
Expression Profiles of CDKN2A, MDM2, E2F2 and LTF Genes in Oral Squamous Cell Carcinoma

 , , ,
, , , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient and Samples
2.2. RNA Extraction
2.3. Complementary DNA (cDNA) Synthesis
2.4. Gene Expression Analysis
2.5. Statistical Analysis
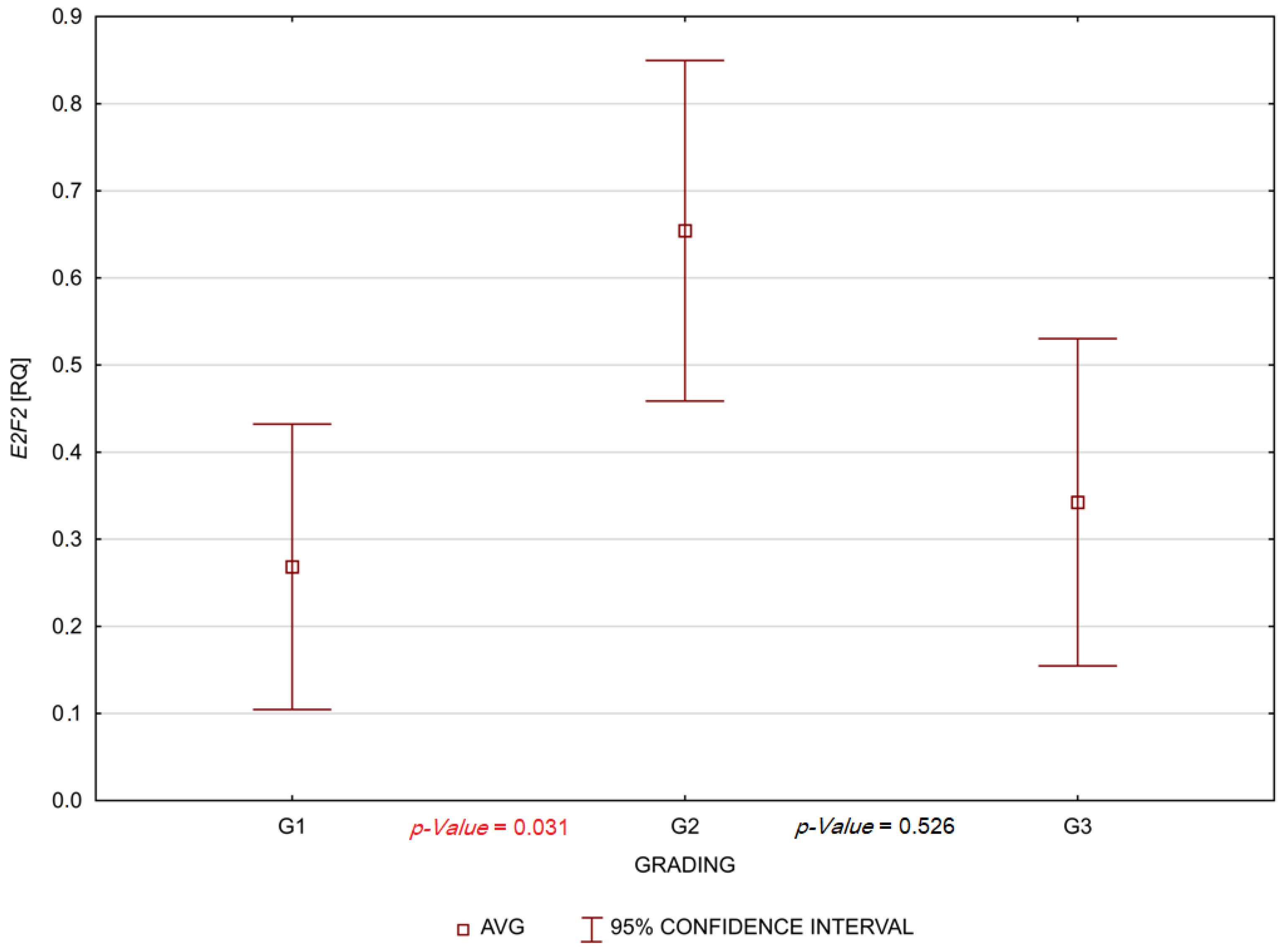
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 2096–2249. [Google Scholar] [CrossRef] [PubMed]
- Gollin, S.M. Cytogenetic alterations and their molecular genetic correlates in head and neck squamous cell carcinoma: A next generation window to the biology of disease. Genes Chromosom. Cancer 2014, 53, 972–990. [Google Scholar] [CrossRef] [PubMed]
- Cho, U.; Sung, Y.E.; Kim, M.S.; Lee, Y.S. Prognostic Role of Systemic Inflammatory Markers in Patients Undergoing Surgical Resection for Oral Squamous Cell Carcinoma. Biomedicines 2022, 10, 1268. [Google Scholar] [CrossRef]
- Gallagher, S.; Kefford, R.F.; Rizos, H. Enforced expression of p14ARF induces p53-dependent cell cycle arrest but not apoptosis. Cell Cycle 2005, 4, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Ausoni, S.; Boscolo-Rizzo, P.; Singh, B.; Da Mosto, M.C.; Spinato, G.; Tirelli, G.; Spinato, R.; Azzarello, G. Targeting cellular and molecular drivers of head and neck squamous cell carcinoma: Current options and emerging perspectives. Cancer Metastasis Rev. 2016, 35, 413–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, H.; Sun, D.; Zhang, X. The role of MDM2 amplification and overexpression in therapeutic resistance of malignant tumors. Cancer Cell Int. 2019, 19, 216. [Google Scholar] [CrossRef] [Green Version]
- Rayburn, E.; Zhang, R.; He, J.; Wang, H. MDM2 and human malig-nancies: Expression, clinical pathology, prognostic markers, and implications for chemotherapy. Curr. Cancer Drug Targets 2005, 5, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, S.; Zhang, Y.; Pan, J. The E2F transcription factor 2: What do we know? BioSci. Trends 2021, 15, 83–92. [Google Scholar] [CrossRef]
- Cutone, A.; Rosa, L.; Ianiro, G.; Lepanto, M.S.; Bonaccorsi di Patti, M.C.; Valenti, P.; Musci, G. Lactoferrin’s Anti-Cancer Properties: Safety, Selectivity, and Wide Range of Action. Biomolecules 2020, 15, 456. [Google Scholar] [CrossRef] [Green Version]
- Bukowska-Ośko, I.; Sulejczak, D.; Kaczyńska, K.; Kleczkowska, P.; Kramkowski, K.; Popiel, M.; Wietrak, E.; Kowalczyk, P. Lactoferrin as a Human Genome “Guardian”—An Overall Point of View. Int. J. Mol. Sci. 2022, 23, 5248. [Google Scholar] [CrossRef]
- Brunner, M.; Ng, B.C.; Veness, M.J.; Clark, J.R. Comparison of the AJCC N staging system in mucosal and cutaneous squamous head and neck cancer. Laryngoscope 2014, 124, 1598–1602. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, P.C.; Miguel, M.C.; Bagordakis, E.; Fonseca, F.P.; de Aquino, S.N.; Santos-Silva, A.R.; Lopes, M.A.; Graner, E.; Salo, T.; Kowalski, L.P.; et al. Clinicopathological prognostic factors of oral tongue squamous cell carcinoma: A retrospective study of 202 cases. Int. J. Oral. Maxillofac. Surg. 2014, 43, 795–801. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, A.K.; Chan, J.K.C.; Grandis, J.R.; Takata, T.; Slootweg, P.J. WHO Classification of Head and Neck Tumours International Agency for Research on Cancer (IARC), 4th ed.; IARC Publications: Lyon, France, 2017. [Google Scholar]
- Allameh, A.; Moazeni-Roodi, A.; Harirchi, I.; Ravanshad, M.; Motiee-Langroudi, M.; Garajei, A.; Hamidavi, A.; Mesbah-Namin, S.A. Promoter DNA Methylation and mRNA Expression Level of p16 Gene in Oral Squamous Cell Carcinoma: Correlation with Clinicopathological Characteristics. Pathol. Oncol. Res. 2019, 25, 1535–1543. [Google Scholar] [CrossRef]
- Biesaga, B.; Smolarczyk, R.; Mucha-Małecka, A.; Czapla, J.; Ryś, J.; Małecki, K. Prognostic Significance of STING Immunoexpression in Relation to HPV16 Infection in Patients with Squamous Cell Carcinomas of Oral Cavity and Oropharynx. Biomedicines 2022, 10, 2538. [Google Scholar] [CrossRef] [PubMed]
- Omolaoye, T.S.; El Shahawy, O.; Skosana, B.T.; Boillat, T.; Loney, T.; du Plessis, S.S. The mutagenic effect of tobacco smoke on male fertility. Env. Sci. Pollut. Res. 2022, 29, 62055–62066. [Google Scholar] [CrossRef]
- König, F.; Krekeler, G.; Hönig, J.F.; Cordon-Cardo, C.; Fischer, G.; Korabiowska, M. Relation between human papillomavirus positivity and p16 expression in head and neck carcinomas--a tissue microarray study. Anticancer Res. 2007, 27, 283–288. [Google Scholar] [PubMed]
- Li, J.; Poi, M.J.; Tsai, M.D. Regulatory mechanisms of tumor suppressor p16(INK4A) and their relevance to cancer. Biochemistry 2011, 50, 5566–5582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Naggar, A.K.; Westra, W.H. p16 expression as a surrogate marker for HPV-related oropharyngeal carcinoma: A guide for interpretative relevance and consistency. Head Neck 2012, 34, 459–461. [Google Scholar] [CrossRef]
- Wang, D.; Grecula, J.C.; Gahbauer, R.A.; Schuller, D.E.; Jatana, K.R.; Biancamano, J.D.; Lang, J.C. p16 gene alterations in locally advanced squamous cell carcinoma of the head and neck. Oncol. Rep. 2006, 15, 661–665. [Google Scholar] [CrossRef] [Green Version]
- Su, P.F.; Huang, W.L.; Wu, H.T.; Wu, C.H.; Liu, T.Y.; Kao, S.Y. p16(INK4A) promoter hypermethylation is associated with invasiveness and prognosis of oral squamous cell carcinoma in an age-dependent manner. Oral Oncol. 2010, 46, 734–739. [Google Scholar] [CrossRef]
- Cabanillas, R.; Astudillo, A.; Valle, M.; de la Rosa, J.; Álvarez, R.; Durán, N.S.; Cadiñanos, J. Novel germline CDKN2A mutation associated with head and neck squamous cell carcinomas and melanomas. Head Neck 2013, 35, E80–E84. [Google Scholar] [CrossRef] [PubMed]
- Padhi, S.S.; Roy, S.; Kar, M.; Saha, A.; Roy, S.; Adhya, A.; Baisakh, M.; Banerjee, B. Role of CDKN2A/p16 expression in the prognostication of oral squamous cell carcinoma. Oral Oncol. 2017, 73, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Yalniz, Z.; Demokan, S.; Karabulut, B.; Ulusan, M.; Suoglu, Y.; Dalay, N. Copy number profiling of tumor suppressor genes in head and neck cancer. Head Neck 2017, 39, 341–346. [Google Scholar] [CrossRef]
- Jie, W.; Bai, J.; Yan, J.; Chi, Y.; Li, B.B. Multi-Site Tumour Sampling Improves the Detection of Intra-Tumour Heterogeneity in Oral and Oropharyngeal Squamous Cell Carcinoma. Front. Med. 2021, 8, 670305. [Google Scholar] [CrossRef]
- Bhatia, V.; Goel, M.M.; Makker, A.; Tewari, S.; Yadu, A.; Shilpi, P.; Kumar, S.; Agarwal, S.P.; Goel, S.K. Promoter region hypermethylation and mRNA expression of MGMT and p16 genes in tissue and blood samples of human premalignant oral lesions and oral squamous cell carcinoma. Biomed Res. Int. 2014, 2014, 248419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Sayáns, M.; Suárez-Peñaranda, J.M.; Padín-Iruegas, M.E.; Gayoso-Diz, P.; Reis-De Almeida, M.; Barros-Angueira, F.; Gándara-Vila, P.; Blanco-Carrión, A.; García-García, A. The Loss of p16 Expression Worsens the Prognosis of OSCC. Appl. Immunohistochem. Mol. Morphol. 2015, 23, 724–732. [Google Scholar] [CrossRef]
- Salehinejad, J.; Sharifi, N.; Amirchaghmaghi, M.; Ghazi, N.; Shakeri, M.T.; Ghazi, A. Immunohistochemical expression of p16 protein in oral squamous cell carcinoma and lichen planus. Ann. Diagn. Pathol. 2014, 18, 210–213. [Google Scholar] [CrossRef]
- Jitani, A.K.; Raphael, V.; Mishra, J.; Shunyu, N.B.; Khonglah, Y.; Medhi, J. Analysis of Human Papilloma Virus 16/18 DNA and its Correlation with p16 Expression in Oral Cavity Squamous Cell Carcinoma in North-Eastern India: A Chromogenic in-situ Hybridization Based Study. J. Clin. Diagn. Res. 2015, 9, EC04–EC07. [Google Scholar] [CrossRef]
- Wang, S.; Li, T.; Liu, H.; Wei, W.; Yang, Y.; Wang, C.; Li, B.; Han, Z.; Feng, Z. A Combined Prediction Model for Lymph Node Metastasis Based on a Molecular Panel and Clinicopathological Factors in Oral Squamous Cell Carcinoma. Front. Oncol. 2021, 11, 660615. [Google Scholar] [CrossRef]
- Millon, R.; Muller, D.; Schultz, I.; Salvi, R.; Ghnassia, J.P.; Frebourg, T.; Wasylyk, B.; Abecassis, J. Loss of MDM2 expression in human head and neck squamous cell carcinomas and clinical significance. Oral Oncol. 2001, 37, 620–631. [Google Scholar] [CrossRef]
- Carroll, P.E.; Okuda, M.; Horn, H.F.; Biddinger, P.; Stambrook, P.J.; Gleich, L.L.; Li, Y.Q.; Tarapore, P.; Fukasawa, K. Centrosome hyperamplification in human cancer: Chromosome instability induced by p53 mutation and/or Mdm2 overexpression. Oncogene 1999, 18, 1935–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentin-Vega, Y.A.; Barboza, J.A.; Chau, G.P.; El-Naggar, A.K.; Lozano, G. High levels of the p53 inhibitor MDM4 in head and neck squamous carcinomas. Hum. Pathol. 2007, 38, 1553–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesland, S.; Kanter-Lewensohn, L.; Tell, R.; Munck-Wikland, E.; Lewensohn, R.; Nilsson, A. Expression of Ku86 confers favorable outcome of tonsillar carcinoma treated with radiotherapy. Head Neck 2003, 25, 313–321. [Google Scholar] [CrossRef]
- Dong, Y.; Zou, J.; Su, S.; Huang, H.; Deng, Y.; Wang, B.; Li, W. MicroRNA-218 and microRNA-520a inhibit cell proliferation by downregulating E2F2 in hepatocellular carcinoma. Mol. Med. Rep. 2015, 12, 1016–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Li, T.; Yang, L.H. E2F2 induces MCM4, CCNE2 and WHSC1 upregulation in ovarian cancer and predicts poor overall survival. Eur. Rev. Med. Pharm. Sci. 2017, 21, 2150–2156. [Google Scholar]
- Zhou, X.; Tao, H. Overexpression of microRNA-936 suppresses non-small cell lung cancer cell proliferation and invasion via targeting E2F2. Exp. Ther. Med. 2018, 16, 2696–2702. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.Y.; Wang, J.Q.; Wu, L.L.; Zheng, W.E.; Chen, P.R. miR-638 represses the stem cell characteristics of breast cancer cells by targeting E2F2. Breast Cancer 2020, 27, 147–158. [Google Scholar] [CrossRef]
- Liao, P.; Han, S.; Qu, H. Expression, Prognosis, and Immune Infiltrates Analyses of E2Fs in Human Brain and CNS Cancer. Biomed Res. Int. 2020, 2020, 6281635. [Google Scholar] [CrossRef]
- Li, Y.; Sturgis, E.M.; Zhu, L.; Cao, X.; Wei, Q.; Zhang, H.; Li, G. E2F transcription factor 2 variants as predictive biomarkers for recurrence risk in patients with squamous cell carcinoma of the oropharynx. Mol. Carcinog. 2017, 56, 1335–1343. [Google Scholar] [CrossRef]
- Li, T.; Luo, W.; Liu, K.; Lv, X.; Xi, T. miR-31 promotes proliferation of colon cancer cells by targeting E2F2. Biotechnol. Lett. 2015, 37, 523–532. [Google Scholar] [CrossRef]
- Gao, Y.; Ma, X.; Yao, Y.; Li, H.; Fan, Y.; Zhang, Y.; Zhao, C.; Wang, L.; Ma, M.; Lei, Z.; et al. miR-155 regulates the proliferation and invasion of clear cell renal cell carcinoma cells by targeting E2F2. Oncotarget 2016, 7, 20324–20337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Han, D.; Wei, W.; Cao, W.; Zhang, R.; Dong, Q.; Zhang, J.; Wang, Y.; Liu, N. MiR-218 Inhibited Growth and Metabolism of Human Glioblastoma Cells by Directly Targeting E2F2. Cell. Mol. Neurobiol. 2015, 35, 1165–1173. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, X.; Liu, Y.; Ni, Z.; Lin, Y.; Duan, Z.; Shi, Y.; Wang, G.; Li, F. Downregulated miR-31 level associates with poor prognosis of gastric cancer and its restoration suppresses tumor cell malignant phenotypes by inhibiting E2F2. Oncotarget 2016, 7, 36577–36589. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.Q.; Li, M.C.; Zhang, Y.Q.; Liu, X.G. MiR-490-5p inhibits the metastasis of hepatocellular carcinoma by down-regulating E2F2 and ECT2. J. Cell Biochem. 2018, 119, 8317–8324. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Xiao, D.; Cui, Y.; Wang, X. Exosomes-Derived Long Non-Coding RNA HOTAIR Reduces Laryngeal Cancer Radiosensitivity by Regulating microRNA-454-3p/E2F2 Axis. Onco Targets Ther. 2019, 12, 10827–10839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, M.; Sawada, K.; Nakamura, K.; Yoshimura, A.; Ishida, K.; Kobayashi, M.; Shimizu, A.; Yamamoto, M.; Kodama, M.; Hashimoto, K.; et al. Paclitaxel exposure downregulates miR-522 expression and its downregulation induces paclitaxel resistance in ovarian cancer cells. Sci. Rep. 2020, 10, 16755. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Sun, S.Z.; Cheng, F.; Zhang, N.Q. Mediation of circ_RPPH1 on miR-146b-3p/E2F2 pathway to hinder the growth and metastasis of breast carcinoma cells. Aging 2021, 13, 20552–20568. [Google Scholar] [CrossRef]
- Zhang, H.; Qiu, X.; Song, Z.; Lan, L.; Ren, X.; Ye, B. CircCUL2 suppresses retinoblastoma cells by regulating miR-214-5p/E2F2 Axis. Anticancer Drugs 2022, 33, e218–e227. [Google Scholar] [CrossRef]
- Shaheduzzaman, S.; Vishwanath, A.; Furusato, B.; Cullen, J.; Chen, Y.; Bañez, L.; Nau, M.; Ravindranath, L.; Kim, K.H.; Mohammed, A.; et al. Silencing of Lactotransferrin expression by methylation in prostate cancer progression. Cancer Biol. Ther. 2007, 6, 1088–1095. [Google Scholar] [CrossRef] [Green Version]
- Chiu, I.J.; Hsu, Y.H.; Chang, J.S.; Yang, J.C.; Chiu, H.W.; Lin, Y.F. Lactotransferrin Downregulation Drives the Metastatic Progression in Clear Cell Renal Cell Carcinoma. Cancers 2020, 12, 847. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Fan, S.; Zou, G.; Shi, L.; Zeng, Z.; Ma, J.; Zhou, Y.; Li, X.; Zhang, X.; Li, X.; et al. Lactotransferrin could be a novel independent molecular prognosticator of nasopharyngeal carcinoma. Tumor Biol. 2015, 36, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ling, T.; Wu, H.; Wang, K. Re-expression of Lactotransferrin, a candidate tumor suppressor inactivated by promoter hypermethylation, impairs the malignance of oral squamous cell carcinoma cells. J. Oral. Pathol. Med. 2015, 44, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.M.; Haffner, M.C.; Kulac, I.; Maynard, J.P.; Baena-Del Valle, J.A.; Isaacs, W.B.; Yegnasubramanian, S.; De Marzo, A.M.; Sfanos, K.S. Lactoferrin CpG Island Hypermethylation and Decoupling of mRNA and Protein Expression in the Early Stages of Prostate Carcinogenesis. Am. J. Pathol. 2019, 189, 2311–2322. [Google Scholar] [CrossRef]
- Deng, M.; Zhang, W.; Tang, H.; Ye, Q.; Liao, Q.; Zhou, Y.; Wu, M.; Xiong, W.; Zheng, Y.; Guo, X.; et al. Lactotransferrin acts as a tumor suppressor in nasopharyngeal carcinoma by repressing AKT through multiple mechanisms. Oncogene 2013, 32, 4273–4283. [Google Scholar] [CrossRef] [Green Version]
- Slaughter, D.P.; Southwick, H.W.; Smejkal, W. Field cancerization in oral stratified squamous epithelium; clinical implications of multicentric origin. Cancer 1953, 6, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Braakhuis, B.J.; Tabor, M.P.; Kummer, J.A.; Leemans, C.R.; Brakenhoff, R.H. A genetic explanation of Slaughter’s concept of field cancerization: Evidence and clinical implications. Cancer Res. 2003, 63, 1727–1730. [Google Scholar] [PubMed]
- Hoque Apu, E.; Akram, S.U.; Rissanen, J.; Wan, H.; Salo, T. Desmoglein 3—Influence on oral carcinoma cell migration and invasion. Exp. Cell Res. 2018, 370, 353–364. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Clinical Parameters | Patients, n (%) |
|---|---|
| Histological grading | |
| G1 (Well differentiated) | 14 (16) |
| G2 (Moderately differentiated) | 60 (69) |
| G3 (Poorly differentiated) | 13 (15) |
| T classification | |
| T1 | 8 (9) |
| T2 | 24 (27.3) |
| T3 | 23 (26.2) |
| T4 | 33 (37.5) |
| Nodal status | |
| N0 | 40 (45, 5) |
| N1 | 25 (28, 4) |
| N2 | 20 (22, 7) |
| N3 | 3 (3, 4) |
| Gene | Mean RQ ± SD | r | p-Value | |
|---|---|---|---|---|
| Tumour | Margin | |||
| CDKN2A | 2.08 ± 4.99 | 0.45 ± 0.99 | −0.031 | 0.782 |
| MDM2 | 0.7 ± 0.8 | 0.68 ± 0.39 | 0.188 | 0.088 |
| E2F2 | 0.54 ± 0.65 | 0.5 ± 0.44 | 0.208 | 0.061 |
| LTF | 0.04 ± 0.13 | 0.13 ± 0.29 | 0.097 | 0.378 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gołąbek, K.; Rączka, G.; Gaździcka, J.; Miśkiewicz-Orczyk, K.; Zięba, N.; Krakowczyk, Ł.; Misiołek, M.; Strzelczyk, J.K. Expression Profiles of CDKN2A, MDM2, E2F2 and LTF Genes in Oral Squamous Cell Carcinoma. Biomedicines 2022, 10, 3011. https://doi.org/10.3390/biomedicines10123011
Gołąbek K, Rączka G, Gaździcka J, Miśkiewicz-Orczyk K, Zięba N, Krakowczyk Ł, Misiołek M, Strzelczyk JK. Expression Profiles of CDKN2A, MDM2, E2F2 and LTF Genes in Oral Squamous Cell Carcinoma. Biomedicines. 2022; 10(12):3011. https://doi.org/10.3390/biomedicines10123011
Chicago/Turabian StyleGołąbek, Karolina, Grzegorz Rączka, Jadwiga Gaździcka, Katarzyna Miśkiewicz-Orczyk, Natalia Zięba, Łukasz Krakowczyk, Maciej Misiołek, and Joanna Katarzyna Strzelczyk. 2022. "Expression Profiles of CDKN2A, MDM2, E2F2 and LTF Genes in Oral Squamous Cell Carcinoma" Biomedicines 10, no. 12: 3011. https://doi.org/10.3390/biomedicines10123011
APA StyleGołąbek, K., Rączka, G., Gaździcka, J., Miśkiewicz-Orczyk, K., Zięba, N., Krakowczyk, Ł., Misiołek, M., & Strzelczyk, J. K. (2022). Expression Profiles of CDKN2A, MDM2, E2F2 and LTF Genes in Oral Squamous Cell Carcinoma. Biomedicines, 10(12), 3011. https://doi.org/10.3390/biomedicines10123011