


Fecal Microbiota Transplantation Could Improve Chronic Diarrhea in Cynomolgus Monkey by Alleviating Inflammation and Modulating Gut Microbiota
 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects and Treatment Outcomes
2.2. ELISA
2.3. Metagenomics and Functional Prediction Analyses
2.4. High-Throughput Amplicon Sequencing
2.5. Quantification and Statistical Analysis
3. Results
3.1. FMT Ameliorates Diarrhea in Captive Monkeys
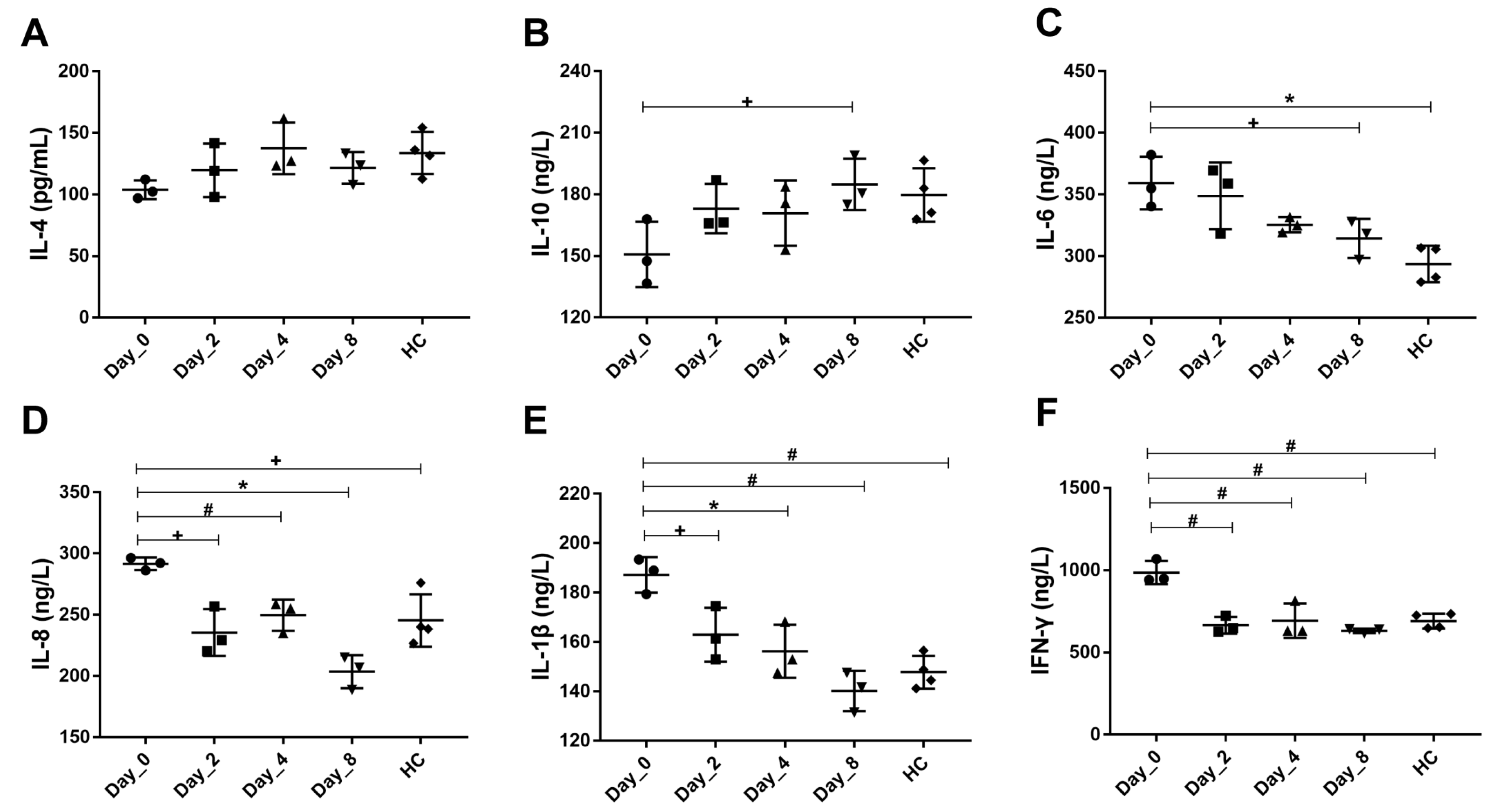
3.2. FMT Mitigates Serum Inflammation Response in Chronic Diarrhea Monkeys
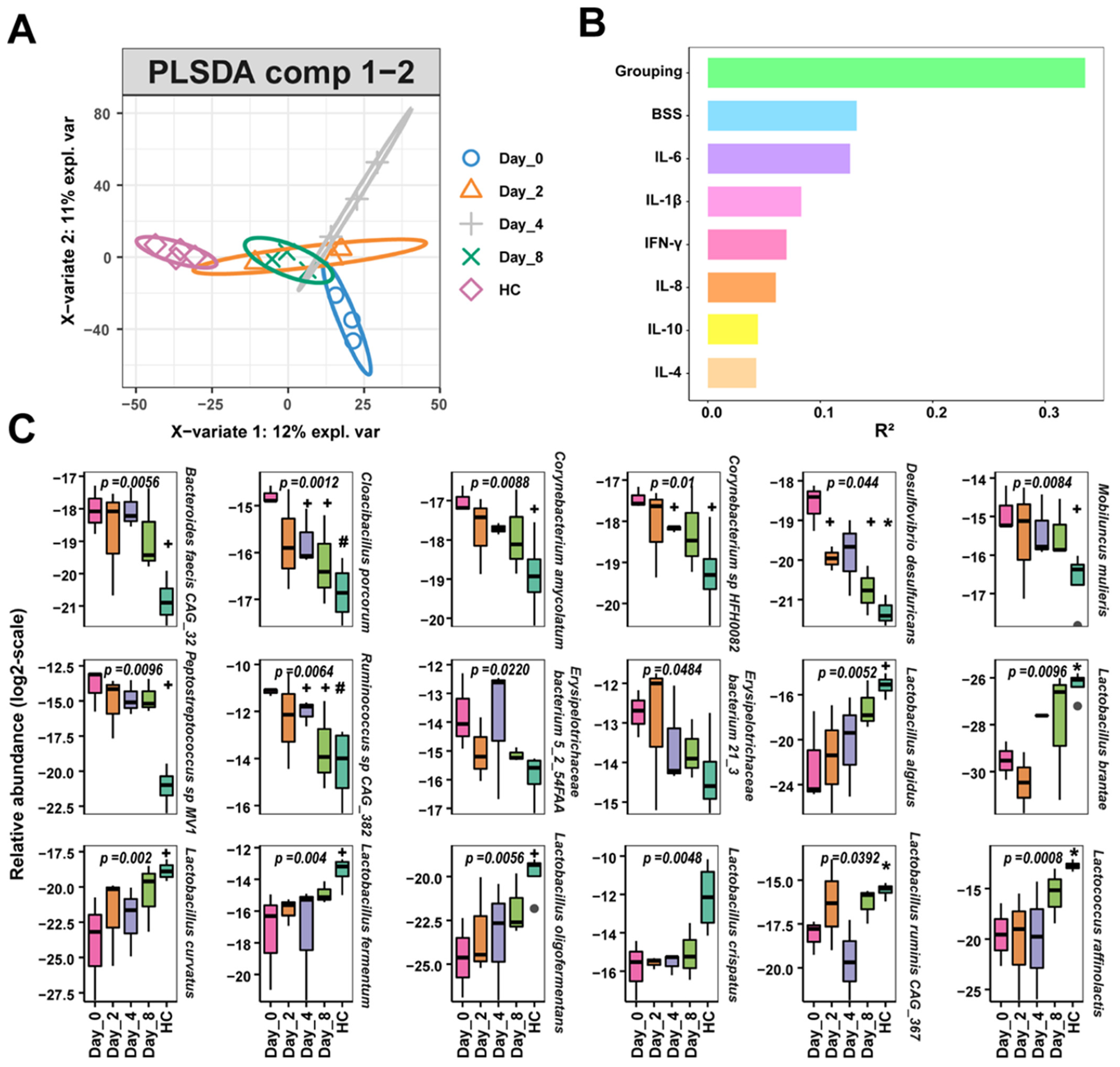
3.3. FMT Alters the Intestinal Bacteria Composition of Chronic Diarrhea Monkeys
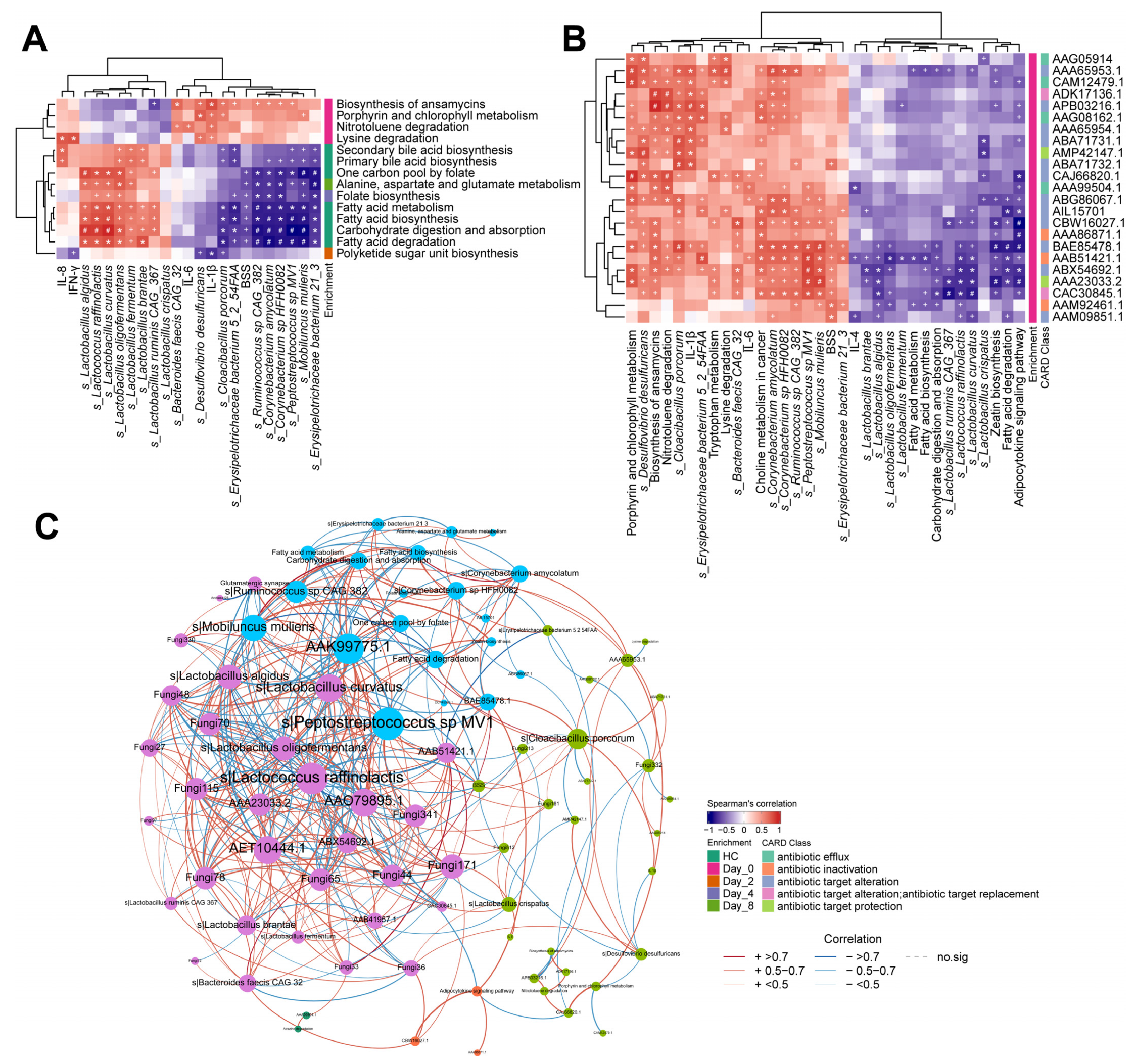
3.4. FMT Changes in Gut Bacteria Compositions Associated with Functional Changes
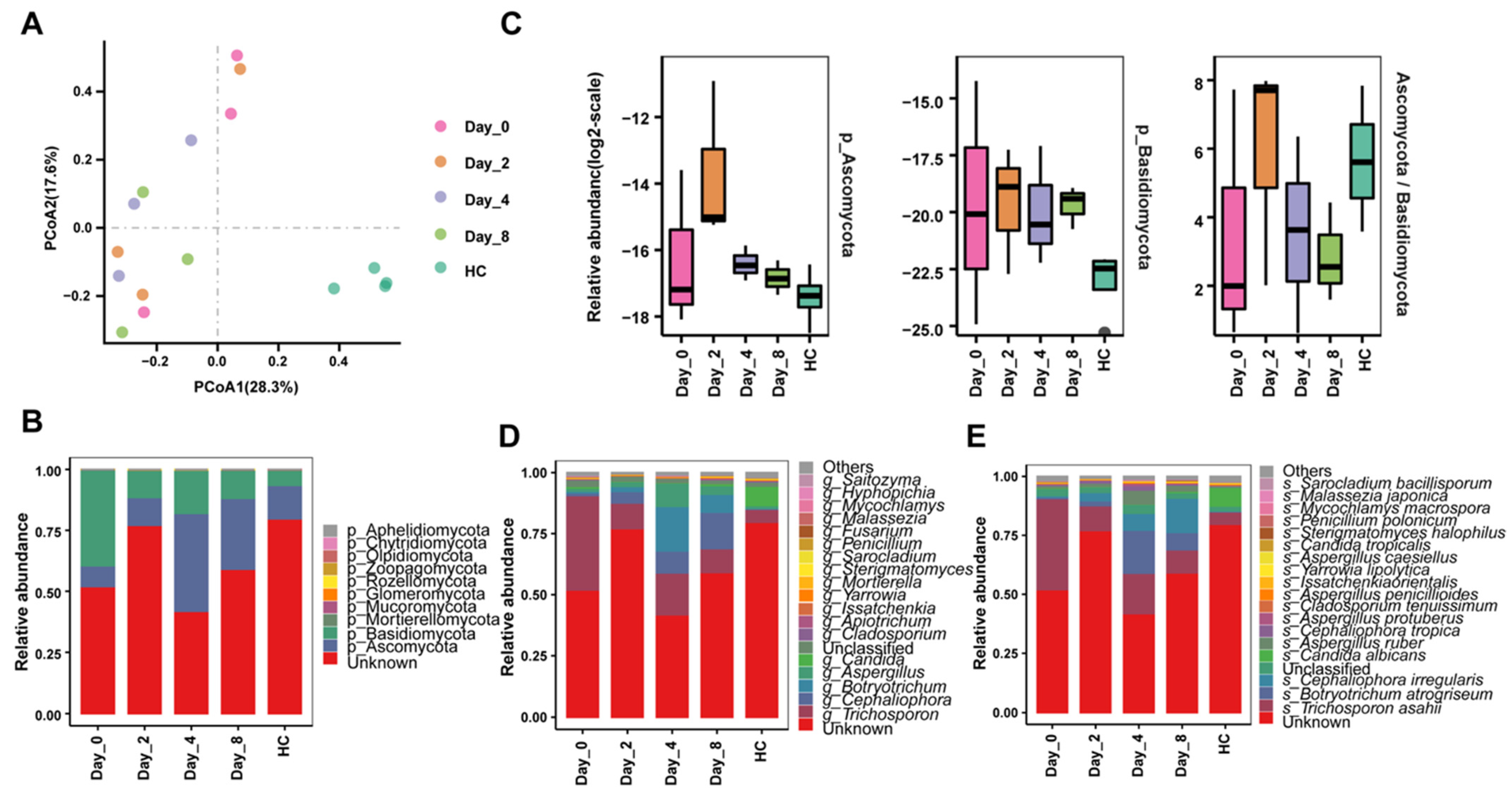
3.5. FMT Alters Gut Fungi of Chronic Diarrhea Monkeys in Relation to Treatment Responses
3.6. FMT Alters the Gut Archaea of Chronic Diarrhea Monkeys in Relation to Treatment Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, P.; Lee, H.N.; Rangan, V.; Ballou, S.; Lembo, J.; Katon, J.; McMahon, C.; Friedlander, D.; Iturrino, J.; Nee, J.; et al. Similarities in clinical and psychosocial characteristics of functional diarrhea and irritable bowel syndrome with diarrhea. Clin. Gastroenterol. H 2020, 18, 399–405. [Google Scholar] [CrossRef]
- Schiller, L.R. Evaluation of chronic diarrhea and irritable bowel syndrome with diarrhea in adults in the era of precision medicine. Am. J. Gastroenterol. 2018, 113, 660–669. [Google Scholar] [CrossRef]
- Yang, B.; Yue, Y.; Chen, Y.; Ding, M.F.; Li, B.W.; Wang, L.L.; Wang, Q.; Stanton, C.; Ross, R.P.; Zhao, J.X.; et al. Lactobacillus plantarum CCFM1143 alleviates chronic diarrhea via inflammation regulation and gut microbiota modulation: A double-blind, randomized, placebo-controlled study. Front. Immunol. 2021, 12, 746585. [Google Scholar] [CrossRef]
- Zollner-Schwetz, I.; Krause, R. Therapy of acute gastroenteritis: Role of antibiotics. Clin. Microbiol. Infect. 2015, 21, 744–749. [Google Scholar] [CrossRef] [Green Version]
- Dai, M.; Liu, Y.F.; Chen, W.; Buch, H.; Shan, Y.; Chang, L.H.; Bai, Y.; Shen, C.; Zhang, X.Y.; Huo, Y.F.; et al. Rescue fecal microbiota transplantation for antibiotic-associated diarrhea in critically ill patients. Crit. Care. 2019, 23, 324. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Ma, Y.; Yang, S.; Zhang, S.Y.; Liu, S.; Xiao, J.X.; Wang, Y.J.; Wang, W.; Yang, H.J.; Li, S.L.; et al. Gut microbiota-derived ursodeoxycholic acid from neonatal dairy calves improves intestinal homeostasis and colitis to attenuate extended-spectrum beta-lactamase-producing enteroaggregative Escherichia coli infection. Microbiome 2022, 10, 79. [Google Scholar] [CrossRef]
- Rhoades, N.; Barr, T.; Hendrickson, S.; Prongay, K.; Haertel, A.; Gill, L.; Garzel, L.; Whiteson, K.; Slifka, M.; Messaoudi, I. Maturation of the infant rhesus macaque gut microbiome and its role in the development of diarrheal disease. Genome Biol. 2019, 20, 173. [Google Scholar] [CrossRef] [Green Version]
- Khoruts, A.; Sadowsky, M.J. Understanding the mechanisms of faecal microbiota transplantation. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.M.; Cui, B.T.; He, X.X.; Nie, Y.Q.; Wu, K.C.; Fan, D.M. Microbiota transplantation: Concept, methodology and strategy for its modernization. Protein Cell. 2018, 9, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Gomez, D.E.; Arroyo, L.G.; Costa, M.C.; Viel, L.; Weese, J.S. Characterization of the fecal bacterial microbiota of healthy and diarrheic dairy calves. J. Vet. Intern. Med. 2017, 31, 928–939. [Google Scholar] [CrossRef]
- Li, S.S.; Zhu, A.; Benes, V.; Costea, P.I.; Hercog, R.; Hildebrand, F.; Huerta-Cepas, J.; Nieuwdorp, M.; Salojärvi, J.; Voigt, A.Y.; et al. Durable coexistence of donor and recipient strains after fecal microbiota transplantation. Science 2016, 352, 586–589. [Google Scholar] [CrossRef]
- Xiao, Y.D.; Angulo, M.T.; Lao, S.Y.; Weiss, S.T.; Liu, Y.Y. An ecological framework to understand the efficacy of fecal microbiota transplantation. Nat. Commun. 2020, 11, 3329. [Google Scholar] [CrossRef]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.W.M.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916. [Google Scholar] [CrossRef]
- Johnsen, P.H.; Hilpusch, F.; Valle, P.C.; Goll, R. The effect of fecal microbiota transplantation on IBS related quality of life and fatigue in moderate to severe non-constipated irritable bowel: Secondary endpoints of a double blind, randomized, placebo-controlled trial. EBioMedicine 2020, 51, 102562. [Google Scholar] [CrossRef] [Green Version]
- Johnsen, P.H.; Hilpüsch, F.; Cavanagh, J.P.; Leikanger, I.S.; Kolstad, C.; Valle, P.C.; Goll, R. Faecal microbiota transplantation versus placebo for moderate-to-severe irritable bowel syndrome: A double-blind, randomised, placebo-controlled, parallel-group, single-centre trial. Lancet Gastroenterol. 2018, 3, 17–24. [Google Scholar] [CrossRef]
- Aroniadis, O.C.; Brandt, L.J.; Oneto, C.; Feuerstadt, P.; Sherman, A.; Wolkoff, A.W.; Kassam, Z.; Sadovsky, R.G.; Elliott, R.J.; Budree, S.; et al. Faecal microbiota transplantation for diarrhoea-predominant irritable bowel syndrome: A double-blind, randomised, placebo-controlled trial. Lancet Gastroenterol. 2019, 4, 675–685. [Google Scholar] [CrossRef]
- Lam, S.; Bai, X.; Shkoporov, A.N.; Park, H.; Wu, X.J.; Lan, P.; Zuo, T. Roles of the gut virome and mycobiome in faecal microbiota transplantation. Lancet Gastroenterol. 2022, 7, 472–484. [Google Scholar] [CrossRef]
- Allegretti, J.R.; Mullish, B.H.; Kelly, C.; Fischer, M. The evolution of the use of faecal microbiota transplantation and emerging therapeutic indications. Lancet 2019, 394, 420–431. [Google Scholar] [CrossRef]
- Li, X.P.; Liang, S.S.; Xia, Z.K.; Qu, J.; Liu, H.; Liu, C.; Yang, H.M.; Wang, J.; Madsen, L.; Hou, Y.; et al. Establishment of a Macaca fascicularis gut microbiome gene catalog and comparison with the human, pig, and mouse gut microbiomes. Gigascience 2018, 7, giy100. [Google Scholar] [CrossRef]
- Yang, S.Z.; Liu, Y.; Yang, N.; Lan, Y.; Lan, W.Q.; Feng, J.Y.; Yue, B.S.; He, M.; Zhang, L.; Zhang, A.Y.; et al. The gut microbiome and antibiotic resistome of chronic diarrhea rhesus macaques (Macaca mulatta) and its similarity to the human gut microbiome. Microbiome 2022, 10, 29. [Google Scholar] [CrossRef]
- Wei, Z.Y.; Rao, J.H.; Tang, M.T.; Zhao, G.A.; Li, Q.C.; Wu, L.M.; Liu, S.Q.; Li, B.H.; Xiao, B.Q.; Liu, X.Y.; et al. Characterization of changes and driver microbes in gut microbiota during healthy aging using a captive monkey model. Genom. Proteom. Bioinform. 2021, 20, 350–365. [Google Scholar] [CrossRef]
- El-Salhy, M.; Winkel, R.; Casen, C.; Hausken, T.; Gilja, O.H.; Hatlebakk, J.G. Efficacy of fecal microbiota transplantation for patients with irritable bowel syndrome at 3 years after transplantation. Gastroenterology 2022, 163, 982–994. [Google Scholar] [CrossRef]
- Limon, J.J.; Skalski, J.H.; Underhill, D.M. Commensal fungi in health and disease. Cell Host. Microbe 2017, 22, 156–165. [Google Scholar] [CrossRef]
- Iliev, I.D.; Leonardi, I. Fungal dysbiosis: Immunity and interactions at mucosal barriers. Nat. Rev. Immunol. 2017, 17, 635–646. [Google Scholar] [CrossRef]
- van de Pol, J.A.; van Best, N.; Mbakwa, C.A.; Hijs, C.; Savelkoul, P.H.; Arts, I.C.W.; Hornef, M.W.; Mommers, M.; Penders, J. Gut colonization by methanogenic Archaea is associated with organic dairy consumption in children. Front. Microbiol. 2017, 8, 355. [Google Scholar] [CrossRef]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.-P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal microbiota dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Coker, O.O.; Nakatsu, G.; Dai, R.Z.; Wu, W.K.K.; Wong, S.H.; Ng, S.C.; Chan, F.K.L.; Sung, J.J.Y.; Yu, J. Enteric fungal microbiota dysbiosis and ecological alterations in colorectal cancer. Gut 2019, 68, 654–662. [Google Scholar] [CrossRef]
- Coker, O.O.; Wu, W.K.K.; Wong, S.H.; Sung, J.J.Y.; Yu, J. Altered gut Archaea composition and interaction with bacteria are associated with colorectal cancer. Gastroenterology 2020, 159, 1459–1470. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, R.; Zhu, H.; Ruan, J.; Qian, W.B.; Fang, X.D.; Shi, Z.B.; Li, Y.R.; Li, S.T.; Shan, G.; Kristiansen, K.; et al. De novo assembly of human genomes with massively parallel short read sequencing. Genome Res. 2010, 20, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Qin, N.; Yang, F.; Li, A.; Prifti, E.; Chen, Y.F.; Shao, L.; Guo, J.; Chatelier, E.L.; Yao, J.; Wu, L.J.; et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef]
- Zhu, W.; Lomsadze, A.; Borodovsky, M. Ab initio gene identification in metagenomic sequences. Nucleic Acids Res. 2010, 38, e132. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.T.; Li, W.Z. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [Green Version]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.H.; et al. CARD 2020, antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Xu, J.; Chen, N.; Song, Y.; Zhe, W.; Na, W.; Zhang, Y.F.; Ren, X.H.; Liu, Y.L. Alteration of fungal microbiota after 5-asa treatment in UC patients. Inflamm. Bowel. Dis. 2020, 26, 380–390. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, H.; Zhu, J.; Lin, X.G.; Feng, Y.Z. Divergent responses of methanogenic Archaeal communities in two rice cultivars to elevated ground-level O3. Environ. Pollut. 2016, 213, 127–134. [Google Scholar] [CrossRef]
- Liu, N.N.; Hu, H.F.; Ma, W.H.; Deng, Y.; Liu, Y.Q.; Hao, B.H.; Zhang, X.Y.; Dimitrov, D.; Feng, X.J.; Wang, Z.H. Contrasting biogeographic patterns of bacterial and Archaeal diversity in the top- and subsoils of temperate grasslands. mSystems 2019, 4, e00566-19. [Google Scholar] [CrossRef] [Green Version]
- Leonardi, I.; Paramsothy, S.; Doron, I.; Semon, A.; Kaakoush, N.O.; Clemente, J.C.; Faith, J.J.; Borody, T.J.; Mitchell, H.M.; Colombel, J.-F.; et al. Fungal trans-kingdom dynamics linked to responsiveness to fecal microbiota transplantation (FMT) therapy in ulcerative colitis. Cell Host. Microbe 2020, 27, 823–829. [Google Scholar] [CrossRef]
- Singh, P.; Mitsuhashi, S.; Ballou, S.; Rangan, V.; Sommers, T.; Cheng, V.; Iturrino-Moreda, J.; Friedlander, D.; Nee, J.; Lembo, A. Demographic and dietary associations of chronic diarrhea in a representative sample of adults in the United States. Am. J. Gastroenterol. 2018, 113, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ma, C.; Zhao, F.; Chen, P.; Liu, Y.H.; Sun, Z.H.; Cui, L.H.; Kwok, L.-Y.; Zhang, H.P. Adjunctive treatment with probiotics partially alleviates symptoms and reduces inflammation in patients with irritable bowel syndrome. Eur. J. Nutr. 2021, 60, 2553–2565. [Google Scholar] [CrossRef] [PubMed]
- Madempudi, R.S.; Ahire, J.J.; Neelamraju, J.; Tripathi, A.; Nanal, S. Randomized clinical trial: The effect of probiotic Bacillus coagulans Unique IS2 vs. placebo on the symptoms management of irritable bowel syndrome in adults. Sci. Rep. 2019, 9, 12210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennet, S.M.; Polster, A.; Tornblom, H.; Isaksson, S.; Capronnier, S.; Tessier, A.; Nevé, B.L.; Simrén, M.; Öhman, L. Global cytokine profiles and association with clinical characteristics in patients with irritable bowel syndrome. Am. J. Gastroenterol. 2016, 111, 1165–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananthakrishnan, A.N.; Luo, C.; Yajnik, V.; Khalili, H.; Garber, J.J.; Stevens, B.W.; Cleland, T.; Xavier, R.J. Gut microbiome function predicts response to anti-integrin biologic therapy in inflammatory bowel diseases. Cell Host. Microbe 2017, 21, 603–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linninge, C.; Xu, J.; Bahl, M.I.; Ahrné, S.; Molin, G. Lactobacillus fermentum and Lactobacillus plantarum increased gut microbiota diversity and functionality, and mitigated Enterobacteriaceae, in a mouse model. Benef. Microbes 2019, 10, 413–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, G.; Burton, J. Use of Lactobacillus to prevent infection by pathogenic bacteria. Microbes Infect. 2002, 4, 319–324. [Google Scholar] [CrossRef]
- Borisa, S.; Barbés, C. Role played by lactobacilli in controlling the population of vaginal pathogens. Microbes Infect. 2000, 2, 543–546. [Google Scholar] [CrossRef]
- Spinler, J.K.; Taweechotipatr, M.; Rognerud, C.L.; Ou, C.N.; Tumwasorn, S.; Versalovic, J. Human-derived probiotic Lactobacillus reuteri demonstrate antimicrobial activities targeting diverse enteric bacterial pathogens. Anaerobe 2008, 14, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; d’Aigle, J.; Atadja, L.; Quaicoe, V.; Honarpisheh, P.; Ganesh, B.P.; Hassan, A.; Graf, J.; Petrosino, J.; Putluri, N.; et al. Gut microbiota-derived short-chain fatty acids promote poststroke recovery in aged mice. Circ. Res. 2020, 127, 453–465. [Google Scholar] [CrossRef]
- Su, L.; Ma, F.; An, Z.; Ji, X.Y.; Zhang, P.; Yue, Q.L.; Zhao, C.; Sun, X.; Li, K.L.; Li, B.J.; et al. The metabolites of Lactobacillus fermentum F-B9-1 relieved dextran sulfate sodium-induced experimental ulcerative colitis in mice. Front. Microbiol. 2022, 13, 865925. [Google Scholar] [CrossRef] [PubMed]
- Castro-Mejia, J.L.; O’Ferrall, S.; Krych, L.; O’Mahony, E.; Namusoke, H.; Lanyero, B.; Kot, W.; Nabukeera-Barungi, N.; Michaelsen, K.F.; Mølgaard, C.; et al. Restitution of gut microbiota in Ugandan children administered with probiotics (Lactobacillus rhamnosus GG and Bifidobacterium animalis subsp. lactis BB-12) during treatment for severe acute malnutrition. Gut Microbes 2020, 11, 855–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaakoush, N.O. Insights into the Role of Erysipelotrichaceae in the Human Host. Front. Cell Infect. Microbiol. 2015, 5, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.H.; Ma, W.; Wang, D.D.; Cao, Y.; Mallick, H.; Gerbaba, T.K.; Lloyd-Price, J.; Abu-Ali, G.; Hall, A.B.; Sikavi, D.; et al. Association between sulfur-metabolizing bacterial communities in stool and risk of distal colorectal cancer in men. Gastroenterology 2020, 158, 1313–1325. [Google Scholar] [CrossRef] [Green Version]
- Looft, T.; Levine, U.Y.; Stanton, T.B. Cloacibacillus porcorum sp. nov., a mucin-degrading bacterium from the swine intestinal tract and emended description of the genus Cloacibacillus. Int. J. Syst. Evol. Microbiol. 2013, 63, 1960–1966. [Google Scholar] [CrossRef] [PubMed]
- Pullan, R.D.; Thomas, G.A.; Rhodes, M.; Newcombe, R.G.; Williams, G.T.; Allen, A.; Rhodes, J. Thickness of adherent mucus gel on colonic mucosa in humans and its relevance to colitis. Gut 1994, 35, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Bromberg, J.S.; Hittle, L.; Xiong, Y.; Saxena, V.; Smyth, E.M.; Li, L.; Zhang, T.S.; Wagner, C.; Fricke, W.F.; Simon, T.; et al. Gut microbiota-dependent modulation of innate immunity and lymph node remodeling affects cardiac allograft outcomes. JCI Insight 2018, 5, e142528. [Google Scholar] [CrossRef]
- Abubucker, S.; Segata, N.; Goll, J.; Schubert, A.M.; Izard, J.; Cantarel, B.L.; Rodriguez-Mueller, B.; Zucker, J.; Thiagarajan, M.; Henrissat, B.; et al. Metabolic reconstruction for metagenomic data and its application to the human microbiome. PLoS Comput. Biol. 2012, 8, e1002358. [Google Scholar] [CrossRef] [Green Version]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Kim, H. The roles of glutamine in the intestine and its implication in intestinal diseases. Int. J. Mol. Sci. 2017, 18, 1051. [Google Scholar] [CrossRef]
- Rao, R.; Samak, G. Role of glutamine in protection of intestinal epithelial tight junctions. J. Epithel. Biol. Pharmacol. 2012, 5, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, Y.; Reshef, L.; Maya, M.; Cohen, D.; Gophna, U.; Muhsen, K. Socioeconomic disparities and household crowding in association with the fecal microbiome of school-age children. NPJ Biofilms Microbiomes 2022, 8, 10. [Google Scholar] [CrossRef]
- Shi, Z.J.; Zhao, R.X.; Wan, J.J.; Li, B.; Shen, Y.; Zhang, S.C.; Luo, G. Metagenomic analysis reveals the fate of antibiotic resistance genes in two-stage and one-stage anaerobic digestion of waste activated sludge. J. Hazard. Mater. 2021, 406, 124595. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.C.; Wang, L.M.; Ke, S.L.; Gálvez, J.A.V.; Pollock, N.R.; Barrett, C.; Sprague, R.; Daugherty, K.; Xu, H.; Lin, Q.Y.; et al. Fecal mycobiota combined with host immune factors distinguish Clostridioides difficile infection from asymptomatic carriage. Gastroenterology 2021, 160, 2328–2339. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Oliveira, C.; Rodrigues, F.; Goncalves, S.M.; Goldman, G.H.; Carvalho, A.; Cunha, C. The cell biology of the trichosporon-host interaction. Front. Cell Infect. Microbiol. 2017, 7, 118. [Google Scholar] [CrossRef] [Green Version]
- Ali, G.A.; Husain, A.; Salah, H.; Goravey, W. Trichosporon asahii fungemia and COVID-19 co-infection: An emerging fungal pathogen; case report and review of the literature. IDCases 2021, 25, e01244. [Google Scholar] [CrossRef] [PubMed]
- Blais Lecours, P.; Marsolais, D.; Cormier, Y.; Berberi, M.; Haché, C.; Bourdages, R.; Duchaine, C. Increased prevalence of Methanosphaera stadtmanae in inflammatory bowel diseases. PLoS ONE 2014, 9, e87734. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, P.; Gao, J.; Liang, L.; Cui, B.; Hu, Q.; Zhou, W.; Li, B.; Liu, Y.; Chen, T.; Rao, J.; et al. Fecal Microbiota Transplantation Could Improve Chronic Diarrhea in Cynomolgus Monkey by Alleviating Inflammation and Modulating Gut Microbiota. Biomedicines 2022, 10, 3016. https://doi.org/10.3390/biomedicines10123016
Tian P, Gao J, Liang L, Cui B, Hu Q, Zhou W, Li B, Liu Y, Chen T, Rao J, et al. Fecal Microbiota Transplantation Could Improve Chronic Diarrhea in Cynomolgus Monkey by Alleviating Inflammation and Modulating Gut Microbiota. Biomedicines. 2022; 10(12):3016. https://doi.org/10.3390/biomedicines10123016
Chicago/Turabian StyleTian, Puyuan, Jiangmei Gao, Lifeng Liang, Bota Cui, Qiming Hu, Wenfeng Zhou, Bihai Li, Yiyan Liu, Tingtao Chen, Junhua Rao, and et al. 2022. "Fecal Microbiota Transplantation Could Improve Chronic Diarrhea in Cynomolgus Monkey by Alleviating Inflammation and Modulating Gut Microbiota" Biomedicines 10, no. 12: 3016. https://doi.org/10.3390/biomedicines10123016
APA StyleTian, P., Gao, J., Liang, L., Cui, B., Hu, Q., Zhou, W., Li, B., Liu, Y., Chen, T., Rao, J., & Wei, H. (2022). Fecal Microbiota Transplantation Could Improve Chronic Diarrhea in Cynomolgus Monkey by Alleviating Inflammation and Modulating Gut Microbiota. Biomedicines, 10(12), 3016. https://doi.org/10.3390/biomedicines10123016