The analysis is organised as follows. First, we check the unboundedness of the prey and predator populations, we derive the dimensionless version of the equations, we compute the equilibrium points and we study their stability by applying linear stability analysis.
Section 3.1,
Section 3.2,
Section 3.3 and
Section 3.4 cover this first part of the study and give the analytical results for each of the models above. Finally, in
Section 3.5, we use standard numerical methods and the software Xppaut to give the one-parameter and two-parameter bifurcation diagrams.
3.1. Predator–Prey Dynamics with Specialist Predator and Herd-Linear Functional Response: Boundedness, Equilibrium Points and Stability Analysis
We study the dynamics of the model by Rosenzweig and MacArthur [
16] with herd-linear response
, conversion factor
e of captured prey into new predators, per capita natural mortality rate
m for the predators and logistic growth
for the prey, where
r denotes their net growth rate and
K their carrying capacity,
We show that the populations do not grow unbounded (we refer to the work by Bulai and Venturino in [
7]). We define with
the total population density and, summing up the equations for the prey and predator populations, we obtain
We collect
on the left-hand side and drop the term
to obtain
The value of
is at
and, by substituting this, we obtain
We solve the equation for
and get the upper bound for
, as well as
and
To reduce the number of parameters, we introduce the dimensionless quantities
,
,
,
,
. Applying the substitutions and dropping the tildes, we obtain the non-dimensional system
with
and
.
The equilibria follow by setting the equations in (12) and (13) to zero. We obtain the trivial equilibrium points of the system
and the interior equilibrium
with full expression below
Note that the interior equilibrium is positive if
.
We use the Jacobian matrix of the system in (12) and (13) to study the stability of the equilibria
The Jacobian evaluated at
has possibly a singularity, but the instability of this point can be assessed looking back at the original Equations (12) and (13). With
, and
x near 0, the first equation behaves like
, so that
x grows. Conversely, on
the second equation is
and
. Hence,
is a saddle. When evaluated at the equilibrium
, the determinant of the Jacobian matrix is
and is positive if
, that is, when the interior equilibrium is not feasible (i.e., does not exist or is negative). Under the same condition, the trace of the Jacobian at
,
, is negative and the equilibrium is stable.
For simplicity, we rewrite the Jacobian matrix evaluated at the interior equilibrium
in terms of the functions
and
The determinant of the matrix in (17) is
(since the functional response is an increasing function of the prey density), therefore the stability of the interior equilibrium depends on the sign of the trace
, and, in particular, on the slope of the prey zero-growth isocline
(see also Gause [
17] and Gause et al. [
18]). We obtain that the equilibrium
is stable if
(check
Table 1 for a summary of the feasibility and stability conditions).
We conclude that a transcritical bifurcation occurs at , where the interior equilibrium exchanges stability with the predator-free equilibrium. A Hopf bifurcation appears at , as the eigenvalues of the community matrix become purely imaginary and the system converges to a stable limit cycle.
3.2. Predator–Prey Dynamics with Generalist Predator and Herd-Linear Functional Response: Boundedness, Equilibrium Points and Stability Analysis
We study the dynamics of the model by Rosenzweig and MacArthur [
16] with herd-linear response
, conversion factor
e of captured prey into new predators and per capita natural mortality rate
m for the predators. We assume logistic growth for both the prey and the predator species,
and
, respectively, where
r is the net growth rate of the prey and
their carrying capacity, while
s denotes the predators’ reproduction rate, i.e., not discounted by deaths,
In this way, the predators are subjected to intraspecific competition, which occurs at rate
.
To check that the populations do not grow unbounded, we set
and, by repeating the steps in
Section 3.1, we get the differential equation for the total population
We differentiate the right-hand term with respect to
x and
y to get the local maximum
. By substitution in the equation above, we obtain
Therefore, the solution for the total population reads
where the upper bound is applicable also for
and
.
To obtain the non-dimensional version of the system in (18) and (19), we consider the dimensionless variables and parameters
,
,
,
,
,
,
. We drop the tildes and obtain the dimensionless system
with
,
and
.
We proceed with computing the equilibria. The trivial equilibria are
with
feasible if
. The interior equilibria are given by the intersection of the isoclines
and
Note that the isocline in (26) intersects the
x-axis at
and
and has a maximum at
for
, while the isocline in (27) intersects the vertical axis at
and is a root function translated by
and dilated by
. Therefore, if the intersection point of the isocline in (27) lies in the positive quadrant, i.e., if
, we find three different configurations for the phase plane: the two isoclines can intersect at most twice at
and
if
, or be tangent at
when
or never intersect in
when
. The equilibria are obtained as the positive roots of the curve
and the non-negativity of
y is ensured by the condition
When , the isocline in (27) intersects the vertical axes at and we find at most one interior equilibrium . We obtain the feasibility condition for by imposing that the curve in (27) takes positive values at , that is, if .
The Jacobian matrix of the system in (23) and (24) is given by
The equilibrium
, restricting the analysis to the trajectories on the coordinate axes, is seen to be either an unstable node if
, or a saddle if
. The prey-only equilibrium
is a stable node if
, otherwise the steady state is an unstable saddle. Under its feasibility condition
, the equilibrium
is always an unstable saddle. We summarise the feasibility and stability conditions studied above in
Table 2 and
Table 3.
We rewrite the Jacobian matrix evaluated at the interior equilibrium in terms of functions
,
and
:
The trace and the determinant at the interior equilibrium are given by
When only one interior equilibrium exists and is positive, the signs of the trace and the determinant determine its asymptotic stability, more specifically if
and
.
It seems rather difficult to obtain more analytical details on the stability of the equilibria and bifurcations for the model in (23) and (24). If possible, a more detailed analysis will be the topic of a future work.
3.3. Predator–Prey Dynamics with Specialist Predator and Herd-Holling Type II Functional Response: Boundedness, Equilibrium Points and Stability Analysis
We study the dynamics of the model by Rosenzweig and MacArthur [
16] with the herd-Holling type II functional response
derived in
Section 2, conversion factor
e of captured prey into new predators and per capita natural mortality rate
m for the predators, logistic growth
for the prey, where
r denotes their net growth rate and
K their carrying capacity,
The total population
verifies
with
as in
Section 3.1.
We obtain the dimensionless version of the model by applying the substitutions
,
,
,
,
,
. We drop the tildes and get the equations
with
and
.
We compute the equilibria by setting the equations in (37) and (38) to zero. The trivial equilibria are
and
, while the unique interior equilibrium
has full expression
and exists and is positive if and only if
.
The Jacobian matrix corresponding to the system in (37) and (38) is given by
The origin is unstable, being a saddle, a fact that is shown restricting the system to the coordinate axes, as previously done for the system (12) and (13). The equilibrium
is stable if and only if
(under this condition the determinant of the Jacobian matrix at the equilibrium is positive and the trace is negative). The prey-only equilibrium changes its stability at
when a transcritical bifurcation occurs. We can use the same formulation as in (17) for the Jacobian evaluated at the interior equilibrium, which for convenience we reproduce here
As the determinant of the Jacobian matrix is always positive, the stability of the interior equilibrium depends on the sign of the trace, in particular on the slope of the predator isocline,
. For the same reason as in
Section 3.1, the system in (37) and (38) undergoes a Hopf bifurcation when
and converges to a stable limit cycle for
, otherwise it converges to the interior equilibrium
. In
Table 4 we give a summary of the feasibility and stability conditions of the equilibria.
3.4. Predator–Prey Dynamics with Generalist Predator and Herd-Holling Type II Functional Response: Boundedness, Equilibrium Points and Stability Analysis
We study the dynamics of the model by Rosenzweig and MacArthur [
16] with the herd-Holling type II functional response
derived in
Section 2, conversion factor
e of captured prey into new predators and per capita natural mortality rate
m for the predators. We assume logistic growth for both the prey and the predator species,
and
, respectively, where
r is the net growth rate of the prey and
their carrying capacity, while
s denotes the predators’ reproduction rate,
Once again, note the second term in the predators’ equation, whose coefficient models predators intraspecific competition.
The boundedness of the populations is ensured by the condition on the total population density
with
as in
Section 3.1.
We use the dimensionless quantities
,
,
,
,
,
,
,
to obtain the dimensionless system of equations
with
,
and
.
The corresponding trivial equilibria correspond to the ones in
Section 3.2 and are given by
with
being feasible if
. The interior equilibria are given by the intersection of the isoclines
and
The isocline in (48) intersects the horizontal axis at
and
, while the isocline in (49) intersects the vertical axis at
. As for the model with generalist predator and linear functional response in
Section 3.2, we may expect that, under some conditions, the system admits two interior equilibria. However, given the formulation of the isoclines in (48) and (49), it seems difficult to find explicit analytical results and we refer to the next
Section 3.5 for more details on the interior equilibria and their feasibility and stability conditions.
We obtain the Jacobian matrix to check the stability of the trivial equilibria:
Again, restricting the analysis to the trajectories on the coordinate axes, we obtain that
is an unstable node for
, or a saddle if
. We find that the origin
is an unstable saddle, as well as the predator-only equilibrium
. The prey-only equilibrium
is a stable node if
, an unstable saddle otherwise. We give a summary of these results in
Table 5.
3.5. One-Parameter and Two-Parameter Bifurcation Diagrams
In this section, we proceed with the bifurcation analysis. We give the one-parameter bifurcation diagrams and vary either the value of the predator mortality rate, m, or the herding index, , when possible. Additionally, we obtain the two-parameter bifurcation diagrams with respect to the parameter pairs or , where 🟉 equals one of the other parameters in the model. Note that in the numerical simulations we use the model and parameters prior to non-dimensionalisation, to obtain a complete analysis with respect to all the model parameters.
We first study the predator–prey model with specialist predator and herd-linear functional response. In the one-parameter bifurcation plots, we fix the parameter values of the model in (6) and (7) as in
Table 6 and we call it the nominal set (hypothetical values).
In
Figure 1 we give the one-parameter bifurcation diagrams with respect to the natural mortality rate of the predators,
m. Note that when
, the system undergoes a supercritical Hopf bifurcation (HB) and a stable limit cycle appears. At
, a transcritical bifurcation occurs, where the coexistence equilibrium loses its stability and the predator-free equilibrium becomes stable.
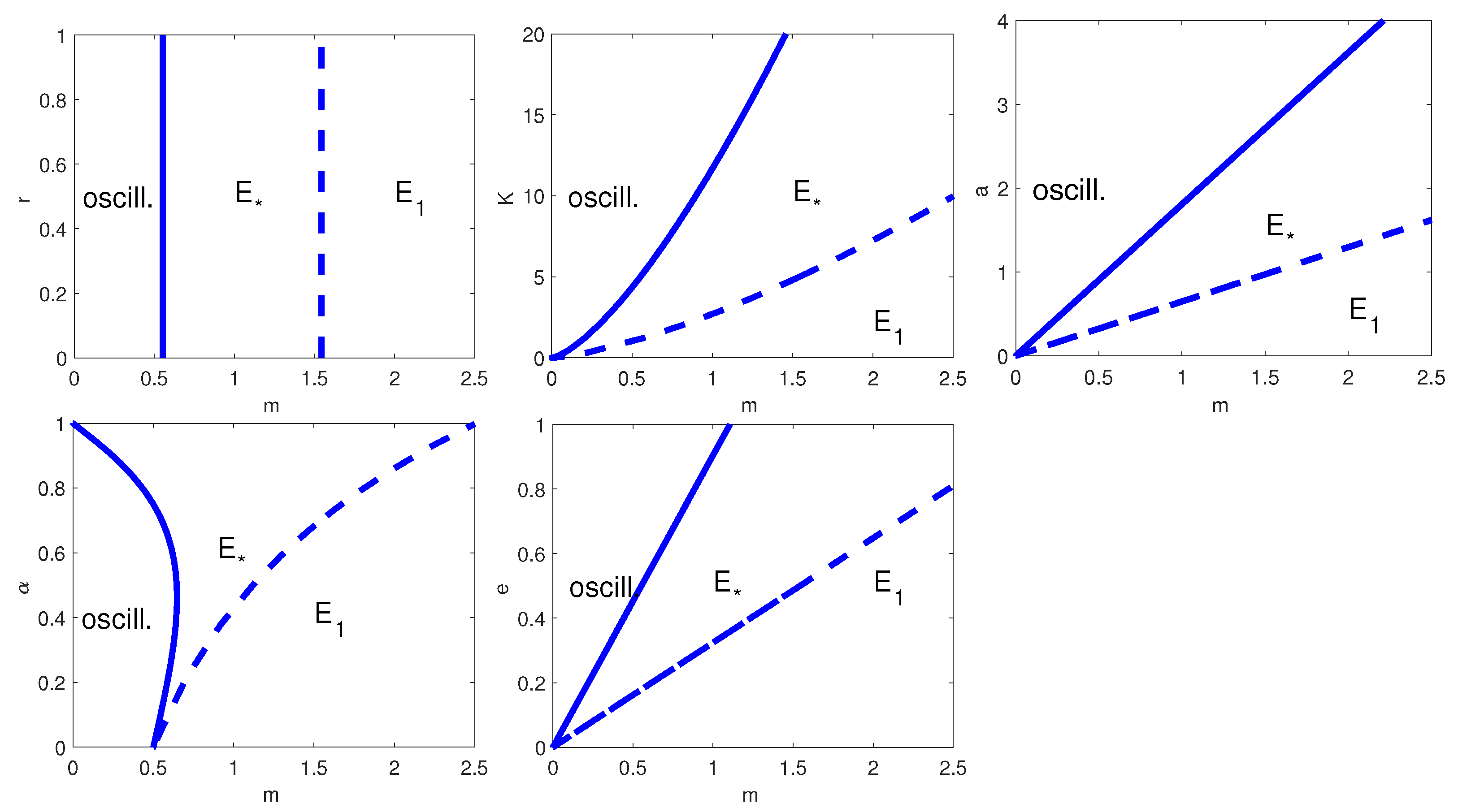
To complete the analysis, in
Figure 2 we have plotted all the possible two-parameter bifurcation diagrams for
, with
,
K,
a,
or
e. Note that the HB curve appears in all two-parameter bifurcation diagrams, but only in the plots where we vary
,
and
it occurs for every value of
m. When we let
r vary, the HB curve is present only at
(
Figure 2, top left); similarly, when we allow
to change, the HB occurs only for
(
Figure 2, bottom left).
As a second example, we analyse the predator–prey dynamics with generalist predator and herd-linear functional response. In
Table 7 we give the nominal set of parameter values for the model in (18) and (19).
In
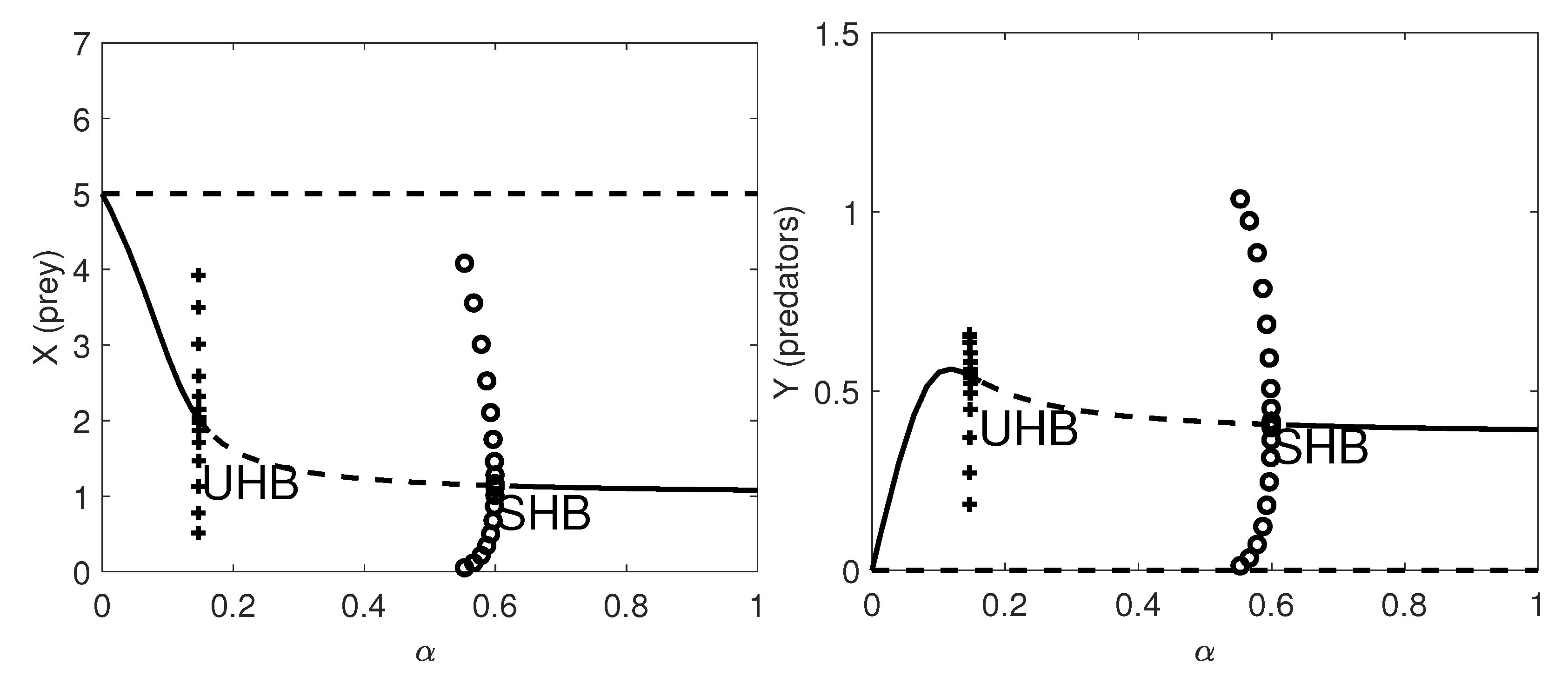
Figure 3 we give the one-parameter bifurcation diagram with respect to the herd exponent,
. Here a supercritical HB occurs at
and a subcritical HB appears at
.
The two-parameters bifurcation diagrams with respect to
, with
,
,
a,
s,
,
e, and
m for the model with Equations (18) and (19) are given in
Figure 4. When we vary the parameter pair
, a HB appears for all values of
independently of the value of
a, while for the remaining cases the HB is present only for some parameter values. Moreover, we observe that only when we vary the parameter pair
, the transcritical bifurcation curve is present. Finally, if one of the parameters
,
a,
s,
, and
e are below certain threshold values, with the other parameter values fixed as in
Figure 4, the coexistence equilibrium is asymptotically stable; analogously, when
r is above a certain threshold value the system converges to the interior equilibrium.
In this paragraph, we focus on the predator–prey dynamics with specialist predator and herd-Holling type II functional response. In
Table 8 we list the parameter values for the model in (34) and (35).
In
Figure 5 we obtain qualitatively similar results as for (6) and (7) for the one-parameter bifurcation diagrams with respect to the natural mortality rate of the predators,
m.
The dynamic described in
Figure 6 is similar to the one in
Figure 2. There are two main differences: in the two-parameter bifurcation diagrams with respect to
and
the HB curve is more concave; when we vary the parameter pair
(this case is not present in
Figure 2), one can see that the HB occurs for all values of
h and for
m smaller than a threshold value.
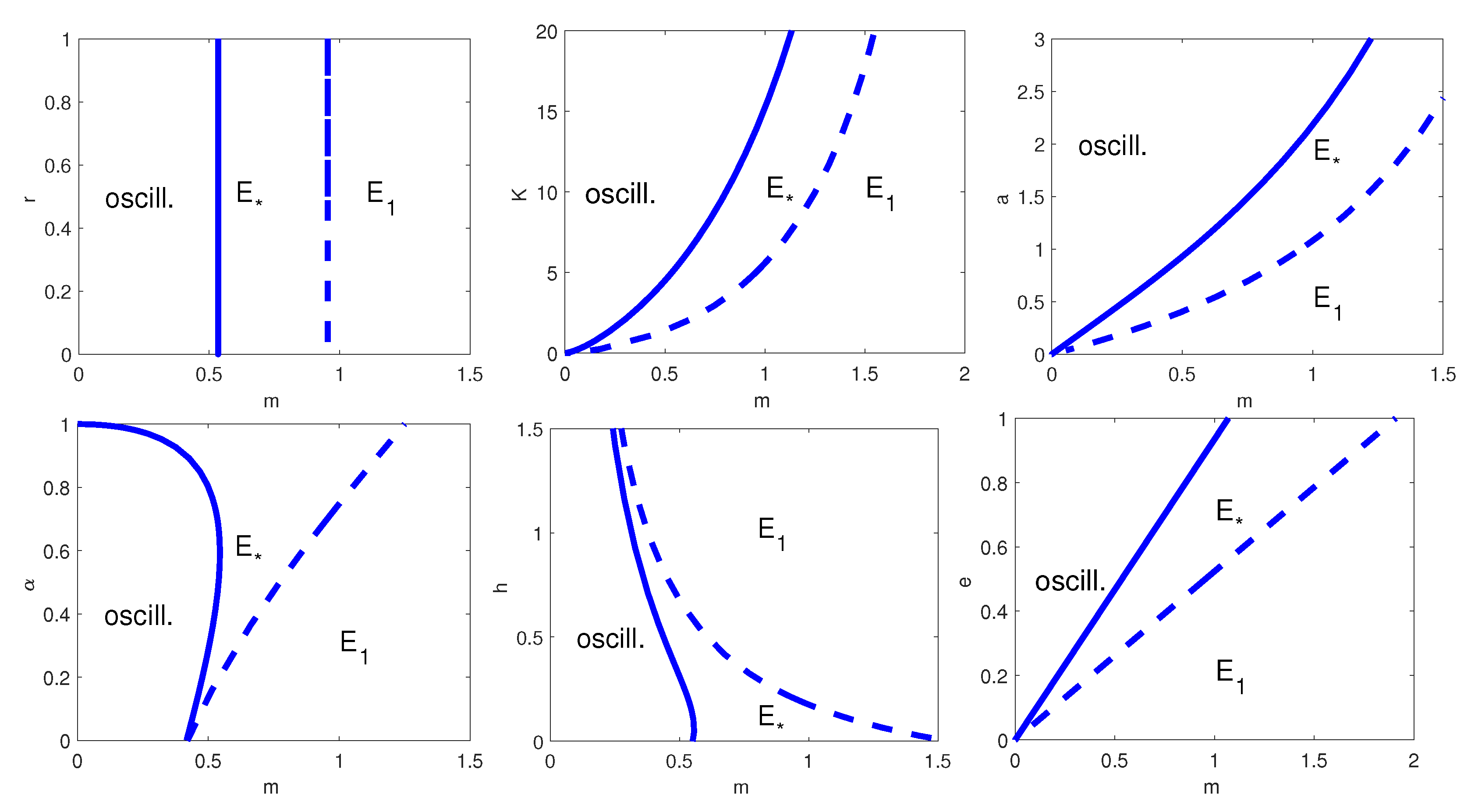
Finally, we study the predator–prey dynamics with generalist predator and herd-Holling type II functional response. In
Table 9 we list the nominal set of parameter values for model in (42) and (43). This is the most general model that we study which encompasses all the previously considered cases.
Both the one-parameter bifurcation diagram with respect to
m, and two-parameter bifurcation diagrams with respect to
, with
,
,
a,
,
h,
s,
and
e show behaviours similar to the previous models, see
Figure 7 and
Figure 8, respectively. It is worth noting that the results for models (34) and (35) are different as we give the two-parameter bifurcation diagrams with respect to the herd exponent as first parameter, while we refer to the predator natural mortality rate in the other two-parameter bifurcation plots.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}