From Gas Sensors to Biomimetic Artificial Noses


Abstract
:1. Introduction
2. Information from the Olfactory System
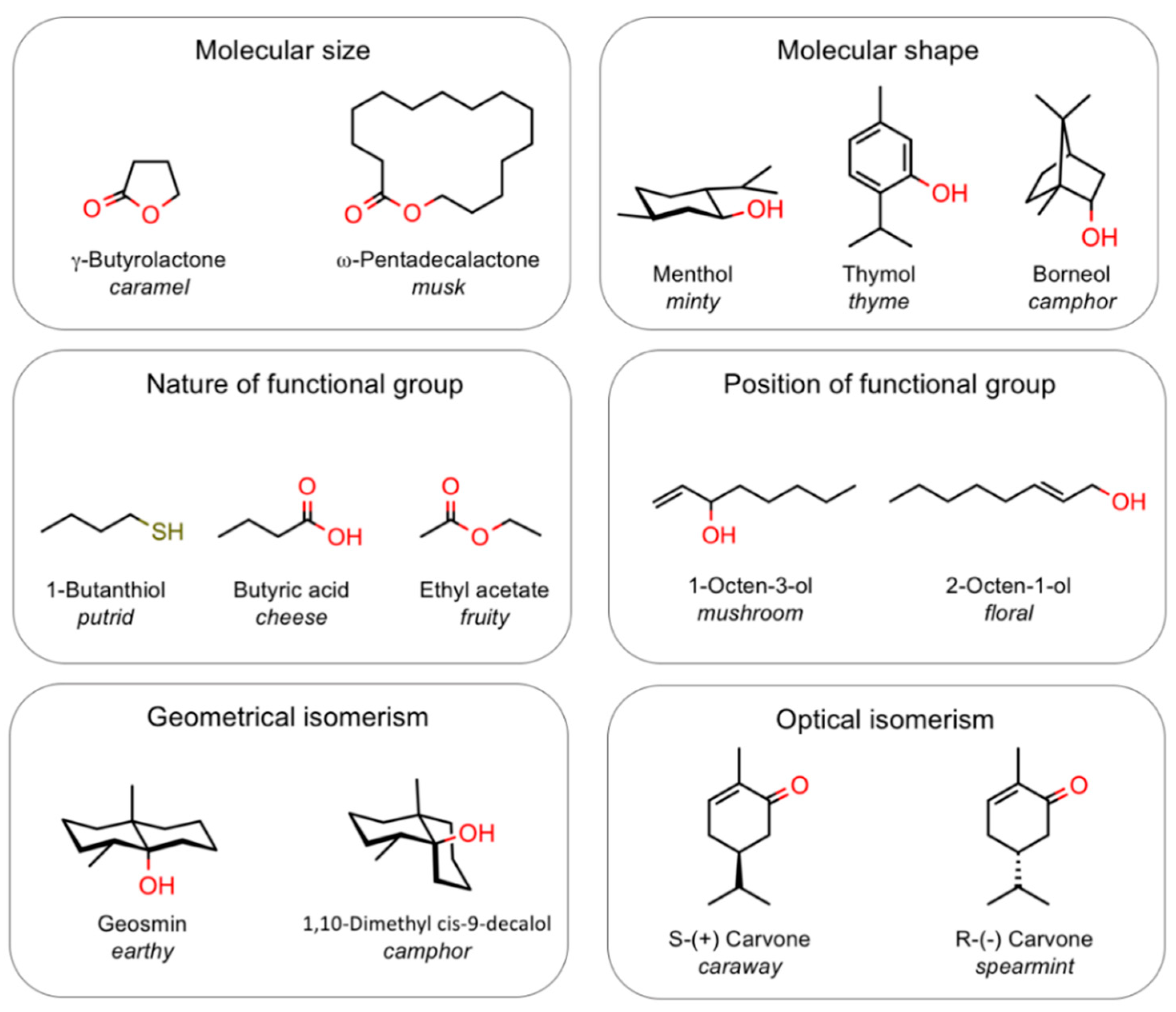
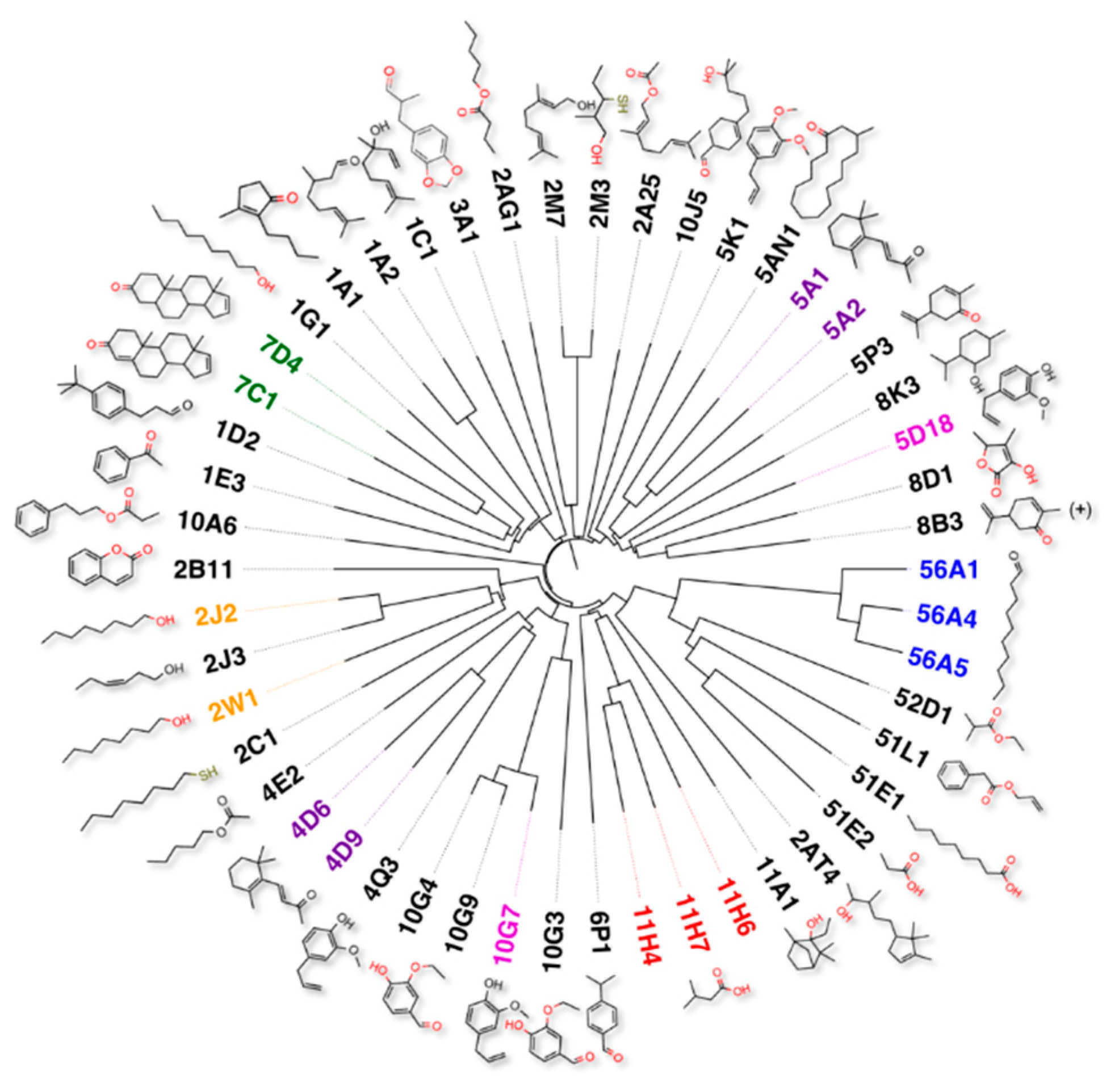
2.1. Complexity of the Olfactory Code
2.2. Knowledge of the Biological System
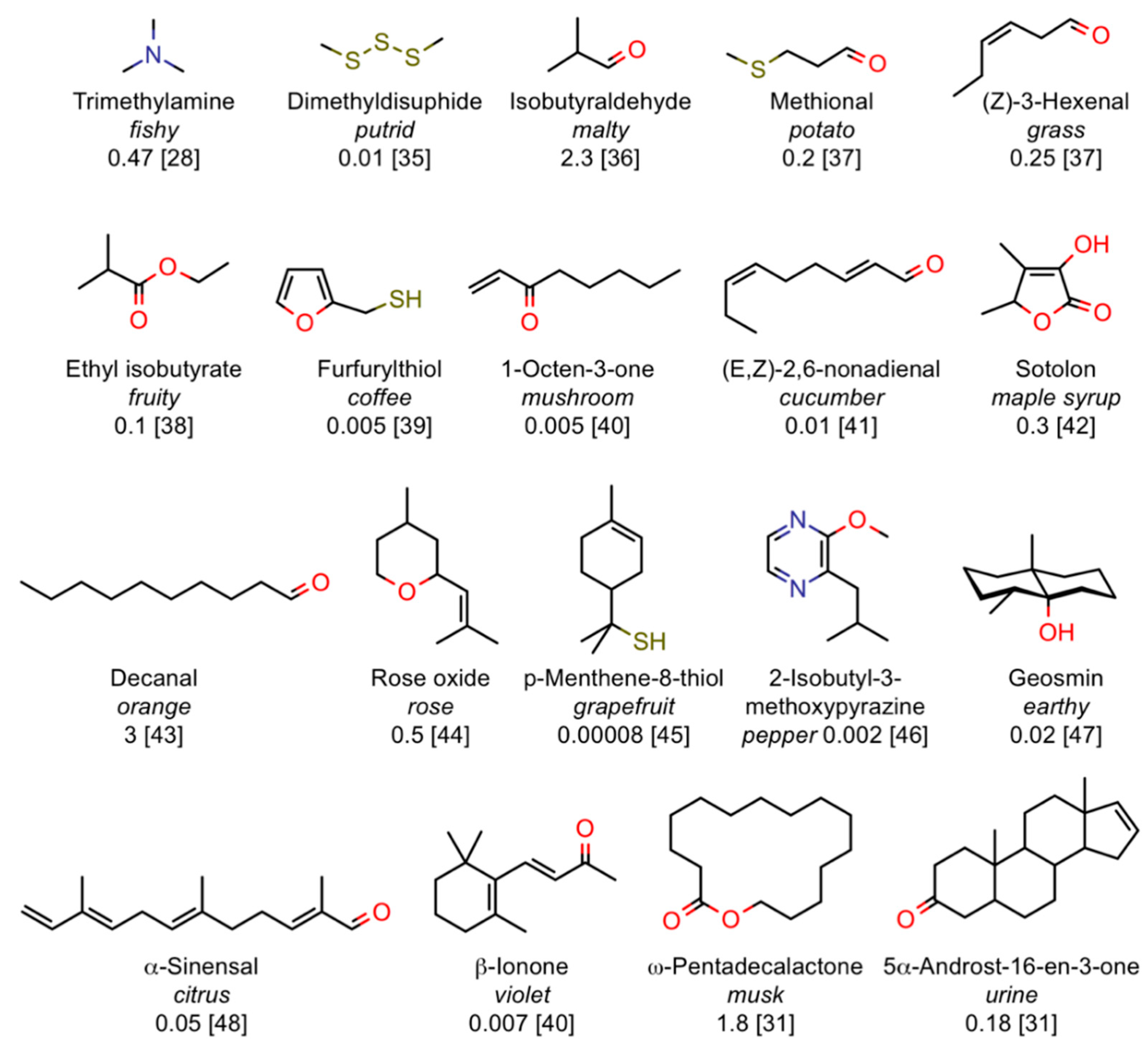
2.3. Sensitivity
3. An Artificial Nose
- An array of gas sensors, able to interact with volatile molecules and produce some sort of signal (electrical, optical, etc.) suitable to be amplified and processed.
- An amplifier to cope with the very low concentrations associated with odours.
- A pattern recognition software for recognizing specific response profiles associated with different odours.
3.1. Detecting the Signals
- Type of sensing elements. Obviously we can adopt sensors very different in their nature from olfactory receptors (they do not even have to be proteins), as far as they discriminate between different molecules on the same basis adopted by the biological systems; we know that, apart from few exceptions, stereochemical parameters are more important than functional groups, therefore our sensors should discriminate on the basis of size and shape; as an example, it is important that 1-octanol and 3-octanol, which smell floral and mushroom, respectively, should be clearly discriminated, while the latter should be rated as similar to 3-octanone, which also smells mushroom.
- Number of sensing elements. Mammals use several hundreds of different receptors, and humans in particular just over 300; are they really required, when with only three optical sensors we can discriminate millions of colours? The answer lies in the different performances of colour vision and olfaction. While with our nose we can pick-up single specific olfactory notes in a complex odour (just think of how we can detect with our nose the ingredients present in foods by inhaling their volatiles without separating the components), the three colours mix in our brain and we only perceive the final result. Such feature of olfaction is very important when it comes to smell some off-odour in our food among a variety of different good flavours, but is much more important in other animals, whose life depends on a correct functioning of their sense of smell: the gazelle can detect the smell of the lion, and vice versa, among a large number of environmental odours; likewise, an insect can recognise the sex pheromone released by the female of its own species among dozen of similar odours produced by other insects. On the other hand, the auditory system is equipped with thousands of sensors, each tuned to a very narrow part of the spectrum, thus enabling us to understand a conversation against an intense background of different sounds.
- The question of the code. Physiological noses detect and discriminate between a very large number of odorants using a combinatorial approach, which is based on a certain code. Just like the letters of the alphabet, used to form thousands of different words, elementary signals generated by each olfactory receptor combine into a complex odour picture. Now, the question is whether we are bound to use the same code as the natural one when designing an electronic nose. The answer is complex and depends on what sort of an instrument we aim to achieve. For a faithful of reproduction of the human nose, probably we have to use chemical sensors with more or less the selectivity of the natural olfactory receptors. In other words, we can better use the same code, but not necessarily. Our olfactory receptors are rather broadly tuned and cover with their sensitivity most of the spectrum of chemical structures within certain size limits: therefore, we could also adopt sensing elements whose individual response spectra are different from those of the natural receptors, provided that the all set of sensors covers the same area of chemical structures detected by the human nose. To make a simple analogy with colour vision, an instrument for measuring the colour could be made with three sensors not necessarily tuned to the same wavelength of our three rhodopsins, as long as the set of the three sensors covers all the interval from 400 to 800 nm.
3.2. The Question of the Environment
4. Types and Performance of Gas Sensors
4.1. Metal Oxides
4.2. Conducting Polymers
4.3. Olfactory Receptors
4.4. Soluble Binding Proteins
4.5. Transducing Strategies
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Pelosi, P.; Pisanelli, A.M.; Baldaccini, N.E.; Gagliardo, A. Binding of [3H]-2-isobutyl-3-methoxypyrazine to cow olfactory mucosa. Chem. Senses 1981, 6, 77–85. [Google Scholar] [CrossRef]
- Pelosi, P.; Baldaccini, N.E.; Pisanelli, A.M. Identification of a specific olfactory receptor for 2-isobutyl-3-methoxypyrazine. Biochem. J. 1982, 201, 245–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, R.G.; Riddiford, L.M. Pheromone binding and inactivation by moth antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Persaud, K.; Dodd, G. Analysis of discrimination mechanisms in the mammalian olfactory system using a model nose. Nature 1982, 299, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Persaud, K.C.; Marco, S.; Gutiérrez-Gálvez, A. Neuromorphic Olfaction; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2013. [Google Scholar]
- Stitzel, S.E.; Aernecke, M.J.; Walt, D.R. Artificial noses. Annu. Rev. Biomed. Eng. 2011, 13, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Wasilewski, T.; Gębicki, J.; Kamysz, W. Bioelectronic nose: Current status and perspectives. Biosens. Bioelectron. 2017, 87, 480–494. [Google Scholar] [CrossRef] [PubMed]
- Capelli, L.; Sironi, S.; Del Rosso, R. Electronic noses for environmental monitoring applications. Sensors 2014, 14, 19979–20007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikov, A.; Lázár, Z.; Horvath, I. Established methodological issues in electronic nose research: How far are we from using these instruments in clinical settings of breath analysis? J. Breath Res. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, H.; Chen, Q.; Mei, C.; Liu, G. Recent advances in electronic nose techniques for monitoring of fermentation process. World J. Microbiol. Biotechnol. 2015, 31, 1845–1852. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Méndez, M.L.; De Saja, J.A.; González-Antón, R.; García-Hernández, C.; Medina-Plaza, C.; García-Cabezón, C.; Martín-Pedrosa, F. Electronic Noses and Tongues in Wine Industry. Front. Bioeng. Biotechnol. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Wojnowski, W.; Majchrzak, T.; Dymerski, T.; Gębicki, J.; Namieśnik, J. Electronic noses: Powerful tools in meat quality assessment. Meat Sci. 2017, 131, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Majchrzak, T.; Wojnowski, W.; Dymerski, T.; Gębicki, J.; Namieśnik, J. Electronic noses in classification and quality control of edible oils: A review. Food Chem. 2018, 246, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Rossiter, K.J. Structure-Odor Relationships. Chem. Rev. 1996, 96, 3201–3240. [Google Scholar] [CrossRef] [PubMed]
- Sell, C.S. On the unpredictability of odor. Angew. Chem. Int. Ed. Engl. 2006, 45, 6254–6261. [Google Scholar] [CrossRef] [PubMed]
- Mombaerts, P. Molecular biology of odorant receptors in vertebrates. Annu. Rev. Neurosci. 1999, 22, 487–509. [Google Scholar] [CrossRef] [PubMed]
- Mombaerts, P. Odorant receptor genes in humans. Curr. Opin. Genet. Dev. 1999, 9, 315–320. [Google Scholar] [CrossRef]
- Mombaerts, P. The human repertoire of odorant receptor genes and pseudogenes. Annu. Rev. Genom. Hum. Genet. 2001, 2, 493–510. [Google Scholar] [CrossRef] [PubMed]
- Crasto, C.; Singer, M.S.; Shepherd, G.M. The olfactory receptor family album. Genome Biol. 2001, 2. [Google Scholar] [CrossRef]
- Olender, T.; Nativ, N.; Lancet, D. HORDE: Comprehensive resource for olfactory receptor genomics. Methods Mol. Biol. 2013, 1003, 23–38. [Google Scholar] [PubMed]
- Malnic, B.; Hirono, J.; Sato, T.; Buck, L.B. Combinatorial receptor codes for odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef]
- Korsching, S. Olfactory maps and odor images. Curr. Opin. Neurobiol. 2002, 12, 387–392. [Google Scholar] [CrossRef]
- Imai, T.; Sakano, H.; Vosshall, L.B. Topographic mapping—The olfactory system. Cold Spring Harb. Perspect. Biol. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Murthy, V.N. Olfactory maps in the brain. Annu. Rev. Neurosci. 2011, 34, 233–258. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Sakano, H. How is the olfactory map formed and interpreted in the mammalian brain? Annu. Rev. Neurosci. 2011, 34, 467–499. [Google Scholar] [CrossRef] [PubMed]
- Amoore, J.E. Specific anosmia: A clue to the olfactory code. Nature 1967, 214, 1095–1098. [Google Scholar] [CrossRef] [PubMed]
- Amoore, J.E. A Plan to Identify Most of the Primary Odors. In Olfaction and Taste III; Pfaffmann, C., Ed.; Rockefeller University Press: New York, NY, USA, 1969; pp. 158–171. [Google Scholar]
- Amoore, J.E. Specific anosmia and the concept of primary odors. Chem. Senses 1977, 2, 267–281. [Google Scholar] [CrossRef]
- Guillot, M. Anosmies Partielles et Odeurs Fondamentales. CRH Acad. Sci. 1948, 226, 1307–1309. [Google Scholar]
- Le Magnen, J. Les Phenomenes Olfacto-sexuels chez l’Homme. Arch. Sci. Physiol. 1952, 6, 125–160. [Google Scholar]
- Amoore, J.E.; Pelosi, P.; Forrester, L.J. Specific anosmia to 5α-androst-16-en-3-one and ω pentadecalactone: The urinous and musky primary odors. Chem. Senses 1977, 2, 401–425. [Google Scholar] [CrossRef]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Olender, T.; Keydar, I.; Pinto, J.M.; Tatarskyy, P.; Alkelai, A.; Chien, M.S.; Fishilevich, S.; Restrepo, D.; Matsunami, H.; Gilad, Y.; et al. The human olfactory transcriptome. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Verbeurgt, C.; Wilkin, F.; Tarabichi, M.; Gregoire, F.; Dumont, J.E.; Chatelain, P. Profiling of olfactory receptor gene expression in whole human olfactory mucosa. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Buttery, R.G.; Guadagni, D.G.; Ling, L.C.; Seifert, R.M.; Lipton, W. Additional volatile components of cabbage, broccoli, and cauliflower. J. Agric. Food Chem. 1976, 24, 829–832. [Google Scholar] [CrossRef]
- Amoore, J.E.; Firrester, L.J.; Pelosi, P. Specific anosmia to isobutyladehyde: The malty primary odor. Chem. Senses 1976, 2, 17–25. [Google Scholar]
- Buttery, R.G.; Teranishi, R.; Ling, L.C.; Turnbaugh, J.G. Quantitative and sensory studies on tomato paste volatiles. J. Agric. Food Chem. 1990, 38, 336–340. [Google Scholar] [CrossRef]
- Buttery, R.G.; Seifert, R.M.; Ling, L.C.; Soderstrom, E.L.; Ogawa, J.M.; Turnbaugh, J.G. Additional aroma components of honeydew melon. J. Agric. Food Chem. 1982, 30, 1208–1211. [Google Scholar] [CrossRef]
- Tressl, R. Formation of Flavor Components in Roasted Coffee. In Thermal Generation of Aromas; Parliment, T.H., McGorrin, R.J., Ho, C.-T., Eds.; American Chemical Society (ACS): Washington, DC, USA, 1989; pp. 285–301. [Google Scholar]
- Buttery, R.G.; Teranishi, R.; Flath, R.A.; Ling, L.C. Identification of additional tomato paste volatiles. J. Agric. Food Chem. 1990, 38, 792–795. [Google Scholar] [CrossRef]
- Teranishi, R.; Buttery, R.G.; Gaudagni, D.G. Odor Quality and Chemical Structure in Fruit and Vegetable Flavors. Ann. N. Y. Acad. Sci. 1974, 237, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Blank, I.; Lin, J.; Fumeaux, R.; Welti, D.H.; Fay, L.B. Formation of 3-hydroxy-4,5-dimethyl-2(5H)-furanone (sotolone) from 4-hydroxy-L-isoleucine and 3-amino-4,5-dimethyl-3,4-dihydro-2(5H)-furanone. J. Agric. Food Chem. 1996, 44, 1851–1856. [Google Scholar] [CrossRef]
- Ahmed, E.M.; Dennison, R.A.; Dougherty, R.H.; Shaw, P.E. Flavor and odor thresholds in water of selected orange juice components. J. Agric. Food Chem. 1978, 26, 187–191. [Google Scholar] [CrossRef]
- Plotto, A.; Margaria, C.A.; Goodner, K.L.; Goodrich, R.; Baldwin, E.A. Odour and flavor thresholds for key aroma components in an orange juice matrix: Ester and miscellaneous compounds. Flavour Fragr. J. 2008, 23, 398–406. [Google Scholar] [CrossRef]
- Demole, E.; Enggist, P.; Ohloff, G. p-Menthene-8-thiol: A powerful flavor impact constituent of grapefruit juice. Helv. Chim. Acta 1982, 65, 1785–1794. [Google Scholar] [CrossRef]
- Buttery, R.G.; Seifert, R.M.; Guadagni, D.G.; Ling, L.C. Characterization of some volatile constituents of bell peppers. J. Agric. Food Chem. 1969, 17, 1322–1327. [Google Scholar] [CrossRef] [PubMed]
- Buttery, R.G.; Guadagni, D.G.; Ling, L.C. Geosmin, a musty off-flavor of dry beans. J. Agric. Food Chem. 1976, 24, 419–420. [Google Scholar] [CrossRef]
- Ohloff, G. Recent Developments in the Field of Naturally Occurring Aroma Components. Prog. Chem. Org. Nat. Prod. 1978, 35, 431–527. [Google Scholar]
- Kaissling, K.E.; Priesner, E. Die Riechschwelle des Seidenspinners. Naturwissenschaften 1970, 57, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Kaissling, K.E. Chemo-electrical transduction in insect olfactory receptors. Annu. Rev. Neurosci. 1986, 9, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Kaissling, K.E. Pheromone Reception in Insects: The Example of Silk Moths. In Neurobiology of Chemical Communication; Mucignat-Caretta, C., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2014; Chapter 4. [Google Scholar]
- Firestein, S. How the olfactory system makes sense of scents. Nature 2001, 413, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Ronnett, G.V.; Moon, C. G proteins and olfactory signal transduction. Annu. Rev. Physiol. 2002, 64, 189–222. [Google Scholar] [CrossRef] [PubMed]
- Maresh, A.; Gil, D.R.; Whitman, M.C.; Greer, C.A. Principles of glomerular organization in the human olfactory bulb-implications for odor processing. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- Gladysheva, O.; Kukushkina, D.; Martynova, G. Glycoprotein composition of olfactory mucus in vertebrates. Acta Histochem. 1986, 78, 141–146. [Google Scholar] [CrossRef]
- Schleicher, S.; Boekhoff, I.; Arriza, J.; Lefkowitz, R.J.; Breer, H. A beta-adrenergic receptor kinase-like enzyme is involved in olfactory signal termination. Proc. Natl. Acad. Sci. USA 1993, 90, 1420–1424. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhao, A.Z.; Chan, G.C.; Baker, L.P.; Impey, S.; Beavo, J.A.; Storm, D.R. Phosphorylation and inhibition of olfactory adenylyl cyclase by CaM kinase II in Neurons: A mechanism for attenuation of olfactory signals. Neuron 1998, 21, 495–504. [Google Scholar] [CrossRef]
- Joshi, N.; Hayasaka, T.; Liu, Y.; Liu, H.; Oliveira, O.N., Jr.; Lin, L. A review on chemiresistive room temperature gas sensors based on metal oxide nanostructures, graphene and 2D transition metal dichalcogenides. Mikrochim. Acta 2018, 185. [Google Scholar] [CrossRef] [PubMed]
- Persaud, K.C.; Pelosi, P. An approach to an artifiacial nose. Trans. Am. Soc. Artif. Intern. Organs 1985, 31, 297–300. [Google Scholar] [PubMed]
- Amrani, M.E.H.; Persaud, K.C.; Payne, P.A. High-frequency measurements of conducting polymers: Development of a new technique for sensing volatile chemicals. Meas. Sci. Technol. 1995, 6, 1500–1507. [Google Scholar] [CrossRef]
- Hatfield, J.V.; Neaves, P.; Hicks, P.J.; Persaud, K.; Travers, P. Towards an integrated electronic nose using conducting polymer sensors. Sens. Actuators B Chem. 1994, 18, 221–228. [Google Scholar] [CrossRef]
- Lange, U.; Roznyatovskaya, N.V.; Mirsky, V.M. Conducting polymers in chemical sensors and arrays. Anal. Chim. Acta 2008, 614, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Persaud, K.C. Gas Sensors: Towards an artificial nose. In Sensors and Sensory Systems for Advanced Robots; Dario, P., Ed.; Springer: Berlin, Germany, 1988; pp. 361–382. [Google Scholar]
- Pelosi, P. The role of perireceptor events in vertebrate olfaction. Cell. Mol. Life Sci. 2001, 58, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Missbach, C.; Dweck, H.K.; Vogel, H.; Vilcinskas, A.; Stensmyr, M.C.; Hansson, B.S.; Grosse-Wilde, E. Evolution of insect olfactory receptors. eLife 2014, 26. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.B. Olfactory receptors and odor coding in mammals. Nutr. Rev. 2004, 62, 184–188. [Google Scholar] [CrossRef]
- Touhara, K.; Vosshall, L.B. Sensing odorants and pheromones with chemosensory receptors. Annu. Rev. Physiol. 2009, 71, 307–332. [Google Scholar] [CrossRef] [PubMed]
- Wasilewskia, T.; Gebickib, J.; Kamysza, W. Advances in olfaction-inspired biomaterials applied to bioelectronic noses. Sens. Actuator B Chem. 2018, 257, 511–537. [Google Scholar] [CrossRef]
- Breer, H. Olfactory receptors: Molecular basis for recognition and discrimination of odors. Anal. Bioanal. Chem. 2003, 377, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Latek, D.; Modzelewska, A.; Trzaskowski, B.; Palczewski, K.; Filipek, S. G protein-coupled receptors-recent advances. Acta Biochim. Pol. 2012, 59, 515–529. [Google Scholar] [PubMed]
- Bianchet, M.A.; Bains, G.; Pelosi, P.; Pevsner, J.; Snyder, S.H.; Monaco, H.L.; Amzel, L.M. The three dimensional structure of bovine odorant-binding protein and its mechanism of odor recognition. Nat. Struct. Biol. 1996, 3, 934–939. [Google Scholar] [CrossRef] [PubMed]
- Tegoni, M.; Ramoni, R.; Bignetti, E.; Spinelli, S.; Cambillau, C. Domain swapping creates a third putative combining site in bovine odorant binding protein dimer. Nat. Struct. Biol. 1996, 3, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Tegoni, M.; Pelosi, P.; Vincent, F.; Spinelli, S.; Campanacci, V.; Grolli, S.; Ramoni, R.; Cambillau, C. Mammalian odorant binding proteins. Biochim. Biophys. Acta 2000, 1482, 229–240. [Google Scholar] [CrossRef]
- Sandler, B.H.; Nikonova, L.; Leal, W.S.; Clardy, J. Sexual attraction in the silkworm moth: Structure of the pheromone-binding-protein-bombykol complex. Chem. Biol. 2000, 7, 143–151. [Google Scholar] [CrossRef]
- Tegoni, M.; Campanacci, V.; Cambillau, C. Structural aspects of sexual attraction and chemical communication in insects. Trends Biochem. Sci. 2004, 29, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Mulla, M.Y.; Tuccori, E.; Magliulo, M.; Lattanzi, G.; Palazzo, G.; Persaud, K.; Torsi, L. Capacitance-modulated transistor detects odorant binding protein chiral interactions. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Arena, S.; Spinelli, S.; Liu, D.; Zhang, G.; Wei, R.; Cambillau, C.; Scaloni, A.; Wang, G.; Pelosi, P. Reverse chemical ecology: Olfactory proteins from the giant panda and their interactions with putative pheromones and bamboo volatiles. Proc. Natl. Acad. Sci. USA 2017, 114, 9802–9810. [Google Scholar] [CrossRef] [PubMed]
- Paolini, S.; Tanfani, F.; Fini, C.; Bertoli, E.; Pelosi, P. Porcine odorant-binding protein: Structural stability and ligand affinities measured by fourier-transform infrared spectroscopy and fluorescence spectroscopy. Biochim. Biophys. Acta 1999, 1431, 179–188. [Google Scholar] [CrossRef]
- Schwaighofer, A.; Pechlaner, M.; Oostenbrink, C.; Kotlowski, C.; Araman, C.; Mastrogiacomo, R.; Pelosi, P.; Knoll, W.; Nowak, C.; Larisika, M. Insights into structural features determining odorant affinities to honey bee odorant binding protein 14. Biochem. Biophys. Res. Commun. 2014, 446, 1042–1046. [Google Scholar] [CrossRef] [PubMed]
- Schwaighofer, A.; Kotlowski, C.; Araman, C.; Chu, N.; Mastrogiacomo, R.; Becker, C.; Pelosi, P.; Knoll, W.; Larisika, M.; Nowak, C. Honey bee odorant-binding protein 14: Effects on thermal stability upon odorant binding revealed by FT-IR spectroscopy and CD measurements. Eur. Biophys. J. 2014, 43, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Mastrogiacomo, R.; Iovinella, I.; Tuccori, E.; Persaud, K.C. Structure and biotechnological applications of odorant-binding proteins. Appl. Microbiol. Biotechnol. 2014, 98, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Paolini, S.; Scaloni, A.; Amoresano, A.; Marchese, S.; Napolitano, E.; Pelosi, P. Amino acid sequence, post-translational modifications, binding and labelling of porcine odorant-binding protein. Chem. Senses 1998, 23, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Brandazza, A.; Pelosi, P. Binding of Polyclycic Aromatic Hydrocarbons to Mutants of Odorant-Binding Protein: A first step towards biosensors for environmental monitoring. Biochim. Biophys. Acta 2008, 1784, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Ban, L.; Song, L.M.; Liu, Y.; Pelosi, P.; Wang, G. General odorant-binding proteins and sex pheromone guide larvae of Plutella xylostella to better food. Insect Biochem. Mol. Biol. 2016, 72, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Hotel, O.; Poli, J.P.; Mer-Calfati, C.; Scorsone, E.; Saada, S. SAW Sensor’s Frequency Shift Characterization for Odor Recognition and Concentration Estimation. IEEE Sens. J. 2017, 17, 7011–7018. [Google Scholar] [CrossRef]
- Hoß, S.G.; Bendas, G. Mass-Sensitive Biosensor Systems to Determine the Membrane Interaction of Analytes. Methods Mol. Biol. 2017, 1520, 145–157. [Google Scholar] [PubMed]
- Di Pietrantonio, F.; Cannatà, D.; Benetti, M.; Verona, E.; Varriale, A.; Staiano, M.; D’Auria, S. Detection of odorant molecules via surface acoustic wave biosensor array based on odorant-binding proteins. Biosens. Bioelectron. 2013, 41, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Benilova, I.; Chegel, V.I.; Ushenin, Y.V.; Vidic, J.; Soldatkin, A.P.; Martelet, C.; Pajot, E.; Jaffrezic-Renault, N. Stimulation of human olfactory receptor 17–40 with odorants probed by surface plasmon resonance. Eur. Biophys. J. 2008, 37, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Sanmartí-Espinal, M.; Iavicoli, P.; Calò, A.; Taulés, M.; Galve, R.; Marco, M.P.; Samitier, J. Quantification of interacting cognate odorants with olfactory receptors in nanovesicles. Sci. Rep. 2017, 7, 17483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cennamo, N.; Di Giovanni, S.; Varriale, A.; Staiano, M.; Di Pietrantonio, F.; Notargiacomo, A.; Zeni, L.; D’Auria, S. Easy to use plastic optical fiber-based biosensor for detection of butanal. PLoS ONE 2015, 10, e0116770. [Google Scholar] [CrossRef] [PubMed]
- Larisika, M.; Kotlowski, C.; Steininger, C.; Mastrogiacomo, R.; Pelosi, P.; Schütz, S.; Peteu, S.F.; Kleber, C.; Reiner-Rozman, C.; Nowak, C.; et al. Electronic Olfactory Sensor Based on A. mellifera Odorant-Binding Protein 14 on a Reduced Graphene Oxide Field—Effect Transistor. Angew. Chem. Int. Ed. Engl. 2015, 54, 13245–13248. [Google Scholar] [CrossRef] [PubMed]
- Kotlowski, C.; Larisika, M.; Guerin, P.M.; Kleber, C.; Kröber, T.; Mastrogiacomo, R.; Nowak, C.; Schutz, S.; Schwaighofer, A.; Knoll, W. Fine discrimination of volatile compounds by graphene-immobilized odorant-binding proteins. Sens. Actuators B Chem. 2018, 256, 564–572. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, H.; Li, H.; Zhang, J.; Zhuang, S.; Zhang, F.; Hsia, K.J.; Wang, P. Impedance sensing and molecular modeling of an olfactory biosensor based on chemosensory proteins of honeybee. Biosens. Bioelectron. 2013, 40, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Li, H.; Zhuang, S.; Zhang, D.; Zhang, Q.; Zhou, J.; Dong, S.; Liu, Q.; Wang, P. Olfactory biosensor using odorant-binding proteins from honeybee: Ligands of floral odors and pheromones detection by electrochemical impedance. Sens. Actuators B Chem. 2014, 193, 420–427. [Google Scholar] [CrossRef]
- Lu, Y.; Yao, Y.; Zhang, Q.; Zhang, D.; Zhuang, S.; Li, H.; Liu, Q. Olfactory biosensor for insect semiochemicals analysis by impedance sensing of odorant-binding proteins on interdigitated electrodes. Biosens. Bioelectron. 2015, 67, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, D.; Zhang, Q.; Huang, Y.; Luo, S.; Yao, Y.; Li, S.; Liu, Q. Impedance spectroscopy analysis of human odorant binding proteins immobilized on nanopore arrays for biochemical detection. Biosens. Bioelectron. 2016, 79, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, E.; Magna, G.; Polese, D.; Vergara, A.; Schild, D.; Di Natale, C. Stable Odor Recognition by a neuro-adaptive Electronic Nose. Sci. Rep. 2015, 5, 10960. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Feature | Biological Noses | Artificial Noses |
|---|---|---|
| Type of sensing elements | Membrane receptors of broad overlapping specificity | Macromolecules with a moderate degree of specificity (OBPs, other binding proteins, DNA) |
| Number of sensing elements | About 300 in humans, up to 1000 in other mammals, around 100 in insects | At least 100 types to mimic the discrimination and the wide range of odours detected by the human nose |
| Coding strategy | Discrimination mainly on the basis of stereochemical parameters, but in some cases also of functional groups | Stereochemical parameters and functional groups |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pelosi, P.; Zhu, J.; Knoll, W. From Gas Sensors to Biomimetic Artificial Noses. Chemosensors 2018, 6, 32. https://doi.org/10.3390/chemosensors6030032
Pelosi P, Zhu J, Knoll W. From Gas Sensors to Biomimetic Artificial Noses. Chemosensors. 2018; 6(3):32. https://doi.org/10.3390/chemosensors6030032
Chicago/Turabian StylePelosi, Paolo, Jiao Zhu, and Wolfgang Knoll. 2018. "From Gas Sensors to Biomimetic Artificial Noses" Chemosensors 6, no. 3: 32. https://doi.org/10.3390/chemosensors6030032
APA StylePelosi, P., Zhu, J., & Knoll, W. (2018). From Gas Sensors to Biomimetic Artificial Noses. Chemosensors, 6(3), 32. https://doi.org/10.3390/chemosensors6030032