
COX5B-Mediated Bioenergetic Alterations Modulate Cell Growth and Anticancer Drug Susceptibility by Orchestrating Claudin-2 Expression in Colorectal Cancers


Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Western Blot
2.3. Immunohistochemical Staining (IHC)
2.4. Seahorse Assay
2.5. Cell Culture and Transfection
2.6. Cytochrome c Oxidase (COX) Activity Measurement
2.7. Cell Proliferation and Viability Assay
2.8. Transcriptomic Analysis
2.9. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.10. H2O2, Oligomycin, Antimycin, and NaN3 Treatment
2.11. Statistical Analysis
3. Results
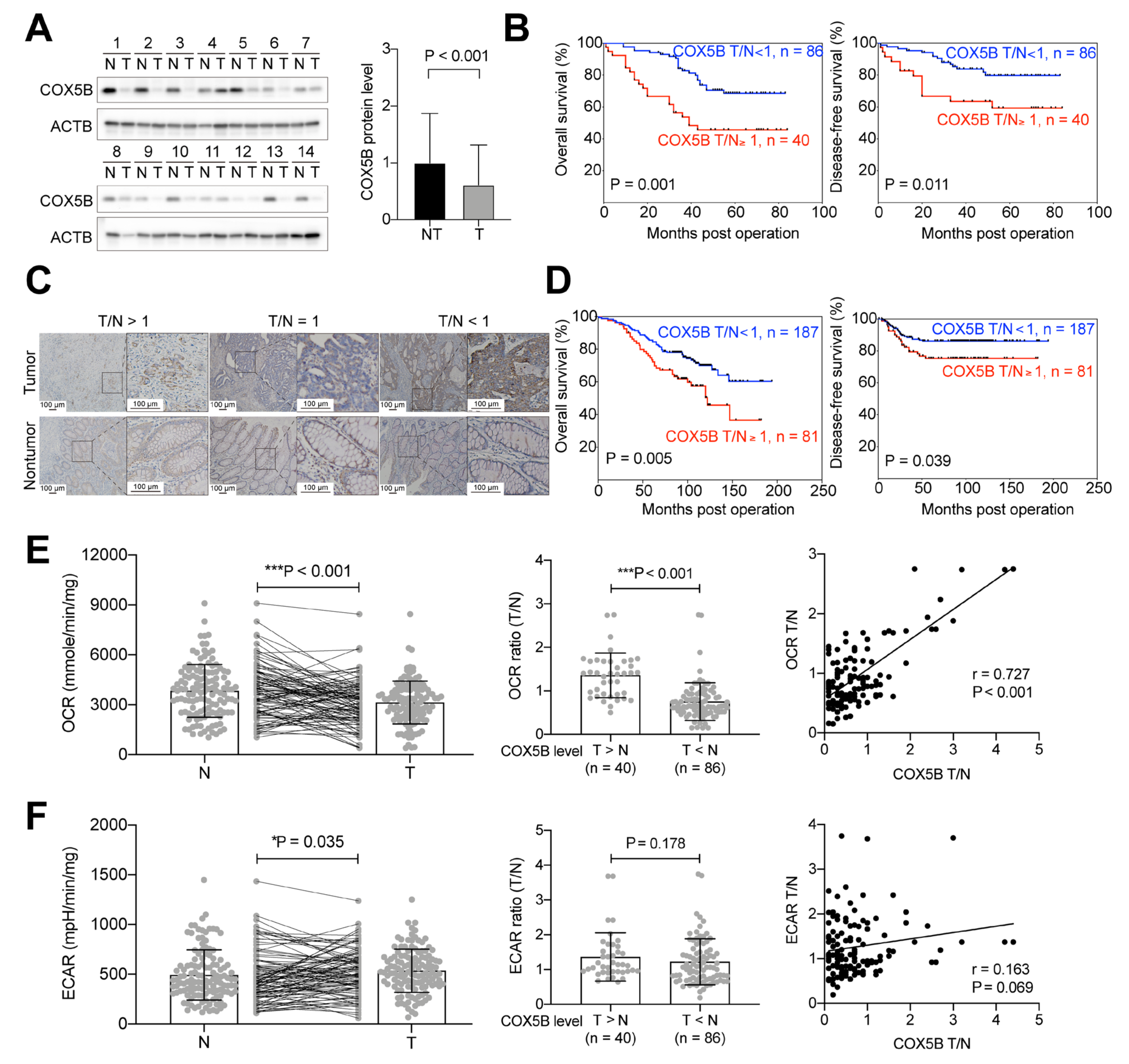
3.1. Higher COX5B Expression Levels in CRCs Correlate with Poorer Clinical Outcomes
3.2. Higher COX5B Expression Levels Correlate with Increased OCR in CRCs
3.3. Higher COX5B Expression Levels Correlate with Increased OCR in the Pre-Cancerous Tissues
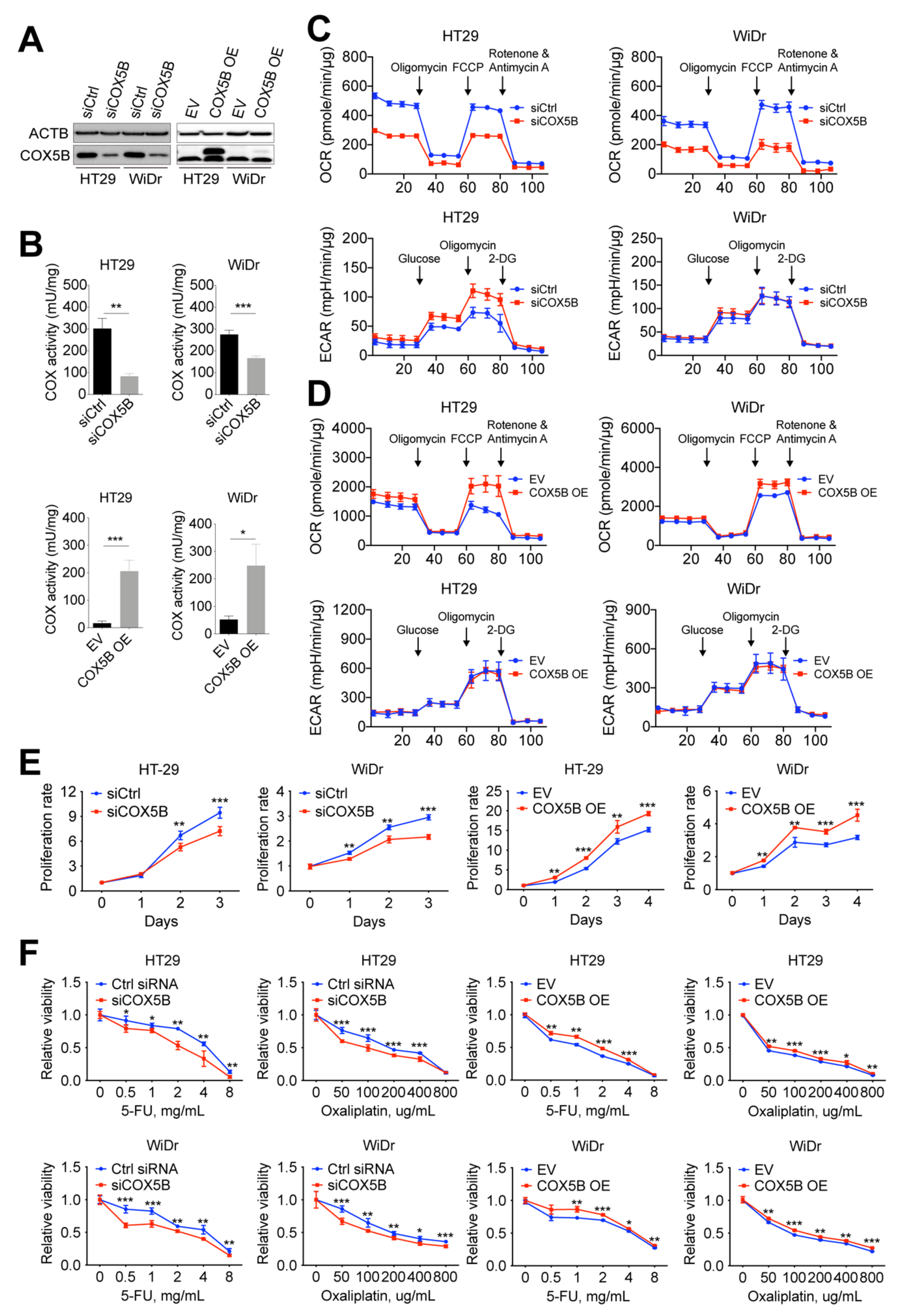
3.4. Change in COX5B Expression Influences Bioenergetic Alterations, Cell Growth, and Susceptibility to 5-Fluorouracil and Oxaliplatin in CRCs Cells
3.5. COX5B-Mediated Bioenergetic Alterations Orchestrated Claudin-2 Expression in CRCs Cells
3.6. Association of COX5B and CLDN2 Expression in Tissues from Patients with CRCs
3.7. CLDN2 Functions Downstream of COX5B to Modulate Ell Growth and Susceptibility to 5-FU and Oxaliplatin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vogelstein, B.; Fearon, E.R.; Hamilton, S.R.; Kern, S.E.; Preisinger, A.C.; Leppert, M.; Nakamura, Y.; White, R.; Smits, A.M.; Bos, J.L. Genetic alterations during colorectal-tumor development. N. Engl. J. Med. 1988, 319, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Isidoro, A.; Martínez, M.; Fernández, P.L.; Ortega, A.; Santamaría, G.S.; Chamorro, M.; Reed, J.C.; Cuezva, J.M. Alteration of the bioenergetic phenotype of mitochondria is a hallmark of breast, gastric, lung and oesophageal cancer. Biochem. J. 2004, 378, 17–20. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.R.; Chiang, J.M.; Lim, S.N.; Su, M.Y.; Chen, T.H.; Huang, S.W.; Chen, C.W.; Wu, R.C.; Tsai, C.L.; Lin, Y.H.; et al. Dynamic bioenergetic alterations in colorectal adenomatous polyps and adenocarcinomas. EBioMedicine 2019, 44, 334–345. [Google Scholar] [CrossRef] [Green Version]
- Cuezva, J.M.; Krajewska, M.; de Heredia, M.L.; Krajewski, S.; Santamaria, G.; Kim, H.; Zapata, J.M.; Marusawa, H.; Chamorro, M.; Reed, J.C. The bioenergetic signature of cancer: A marker of tumor progression. Cancer Res. 2002, 62, 6674–6681. [Google Scholar]
- Sánchez-Aragoó, M.A.; Cuezva, J.M. The bioenergetic signature of isogenic colon cancer cells predicts the cell death response to treatment with 3-bromopyruvate, iodoacetate or 5-fluorouracil. J. Transl. Med. 2011, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Fay, J.R.; Steele, V.; Crowell, J.A. Energy homeostasis and cancer prevention: The AMP-activated protein kinase. Cancer Prev. Res. 2009, 2, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Iommarini, L.; Ghelli, A.; Gasparre, G.; Porcelli, A.M. Mitochondrial metabolism and energy sensing in tumor progression. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 582–590. [Google Scholar] [CrossRef]
- Bost, F.; Decoux-Poullot, A.G.; Tanti, J.F.; Clavel, S. Energy disruptors: Rising stars in anticancer therapy? Oncogenesis 2016, 5, e188. [Google Scholar] [CrossRef] [Green Version]
- Herrera, A.S.; Esparza, M.a.d.C.A.; Ashraf, G.M.; Zamyatnin, A.A.; Aliev, G. Beyond mitochondria, what would be the energy source of the cell? Cent. Nerv. Syst. Agents. Med. Chem. 2015, 15, 32–41. [Google Scholar] [CrossRef]
- Warburg, O.; Wind, F.; Negelein, E. The metabolism of tumors in the body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Hagland, H.R.; Soreide, K. Cellular metabolism in colorectal carcinogenesis: Influence of lifestyle, gut microbiome and metabolic pathways. Cancer Lett. 2015, 356, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Ashton, T.M.; McKenna, W.G.; Kunz-Schughart, L.A.; Higgins, G.S. Oxidative phosphorylation as an emerging target in cancer therapy. Clin. Cancer Res. 2018, 24, 2482–2490. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, R.S.; Banerji, V. Targeting mitochondrial bioenergetics as a therapeutic strategy for chronic lymphocytic leukemia. Oxid. Med. Cell Longev. 2018, 2018, 2426712. [Google Scholar] [CrossRef]
- Formentini, L.; Martinez-Reyes, I.; Cuezva, J.M. The mitochondrial bioenergetic capacity of carcinomas. IUBMB Life 2010, 62, 554–560. [Google Scholar] [CrossRef]
- Ghosh, P.; Vidal, C.; Dey, S.; Zhang, L. Mitochondria targeting as an effective strategy for cancer therapy. Int. J. Mol. Sci. 2020, 21, 3363. [Google Scholar] [CrossRef]
- Schapira, A.H.V. Mitochondrial disease. Lancet 2006, 368, 70–82. [Google Scholar] [CrossRef]
- Kadenbach, B.; Huttemann, M.H.; Arnold, S.; Lee, I.; Bender, E. Mitochondrial energy metabolism is regulated via nuclear-coded subunits of cytochrome c oxidase. Free Radic. Biol. Med. 2000, 29, 211–221. [Google Scholar] [CrossRef]
- Ludwig, B.; Bender, E.; Arnold, S.; Hüttemann, M.; Lee, I.; Kadenbach, B. Cytochrome c oxidase and the regulation of oxidative phosphorylation. Chembiochem 2001, 2, 392–403. [Google Scholar] [CrossRef]
- Kadenbach, B. Regulation of mammalian 13-subunit cytochrome c oxidase and binding of other proteins: Role of NDUFA4. Trends Endocrinol. Metab. 2017, 28, 761–770. [Google Scholar] [CrossRef]
- Deng, S.; Li, Y.; Yi, G.; Lei, B.; Guo, M.; Xiang, W.; Chen, Z.; Liu, Y.; Qi, S. Overexpression of COX7A2 is associated with a good prognosis in patients with glioma. J. Neurooncol. 2018, 136, 41–50. [Google Scholar] [CrossRef]
- Gao, S.-P.; Sun, H.-F.; Fu, W.-Y.; Li, L.-D.; Zhao, Y.; Chen, M.-T.; Jin, W. High expression of COX5B is associated with poor prognosis in breast cancer. Future Oncol. 2017, 13, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Hewedi, I.H.; Farid, R.M.; Sidhom, K.F.; Salman, M.I.; Mostafa, R.G. Differential expression of cytochrome C oxidase subunit I along the colorectal adenoma: Carcinoma progression. Appl. Immunohistochem. Mol. Morphol. 2018, 26, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Le, P.H.; Huang, S.C.; Lim, S.N.; Chou, C.H.; Yeh, T.S.; Chen, T.C.; Yen, T.H.; Su, M.Y.; Chiu, C.T.; Yeh, C.T.; et al. Complex IV subunit 1 defect predicts postoperative survival in hepatocellular carcinoma. Oncol. Lett. 2014, 7, 1430–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, Y.D.; Lin, W.R.; Lin, Y.H.; Kuo, W.H.; Tseng, C.J.; Lim, S.N.; Huang, Y.L.; Huang, S.C.; Wu, T.J.; Lin, K.H.; et al. COX5B-mediated bioenergetic alteration regulates tumor growth and migration by modulating AMPK-UHMK1-ERK cascade in hepatoma. Cancers 2020, 12, 1646. [Google Scholar] [CrossRef]
- Chu, Y.D.; Lai, H.Y.; Pai, L.M.; Huang, Y.H.; Lin, Y.H.; Liang, K.H.; Yeh, C.T. The methionine salvage pathway-involving ADI1 inhibits hepatoma growth by epigenetically altering genes expression via elevating S-adenosylmethionine. Cell Death Dis. 2019, 10, 240. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.W.; Chu, Y.D.; Lai, M.W.; Lin, C.L.; Liang, K.H.; Lin, Y.H.; Yeh, C.T. Hepatitis B virus covalently closed circular DNA predicts postoperative liver cancer metastasis independent of virological suppression. Cancers 2021, 13, 538. [Google Scholar] [CrossRef]
- Lin, C.L.; Chu, Y.D.; Yeh, C.T. Emergence of oncogenic-enhancing hepatitis B virus X gene mutants in patients receiving suboptimal entecavir treatment. Hepatology 2019, 69, 2292–2296. [Google Scholar] [CrossRef]
- Chu, Y.D.; Chen, H.K.; Huang, T.; Chan, S.P. A novel function for the DEAD-box RNA helicase DDX-23 in primary microRNA processing in Caenorhabditis elegans. Dev. Biol. 2016, 409, 459–472. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.-P.; Sun, H.-F.; Jiang, H.-L.; Li, L.-D.; Hu, X.; Xu, X.-E.; Jin, W. Loss of COX5B inhibits proliferation and promotes senescence via mitochondrial dysfunction in breast cancer. Oncotarget 2015, 6, 43363–53374. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Xi, J. Identification of COX5B as a novel biomarker in high-grade glioma patients. Onco. Targets Ther. 2017, 10, 5463–5470. [Google Scholar] [CrossRef] [Green Version]
- Stein, J.; Tenbrock, J.; Kristiansen, G.; Muller, S.C.; Ellinger, J. Systematic expression analysis of the mitochondrial respiratory chain protein subunits identifies COX5B as a prognostic marker in clear cell renal cell carcinoma. Int. J. Urol. 2019, 26, 910–916. [Google Scholar] [CrossRef]
- Krupar, R.; Hautmann, M.G.; Pathak, R.R.; Varier, I.; McLaren, C.; Gaag, D.; Hellerbrand, C.; Evert, M.; Laban, S.; Idel, C.; et al. Immunometabolic determinants of chemoradiotherapy response and survival in head and neck squamous cell carcinoma. Am. J. Pathol. 2018, 188, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Soro-Arnaiz, I.; Li, Q.O.Y.; Torres-Capelli, M.; Melendez-Rodriguez, F.; Veiga, S.; Veys, K.; Sebastian, D.; Elorza, A.; Tello, D.; Hernansanz-Agustin, P.; et al. Role of mitochondrial complex IV in age-dependent obesity. Cell Rep. 2016, 16, 2991–3002. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, R.; Zhang, H.; Kim, J.W.; Shimoda, L.; Dang, C.V.; Semenza, G.L. HIF-1 regulates cytochrome oxidase subunits to optimize efficiency of respiration in hypoxic cells. Cell 2007, 129, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Fraisl, P.; Mazzone, M.; Schmidt, T.; Carmeliet, P. Regulation of angiogenesis by oxygen and metabolism. Dev. Cell 2009, 16, 167–179. [Google Scholar] [CrossRef]
- Dhawan, P.; Ahmad, R.; Chaturvedi, R.; Smith, J.J.; Midha, R.; Mittal, M.K.; Krishnan, M.; Chen, X.; Eschrich, S.; Yeatman, T.J.; et al. Claudin-2 expression increases tumorigenicity of colon cancer cells: Role of epidermal growth factor receptor activation. Oncogene 2011, 30, 3234–3247. [Google Scholar] [CrossRef] [Green Version]
- Paquet-Fifield, S.; Koh, S.L.; Cheng, L.; Beyit, L.M.; Shembrey, C.; Molck, C.; Behrenbruch, C.; Papin, M.; Gironella, M.; Guelfi, S.; et al. Tight junction protein claudin-2 promotes self-renewal of human colorectal cancer stem-like cells. Cancer Res. 2018, 78, 2925–2938. [Google Scholar] [CrossRef] [Green Version]
- Tabaries, S.; Annis, M.G.; Lazaris, A.; Petrillo, S.K.; Huxham, J.; Abdellatif, A.; Palmieri, V.; Chabot, J.; Johnson, R.M.; Van Laere, S.; et al. Claudin-2 promotes colorectal cancer liver metastasis and is a biomarker of the replacement type growth pattern. Commun. Biol. 2021, 4, 657. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Parameters | Univariate Analysis | Multivariate Analysis | ||
|---|---|---|---|---|
| HR (95% CI) | P | HR (95% CI) | P | |
| For OS | ||||
| COX5B level, high expression = 1 | 2.451 (1.363–4.406) | 0.003 | 2.424 (1.220–4.815) | 0.011 |
| Gender, male = 1 | 2.581 (1.368–4.867) | 0.003 | 3.713 (1.182–11.664) | 0.025 |
| Age, per year increase | 1.000 (0.974–1.027) | 0.982 | ||
| Height, per cm increase | 1.054 (1.011–1.098) | 0.013 | 0.971 (0.911–1.036) | 0.373 |
| Weight, per kg increase | 0.998 (0.968–1.029) | 0.913 | ||
| BMI, per kg/m2 increase | 0.918 (0.831–1.014) | 0.091 | ||
| Tumor location, left-handed = 1 | 0.903 (0.486–1.679) | 0.747 | ||
| Differentiation, poor = 1 | 2.483 (1.100–5.607) | 0.029 | 1.234 (0.443–3.432) | 0.688 |
| Local invasion, per grade increase | 1.702 (1.070–2.708) | 0.025 | 1.707 (0.913–3.191) | 0.094 |
| Tumor size, per cm increase | 1.003 (1.001–1.004) | 0.006 | 1.001 (0.999–1.003) | 0.196 |
| CEA, per ng/mL increase | 1.004 (1.002–1.005) | <0.001 | 1.004 (1.002–1.005) | <0.001 |
| Adjuvant chemotherapy, yes = 1 | 1.129 (0.627–2.034) | 0.686 | ||
| Distant to serosa, per mm increase | 0.923 (0.845–1.008) | 0.073 | ||
| OCR T/N, per number increase | 2.676 (1.713–4.180) | <0.001 | 2.105 (1.157–3.830) | 0.015 |
| ECAR T/N, per number increase | 1.610 (1.123–2.309) | 0.010 | 1.683 (1.066–2.658) | 0.025 |
| For DFS | ||||
| COX5B level, high expression = 1 | 3.047 (1.503–6.177) | 0.002 | 3.676 (1.514–8.927) | 0.004 |
| Gender, male = 1 | 2.439 (1.143–5.208) | 0.021 | 8.162 (1.336–49.870) | 0.023 |
| Age, per year increase | 0.953 (0.923–0.984) | 0.003 | 0.938 (0.893–0.985) | 0.011 |
| Height, per cm increase | 1.067 (1.018–1.118) | 0.007 | 0.892 (0.809–0.984) | 0.023 |
| Weight, per kg increase | 1.031 (1.000–1.064) | 0.052 | ||
| BMI, per kg/m2 increase | 1.019 (0.938–1.106) | 0.663 | ||
| Tumor location, left-handed = 1 | 1.649 (0.710–3.830) | 0.245 | ||
| Differentiation, poor = 1 | 4.547 (2.021–10.228) | <0.001 | 5.018 (1.619–15.547) | 0.005 |
| Local invasion, per grade increase | 2.048 (1.171–3.583) | 0.012 | 1.239 (0.581–2.642) | 0.580 |
| Tumor size, per cm increase | 1.002 (1.000–1.004) | 0.014 | 1.000 (0.998–1.002) | 0.896 |
| CEA, per ng/mL increase | 1.003 (1.001–1.004) | <0.001 | 1.002 (1.000–1.004) | 0.060 |
| Adjuvant chemotherapy, yes = 1 | 4.158 (1.705–10.139) | 0.002 | 4.186 (1.391–12.601) | 0.011 |
| Distant to serosa, per mm increase | 0.895 (0.789–1.015) | 0.083 | ||
| OCR T/N, per number increase | 3.986 (2.428–6.546) | <0.001 | 4.005 (2.049–7.827) | <0.001 |
| ECAR T/N, per number increase | 1.753 (1.165–2.639) | 0.007 | 1.512 (0.878–2.601) | 0.136 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, Y.-D.; Lim, S.-N.; Yeh, C.-T.; Lin, W.-R. COX5B-Mediated Bioenergetic Alterations Modulate Cell Growth and Anticancer Drug Susceptibility by Orchestrating Claudin-2 Expression in Colorectal Cancers. Biomedicines 2022, 10, 60. https://doi.org/10.3390/biomedicines10010060
Chu Y-D, Lim S-N, Yeh C-T, Lin W-R. COX5B-Mediated Bioenergetic Alterations Modulate Cell Growth and Anticancer Drug Susceptibility by Orchestrating Claudin-2 Expression in Colorectal Cancers. Biomedicines. 2022; 10(1):60. https://doi.org/10.3390/biomedicines10010060
Chicago/Turabian StyleChu, Yu-De, Siew-Na Lim, Chau-Ting Yeh, and Wey-Ran Lin. 2022. "COX5B-Mediated Bioenergetic Alterations Modulate Cell Growth and Anticancer Drug Susceptibility by Orchestrating Claudin-2 Expression in Colorectal Cancers" Biomedicines 10, no. 1: 60. https://doi.org/10.3390/biomedicines10010060
APA StyleChu, Y. -D., Lim, S. -N., Yeh, C. -T., & Lin, W. -R. (2022). COX5B-Mediated Bioenergetic Alterations Modulate Cell Growth and Anticancer Drug Susceptibility by Orchestrating Claudin-2 Expression in Colorectal Cancers. Biomedicines, 10(1), 60. https://doi.org/10.3390/biomedicines10010060