
Dynamic Image Representation in a Spiking Neural Network Supplied by Astrocytes



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
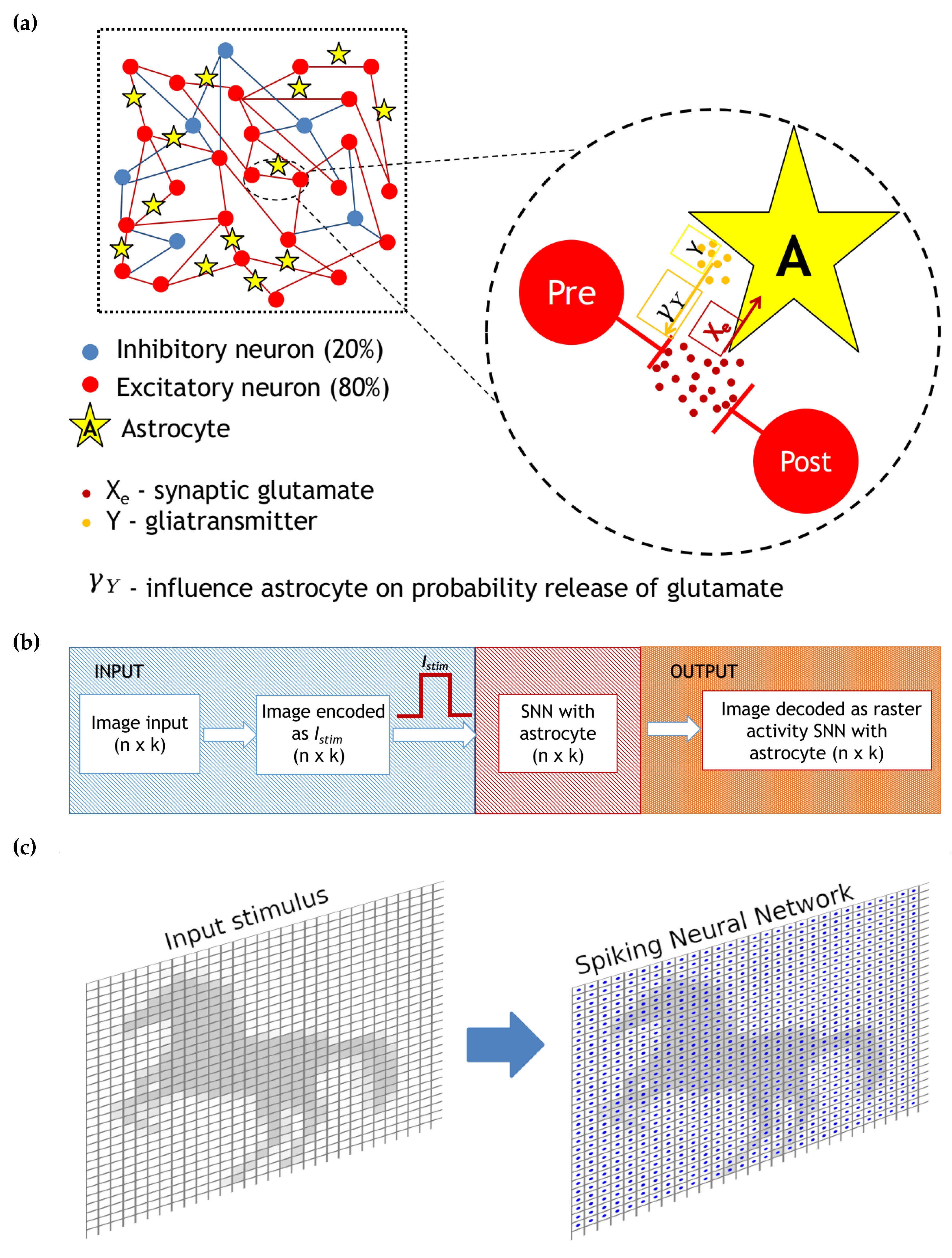
2. The Model
2.1. Mathematical Model of a Single Neuron
2.2. Applied Currents
2.3. Tripartite Synapses
2.4. Neural Network
2.5. Numerical Simulation Method
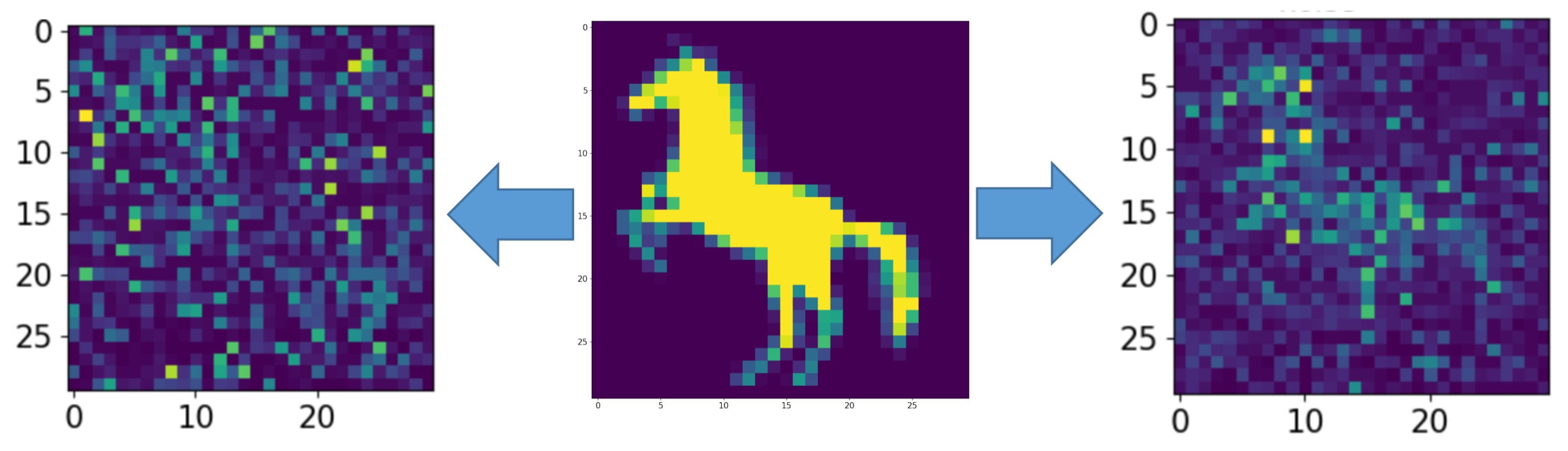
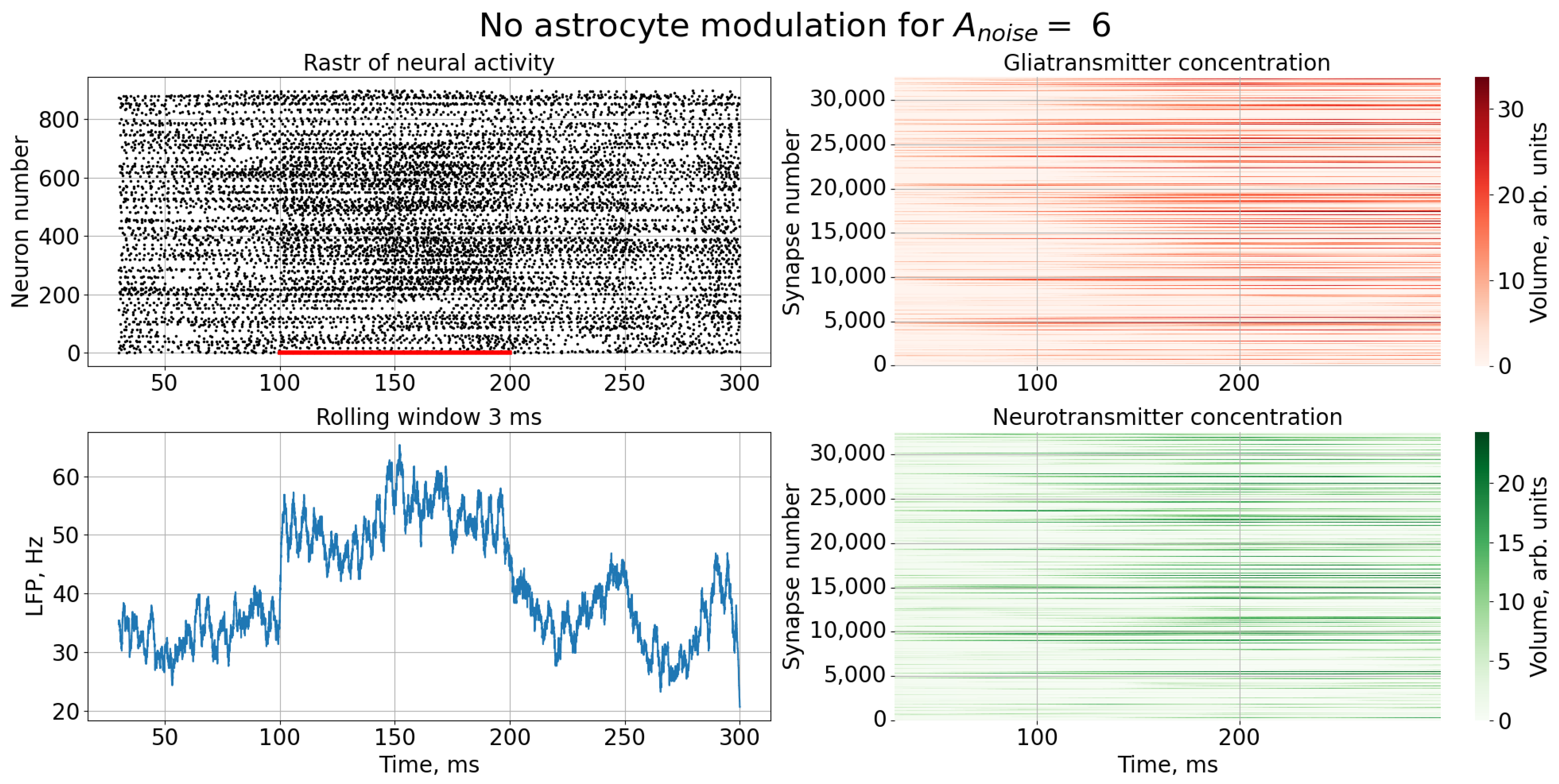
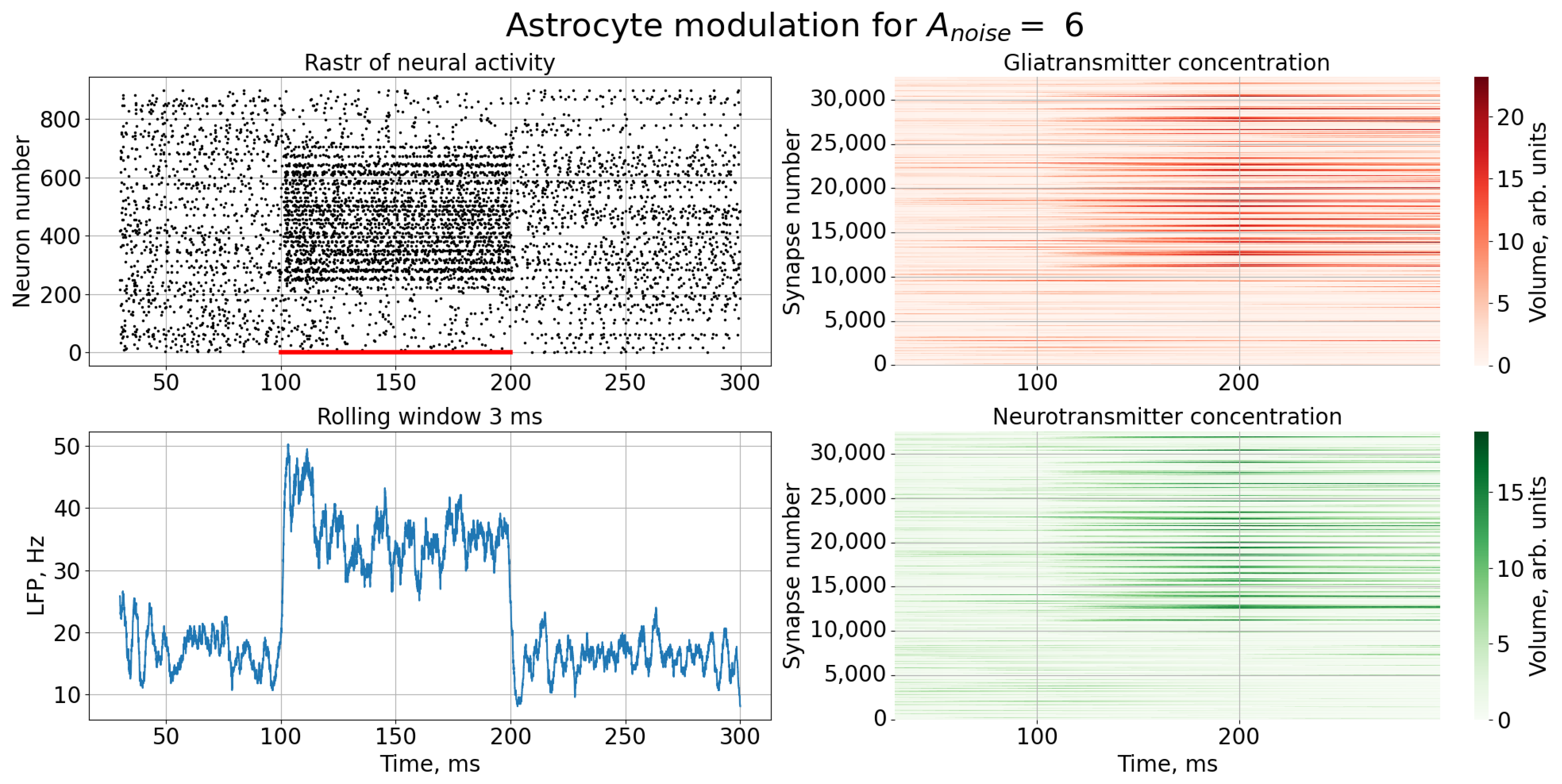
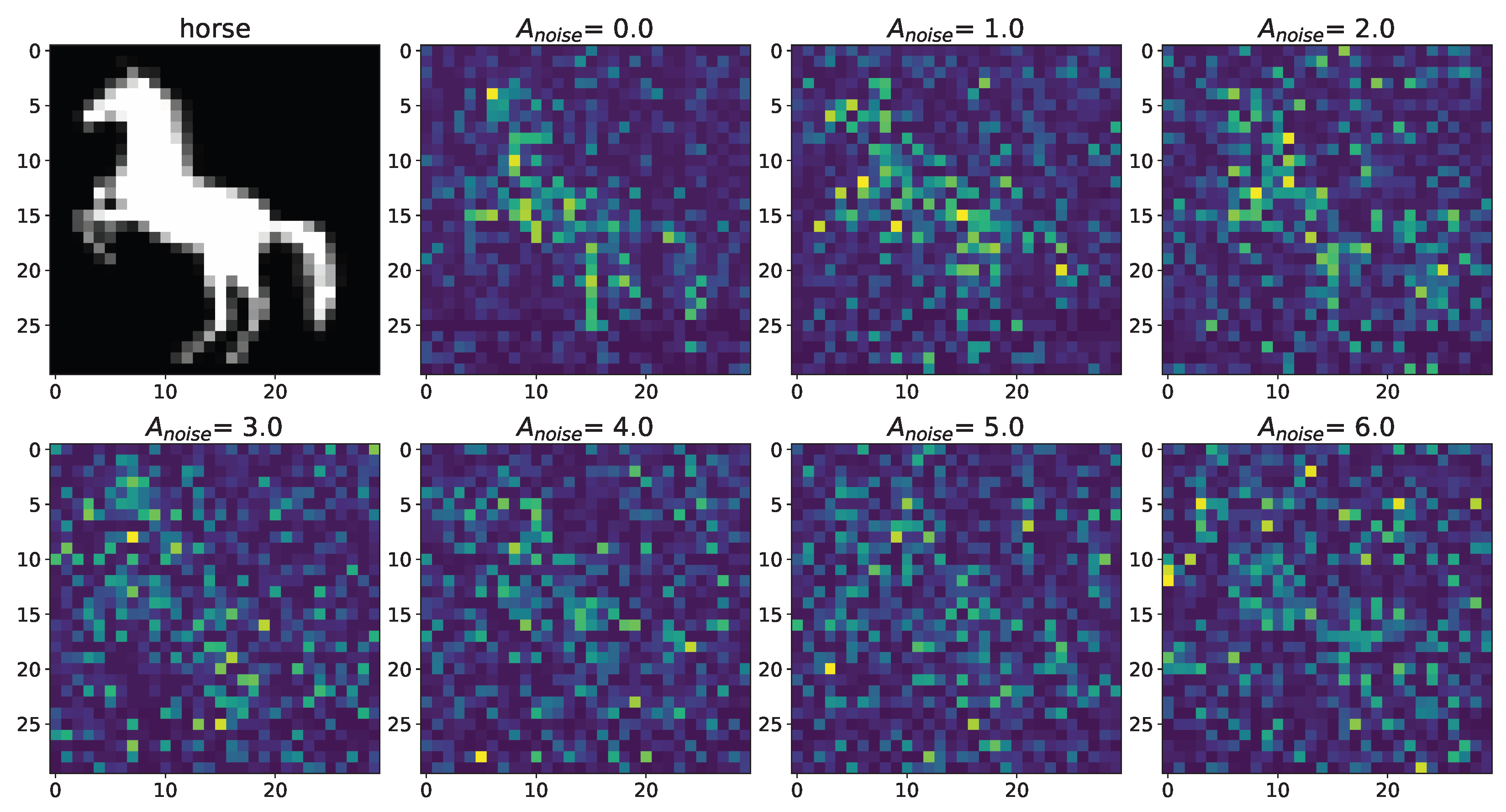
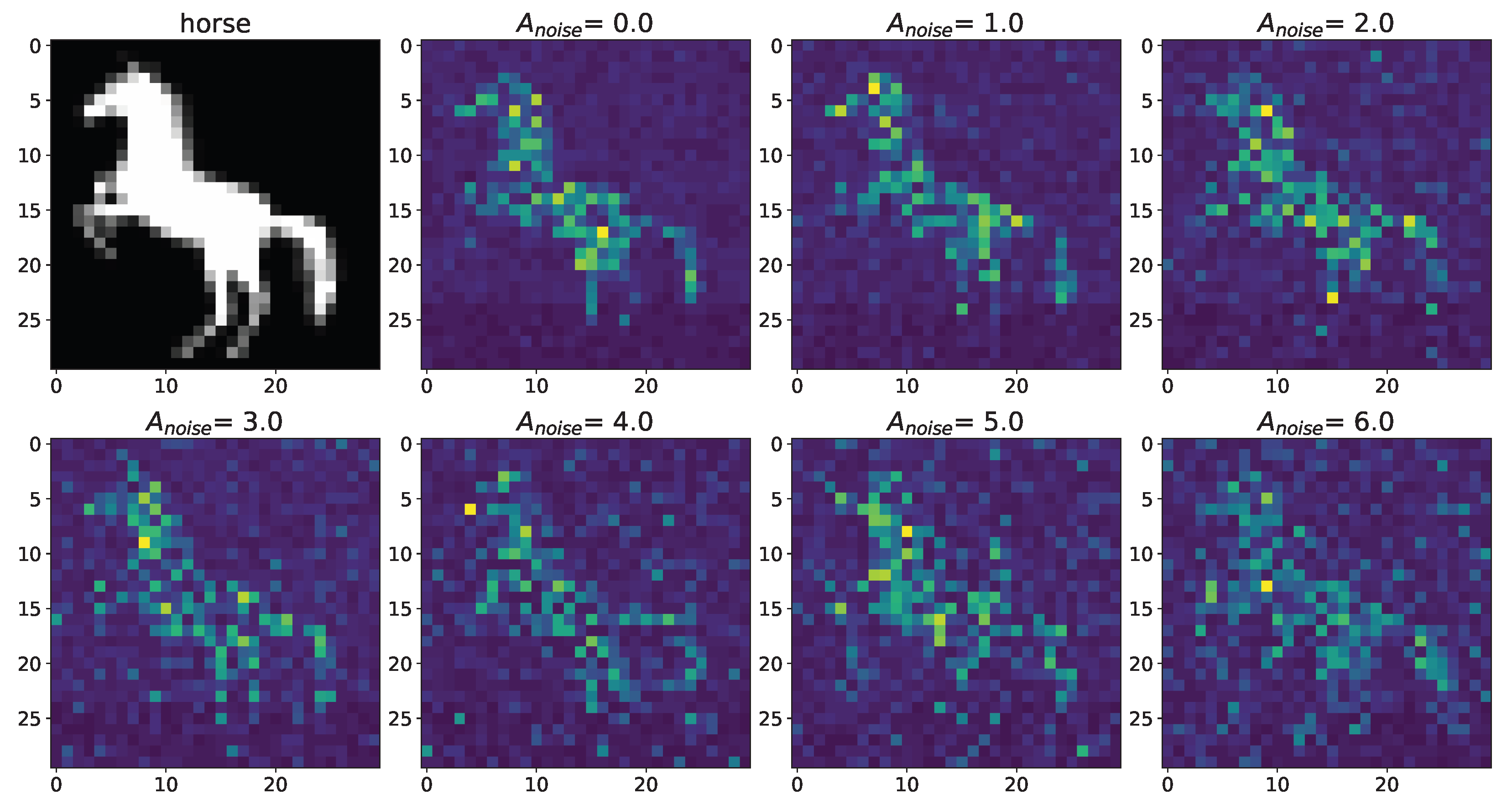
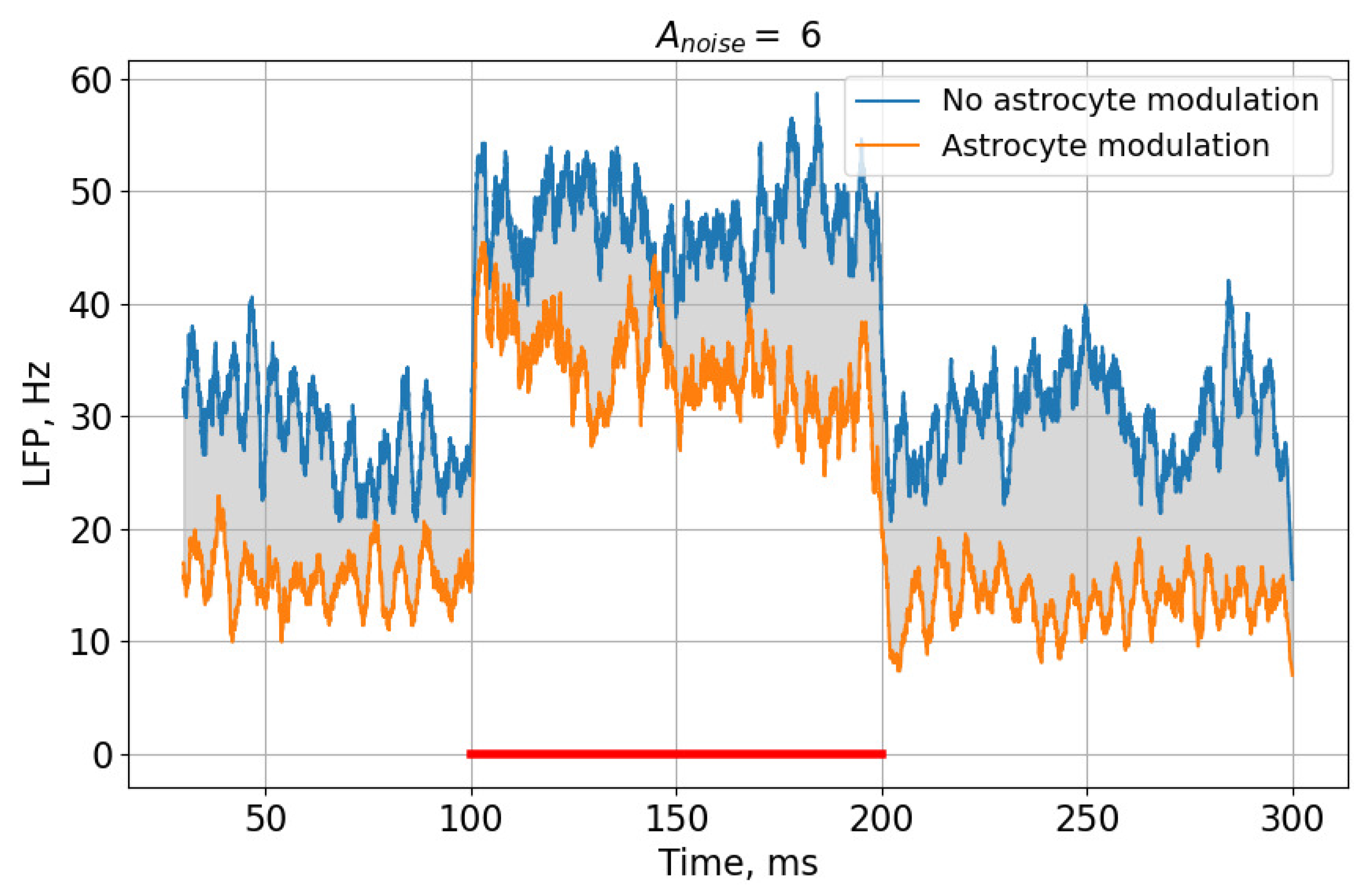
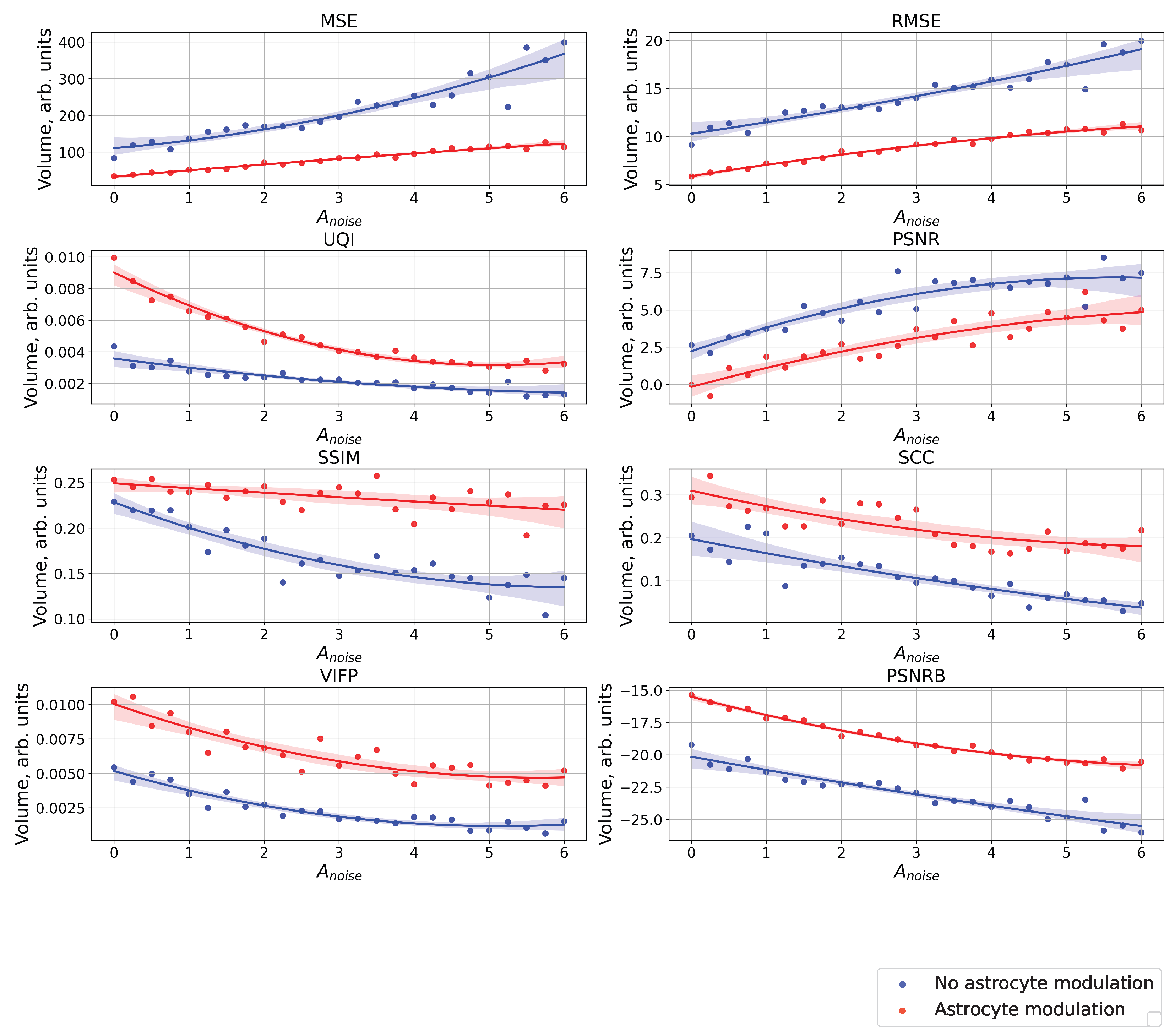
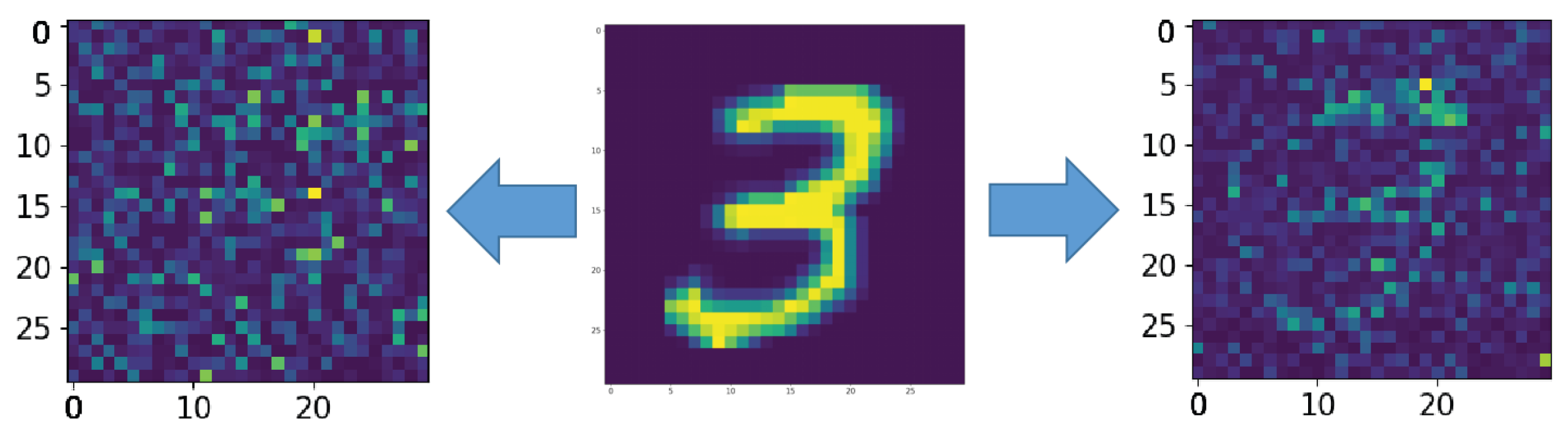
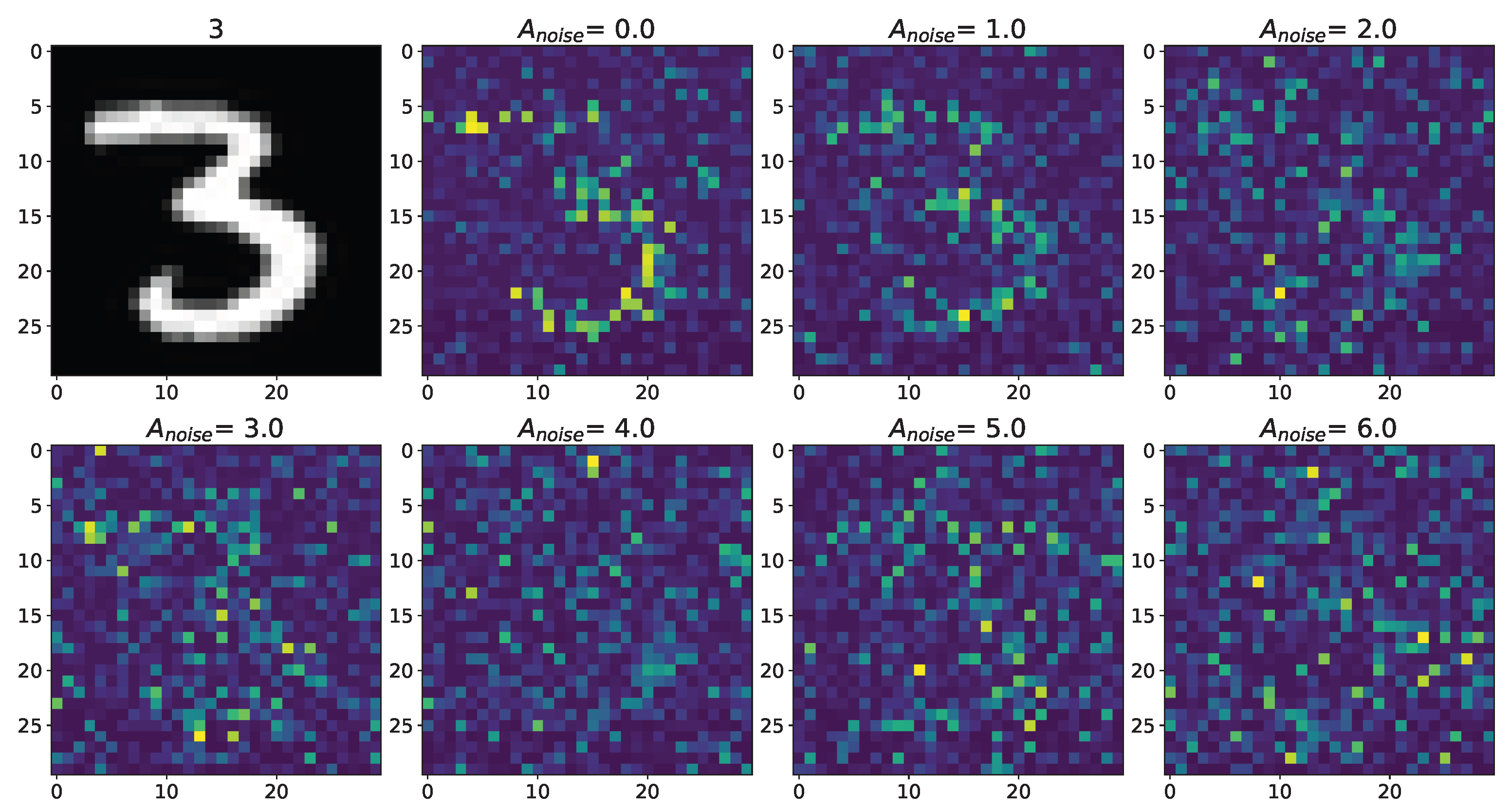
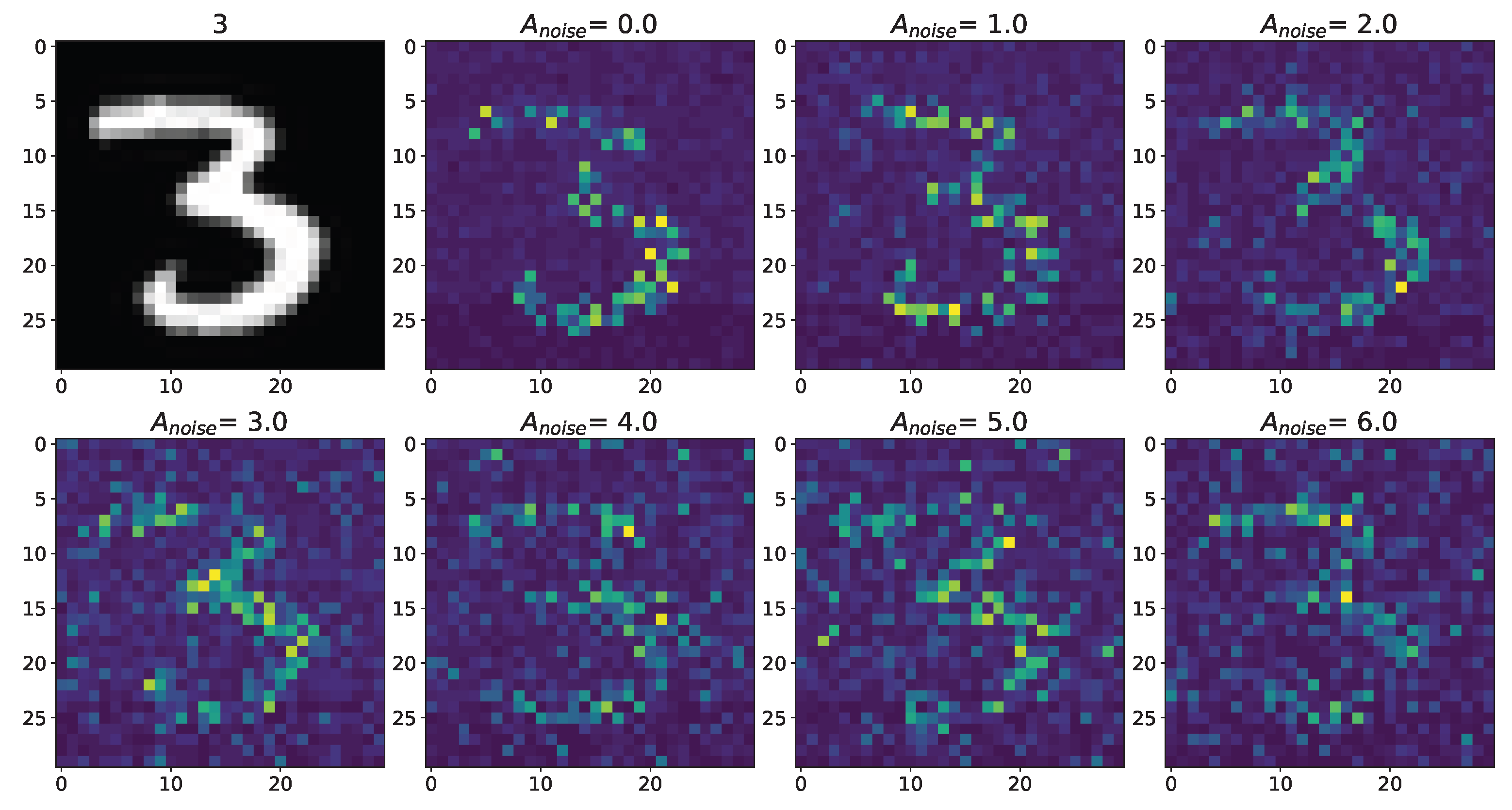
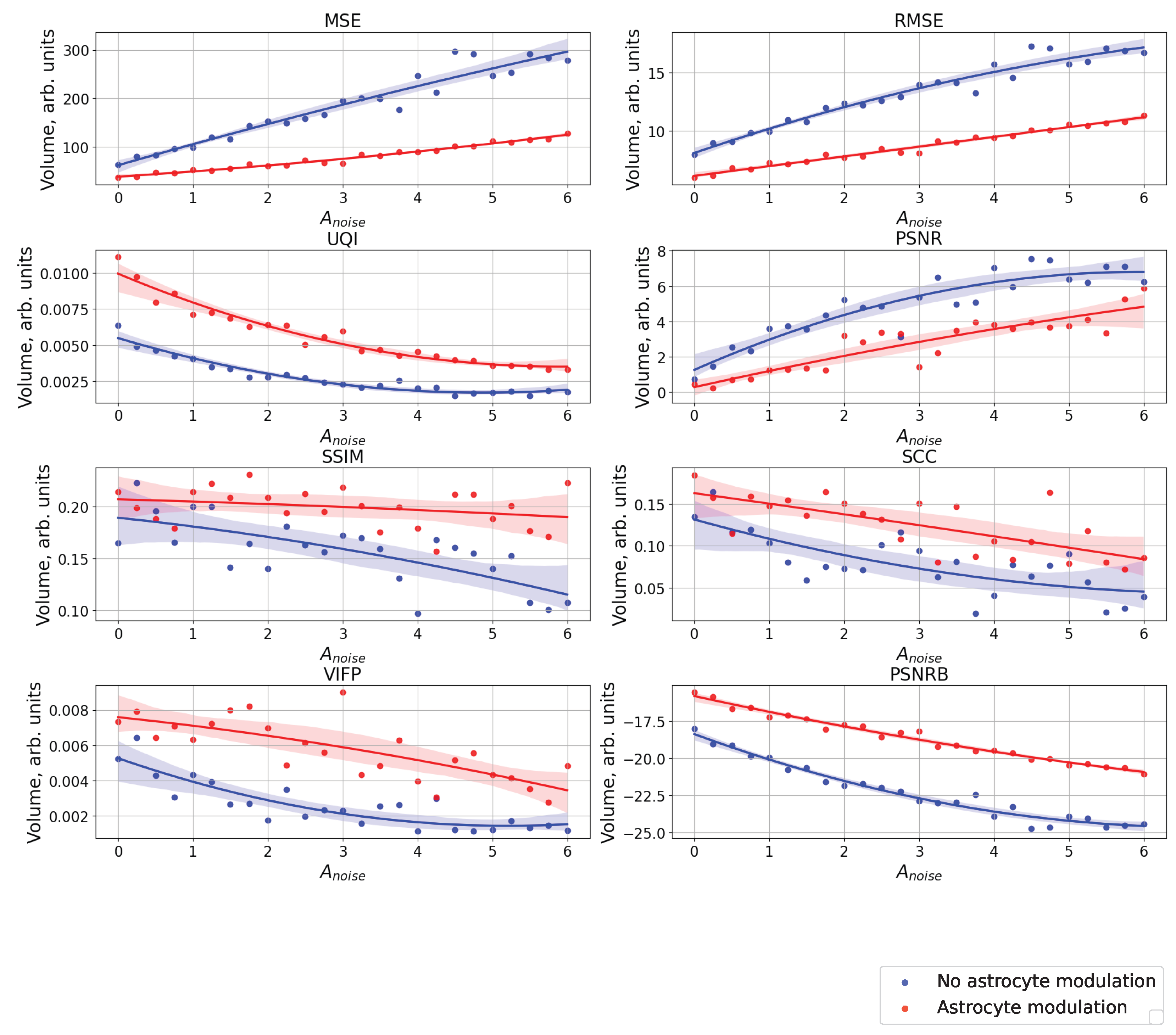
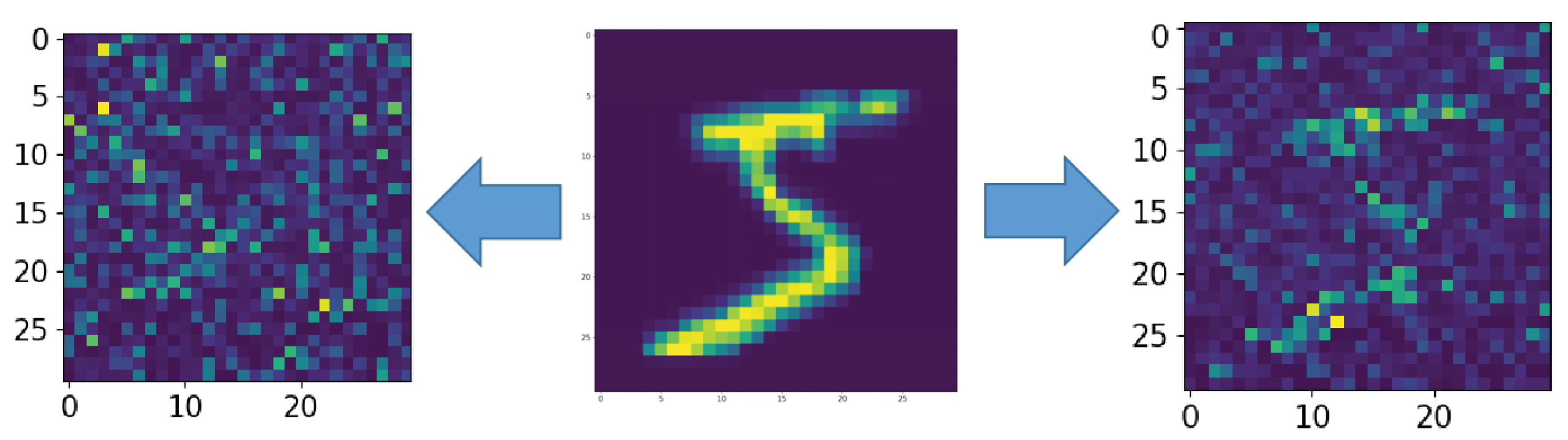
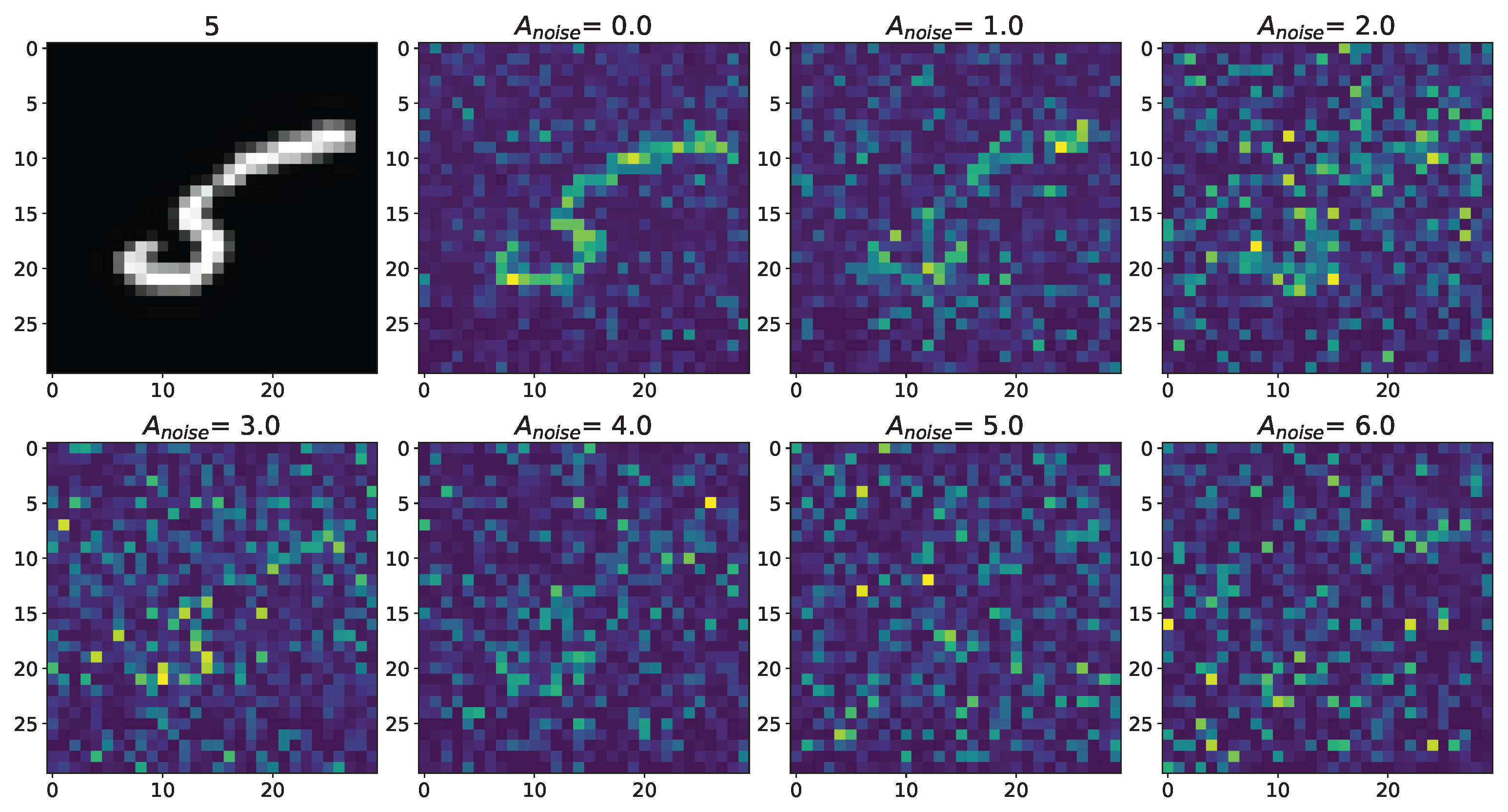
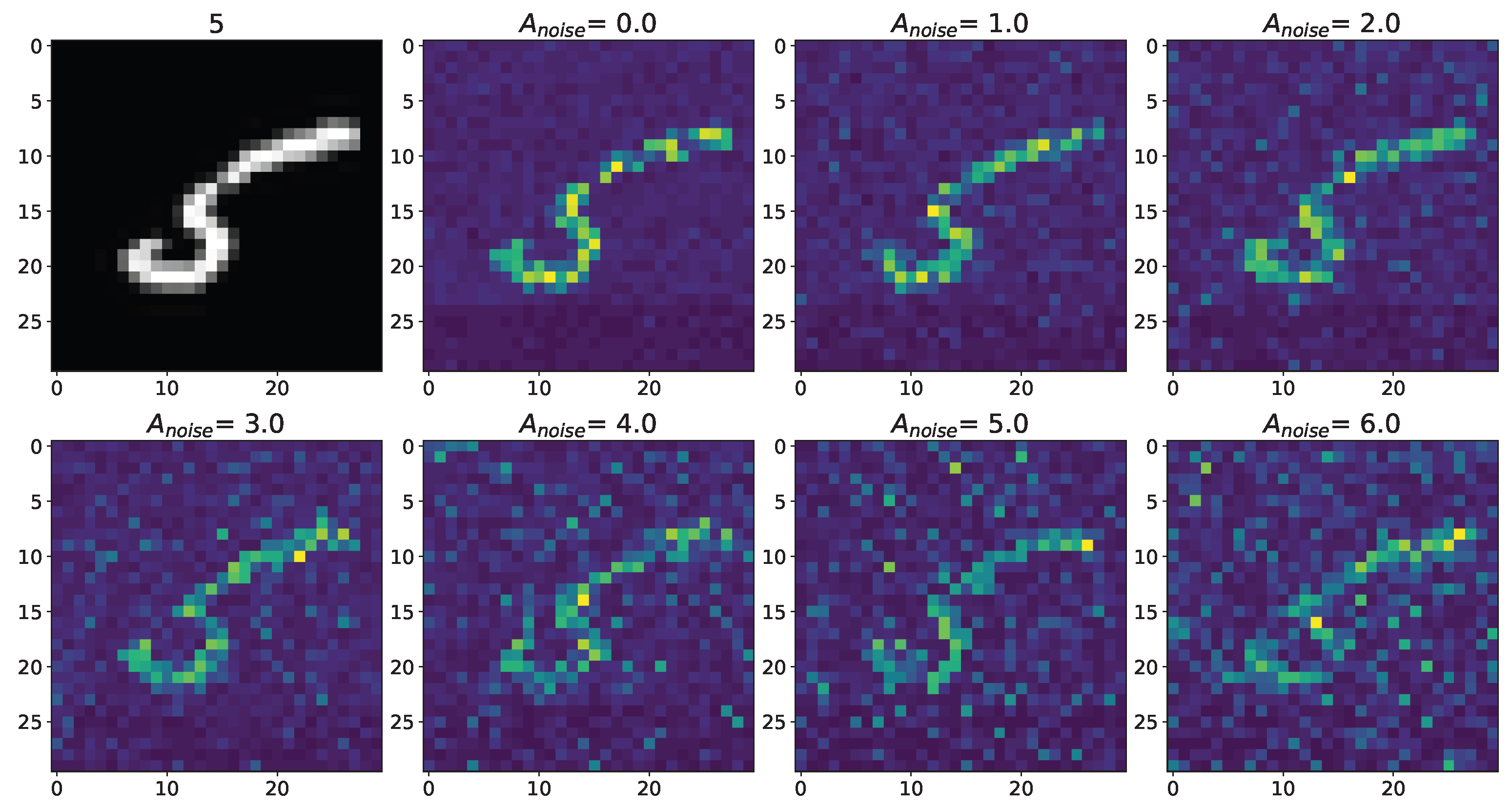
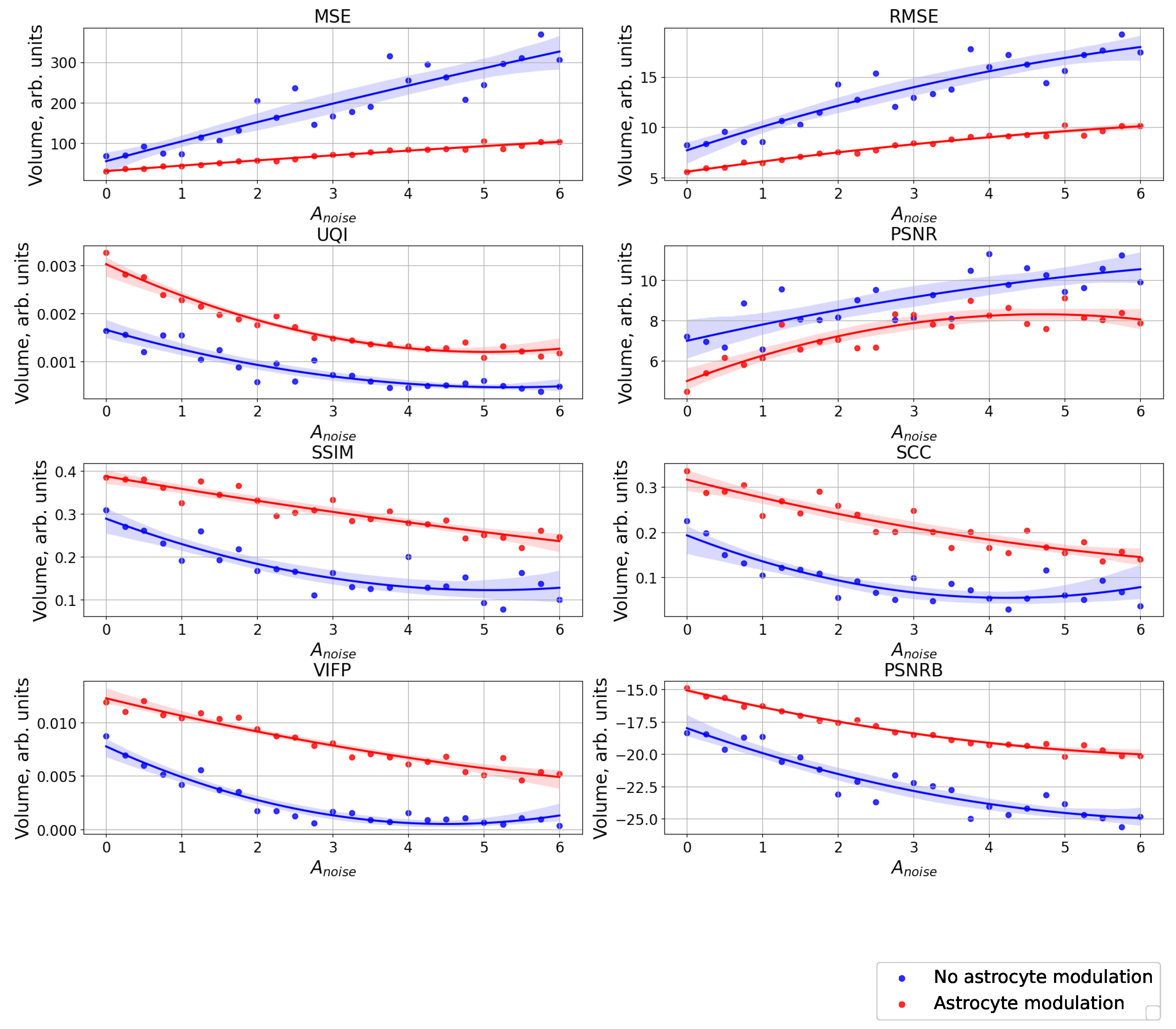
3. Results
4. Conclusions
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix A.1. Cases with Numbers from Database MNIST




Appendix A.2




References
- Araque, A.; Parpura, V.; Sanzgiri, R.; Haydon, P. Glutamate-dependent astrocyte modulation of synaptic transmission between cultured hippocampal neurons. Eur. J. Neurosci. 1998, 10, 2129–2142. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.; Haydon, P. Tripartite synapses: Glia, the unacknowledged partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Wittenberg, G.; Sullivan, M.; Tsien, J. Synaptic reentry reinforcement based network model for long-term memory consolidation. Hippocampus 2002, 12, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Wang, X. Synaptic basis of cortical persistent activity: The importance of NMDA receptors to working memory. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 9587–9603. [Google Scholar] [CrossRef] [Green Version]
- Haydon, P.G. GLIA: Listening and talking to the synapse. Nat. Rev. Neurosci. 2001, 2, 185–193. [Google Scholar] [CrossRef]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite synapses: Astrocytes process and control synaptic information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef]
- Nadkarni, S.; Jung, P. Dressed neurons: Modeling neural-glial interactions. Phys. Biol. 2004, 1, 35–41. [Google Scholar] [CrossRef]
- Nadkarni, S.; Jung, P. Modeling synaptic transmission of the tripartite synapse. Phys. Biol. 2007, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Volman, V.; Ben-Jacob, E.; Levine, H. The astrocyte as a gatekeeper of synaptic information transfer. Neural Comput. 2007, 326, 303–326. [Google Scholar] [CrossRef]
- De Pitta, M.; Volman, V.; Berry, H.; Ben-Jacob, E. A tale of two stories: Astrocyte regulation of synaptic depression and facilitation. PLoS Comput. Biol. 2011, 7, e1002293. [Google Scholar] [CrossRef]
- Postnov, D.; Ryazanova, L.; Sosnovtseva, O. Functional modeling of neural-glial interaction. Bio Syst. 2007, 89, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Amiri, M.; Bahrami, F.; Janahmadi, M. Functional contributions of astrocytes in synchronization of a neuronal network model. J. Theor. Biol. 2011, 292C, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Wade, J.; McDaid, L.; Harkin, J.; Crunelli, V.; Kelso, J. Bidirectional Coupling between Astrocytes and Neurons Mediates Learning and Dynamic Coordination in the Brain: A Multiple Modeling Approach. PLoS ONE 2011, 6, e29445. [Google Scholar] [CrossRef]
- Amiri, M.; Hosseinmardi, N.; Bahrami, F.; Janahmadi, M. Astrocyte-neuron interaction as a mechanism responsible for generation of neural synchrony: A study based on modeling and experiments. J. Comput. Neurosci. 2013, 34, 489–504. [Google Scholar] [CrossRef]
- Pankratova, E.; Kalyakulina, A.; Stasenko, S.; Gordleeva, S.; Lazarevich, I.; Kazantsev, V. Neuronal synchronization enhanced by neuron–astrocyte interaction. Nonlinear Dyn. 2019, 97, 647–662. [Google Scholar] [CrossRef]
- Ullah, G.; Jung, P.; Cornell-Bell, A. Anti-phase calcium oscillations in astrocytes via inositol (1, 4, 5)-trisphosphate regeneration. Cell Calcium 2006, 39, 197–208. [Google Scholar] [CrossRef]
- Kazantsev, V. Spontaneous calcium signals induced by gap junctions in a network model of astrocytes. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2009, 79, 010901. [Google Scholar] [CrossRef]
- Verisokin, A.; Verveyko, D.; Postnov, D.; Brazhe, A. Modeling of astrocyte networks: Toward realistic topology and dynamics. Front. Cell. Neurosci. 2021, 15, 645068. [Google Scholar] [CrossRef]
- Wallach, G.; Lallouette, J.; Herzog, N.; De Pitta, M.; Jacob, E.; Berry, H.; Hanein, Y. Glutamate mediated astrocytic filtering of neuronal activity. PLoS Comput. Biol. 2014, 10, e1003964. [Google Scholar] [CrossRef]
- Riera, J.; Hatanaka, R.; Ozaki, T.; Kawashima, R. Modeling the spontaneous Ca2+ oscillations in astrocytes: Inconsistencies and usefulness. J. Integr. Neurosci. 2011, 10, 439–473. [Google Scholar] [CrossRef]
- Manninen, T.; Havela, R.; Linne, M. Computational models for calcium-mediated astrocyte functions. Front. Comput. Neurosci. 2018, 12, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordleeva, S.; Tsybina, Y.; Krivonosov, M.; Ivanchenko, M.; Zaikin, A.; Kazantsev, V.; Gorban, A. Modeling working memory in a spiking neuron network accompanied by astrocytes. Front. Cell. Neurosci. 2021, 15, 631485. [Google Scholar] [CrossRef]
- Tsybina, Y.; Kastalskiy, I.; Krivonosov, M.; Zaikin, A.; Kazantsev, V.; Gorban, A.; Gordleeva, S. Astrocytes mediate analogous memory in a multi-layer neuron–astrocyte network. Neural Comput. Appl. 2022, 34, 9147–9160. [Google Scholar] [CrossRef]
- Gordleeva, S.; Tsybina, Y.; Krivonosov, M.; Tyukin, I.; Kazantsev, V.; Zaikin, A.; Gorban, A. Situation-based memory in spiking neuron-astrocyte network. arXiv 2022, arXiv:2202.07218. [Google Scholar]
- Hodgkin, A.; Huxley, A. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef]
- Izhikevich, E.M. Dynamical Systems in Neuroscience: The Geometry of Excitability and Bursting, 1st ed.; The MIT Press: Cambridge, MA, USA, 2007; p. 441. [Google Scholar]
- Angulo, M.; Kozlov, A.; Charpak, S.; Audinat, E. Glutamate released from glial cells synchronizes neuronal activity in the hippocampus. J. Neurosci. 2004, 24, 6920–6927. [Google Scholar] [CrossRef] [Green Version]
- Halassa, M.; Fellin, T.; Haydon, P. Tripartite synapses: Roles for astrocytic purines in the control of synaptic physiology and behavior. Neuropharmacology 2009, 57, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Gordleeva, S.; Stasenko, S.; Semyanov, A.; Dityatev, A.; Kazantsev, V. Bi-directional astrocytic regulation of neuronal activity within a network. Front. Comput. Neurosci. 2012, 6, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarevich, I.; Stasenko, S.; Kazantsev, V. Synaptic multistability and network synchronization induced by the neuron–glial interaction in the brain. JETP Lett. 2017, 105, 210–213. [Google Scholar] [CrossRef]
- Stasenko, S.; Lazarevich, I.; Kazantsev, V. Quasi-synchronous neuronal activity of the network induced by astrocytes. Procedia Comput. Sci. 2020, 169, 704–709. [Google Scholar] [CrossRef]
- Martín, E.; Fernández, M.; Perea, G.; Pascual, O.; Haydon, P.; Araque, A.; Ceña, V. Adenosine released by astrocytes contributes to hypoxia-induced modulation of synaptic transmission. Glia 2007, 55, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Jourdain, P.; Bergersen, L.; Bhaukaurally, K.; Bezzi, P.; Santello, M.; Domercq, M.; Matute, C.; Tonello, F.; Gundersen, V.; Volterra, A. Glutamate exocytosis from astrocytes controls synaptic strength. Nat. Neurosci. 2007, 10, 331–339. [Google Scholar] [CrossRef]
- Fiacco, T.; McCarthy, K. Intracellular astrocyte calcium waves in situ increase the frequency of spontaneous AMPA receptor currents in CA1 pyramidal neurons. J. Neurosci. 2004, 24, 722–732. [Google Scholar] [CrossRef] [Green Version]
- Rusakov, D.; Kullmann, D. Extrasynaptic glutamate diffusion in the hippocampus: Ultrastructural constraints, uptake, and receptor activation. J. Neurosci. 1998, 18, 3158–3170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rossum, G.; Drake, F., Jr. Python Tutorial; Centrum voor Wiskunde en Informatica Amsterdam: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Stimberg, M.; Brette, R.; Goodman, D. Brian 2, an intuitive and efficient neural simulator. Elife 2019, 8, e47314. [Google Scholar] [CrossRef]
- Hunter, J. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- LeCun, Y.; Cortes, C.; Burges, C. MNIST Handwritten Digit Database. 2010. Available online: http://yann.lecun.com/exdb/mnist (accessed on 1 November 2022).
- Xiao, H.; Rasul, K.; Vollgraf, R. Fashion-mnist: A novel image dataset for benchmarking machine learning algorithms. arXiv 2017, arXiv:1708.07747. [Google Scholar]
- Yang, R.; Deng, Y.; Zhu, A.; Tong, X.; Chen, Z. Few Shot Learning Based on the Street View House Numbers (SVHN) Dataset. In International Conference on Edge Computing and IoT; Springer: Cham, Switzerland, 2021; pp. 86–102. [Google Scholar]
- Sara, U.; Akter, M.; Uddin, M. Image quality assessment through FSIM, SSIM, MSE and PSNR—A comparative study. J. Comput. Commun. 2019, 7, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Bovik, A. A universal image quality index. IEEE Signal Process. Lett. 2002, 9, 81–84. [Google Scholar] [CrossRef]
- Manninen, T.; Saudargienė, A.; Linne, M. Astrocytic modulation of cortical synaptic plasticity: Integrating biological knowledge and computational modeling. In Glia: XV European Meeting on Glial Cells in Health and Disease Online-GLIA: Abstract Book: 5–9 July 2021; Wiley-Liss: New York, NY, USA, 2021; Volume 69, pp. E335–E4335. [Google Scholar]
- Wang, Y.; Fu, A.; Ip, N. Instructive roles of astrocytes in hippocampal synaptic plasticity: Neuronal activity-dependent regulatory mechanisms. FEBS J. 2022, 289, 2202–2218. [Google Scholar] [CrossRef]
- Noriega-Prieto, J.; Araque, A. Sensing and regulating synaptic activity by astrocytes at tripartite synapse. Neurochem. Res. 2021, 46, 2580–2585. [Google Scholar] [CrossRef]
- González-Arias, C.; Perea, G. Gliotransmission at tripartite synapses. In Computational Glioscience; Springer: Cham, Switzerland, 2019; pp. 213–226. [Google Scholar]
- Santello, M.; Toni, N.; Volterra, A. Astrocyte function from information processing to cognition and cognitive impairment. Nat. Neurosci. 2019, 22, 154–166. [Google Scholar] [CrossRef]
- Kol, A.; Adamsky, A.; Groysman, M.; Kreisel, T.; London, M.; Goshen, I. Astrocytes contribute to remote memory formation by modulating hippocampal–cortical communication during learning. Nat. Neurosci. 2020, 23, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Kastanenka, K.; Moreno-Bote, R.; De Pittà, M.; Perea, G.; Eraso-Pichot, A.; Masgrau, R.; Poskanzer, K.; Galea, E. A roadmap to integrate astrocytes into Systems Neuroscience. Glia 2020, 68, 5–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, J.; Araque, A. Astrocyte regulation of neural circuit activity and network states. Glia 2022, 70, 1455–1466. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stasenko, S.V.; Kazantsev, V.B. Dynamic Image Representation in a Spiking Neural Network Supplied by Astrocytes. Mathematics 2023, 11, 561. https://doi.org/10.3390/math11030561
Stasenko SV, Kazantsev VB. Dynamic Image Representation in a Spiking Neural Network Supplied by Astrocytes. Mathematics. 2023; 11(3):561. https://doi.org/10.3390/math11030561
Chicago/Turabian StyleStasenko, Sergey V., and Victor B. Kazantsev. 2023. "Dynamic Image Representation in a Spiking Neural Network Supplied by Astrocytes" Mathematics 11, no. 3: 561. https://doi.org/10.3390/math11030561
APA StyleStasenko, S. V., & Kazantsev, V. B. (2023). Dynamic Image Representation in a Spiking Neural Network Supplied by Astrocytes. Mathematics, 11(3), 561. https://doi.org/10.3390/math11030561