina: Dispersal Patterns Assessed by Bayesian and Multivariate Methods. J. Med. Entomol. 2020, 57, 1069–1076. [Google Scholar] [CrossRef] [Green Version]
- Tittarelli, E.; Lusso, S.B.; Goya, S.; Rojo, G.L.; Natale, M.I.; Viegas, M.; Mistchenko, A.S.; Valinotto, L.E. Dengue Virus 1 Outbreak in Buenos Aires, Argentina, 2016. Emerg. Infect. Dis. 2017, 23, 1684. [Google Scholar] [CrossRef]
- Barrera, R.; Amador, M.; MacKay, A.J. Population dynamics of Aedes aegypti and dengue as influenced by weather and human behavior in San Juan, Puerto Rico. PLoS Neglected Trop. Dis. 2011, 5, e1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ethier, S.N.; Kurtz, T.G. Markov Processes; John Wiley and Sons: New York, NY, USA, 1986. [Google Scholar]
- Merritt, R.W.; Dadd, R.H.; Walker, E.D. Feeding behavior, natural food, and nutritional relationships of larval mosquitoes. Annu. Rev. Entomol. 1992, 37, 349–376. [Google Scholar] [CrossRef] [PubMed]
- Zwietering, M.H.; de Wit, J.C.; Cuppers, H.G.; van’t Riet, K. Modeling of bacterial growth with shifts in temperature. Appl. Environ. Microbiol. 1994, 60, 204–213. [Google Scholar] [CrossRef] [Green Version]
- von Bertalanffy, L. Principles and theories of growth. In Fundamental Aspects of Normal and Malignant Growth; Nowinski, W., Ed.; Elsevier: Amsterdam, The Netherlands, 1960. [Google Scholar]
- Bar-Zeev, M. The effect of temperature on the growth rate and survival of the immature stages of Aedes aegypti. Bull. Entomol. Res. 1958, 49, 157–163. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| City | Latitude | Longitude | Station (OACI) | Period |
|---|---|---|---|---|
| Buenos Aires | 34°35 S | 58°29 O | 87585 (SABA) | 1981–2018 |
| Dolores | 31°57 S | 65°09 O | 87648 (SAZD) | 1998–2018 |
| Azul | 36°50 S | 59°53 O | 87641 (SAZA) | 2008–2018 |
| Tandil | 37°14 S | 57°14 O | 87645 (SAZT) | 2008–2018 |
| Mar del Plata | 37°56 S | 57°35 O | 87692 (SAZM) | 2008–2018 |
| Resistencia (Chaco) | 27°26 S | 59°03 W | 87155 (SARE) | 2008–2018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso, L.E.; Romeo Aznar, V.; Solari, H.G. Why Is Aedes aegypti Moving South in South America? Mathematics 2022, 10, 4510. https://doi.org/10.3390/math10234510
Alonso LE, Romeo Aznar V, Solari HG. Why Is Aedes aegypti Moving South in South America? Mathematics. 2022; 10(23):4510. https://doi.org/10.3390/math10234510
Chicago/Turabian StyleAlonso, Lucas Ernesto, Victoria Romeo Aznar, and Hernán Gustavo Solari. 2022. "Why Is Aedes aegypti Moving South in South America?" Mathematics 10, no. 23: 4510. https://doi.org/10.3390/math10234510
APA StyleAlonso, L. E., Romeo Aznar, V., & Solari, H. G. (2022). Why Is Aedes aegypti Moving South in South America? Mathematics, 10(23), 4510. https://doi.org/10.3390/math10234510