

Inactivation of Yes-Associated Protein Mediates Trophoblast Dysfunction: A New Mechanism of Pregnancy Loss Associated with Anti-Phospholipid Antibodies?
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibody Preparation
2.2. Cell Culture and Treatments
2.3. Plasmid and siRNA Transfections
2.4. RNA Isolation, cDNA Synthesis, and Real-Time Quantitative PCR
2.5. Immunoblotting
2.6. Immunofluorescence Staining
2.7. Cell Apoptosis and Cell Cycle Assays
2.8. Cell Migration Assay
2.9. Tube Formation Assay
2.10. Statistical Analysis
3. Results
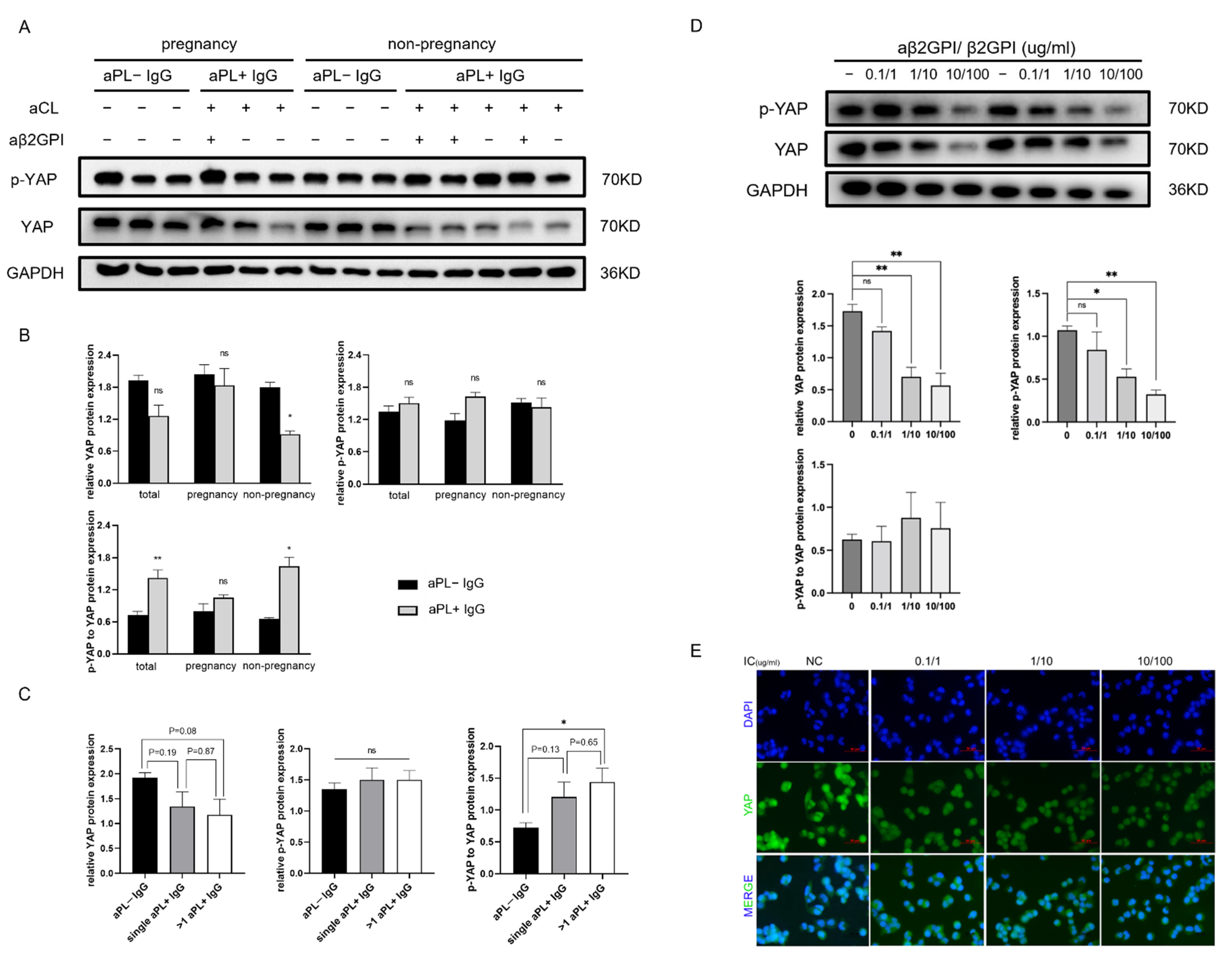
3.1. aPL+ IgG Affects YAP Expression but Does Not Promote the Phosphorylation of YAP Protein in HTR-8 Cells
3.2. Anti-β2GPI-IgG Monoclonal Antibody and Human β2GPI Complex Function in a Dose-Dependent Manner to Reduce YAP Expression and Nuclear Localization in HTR-8 Cells
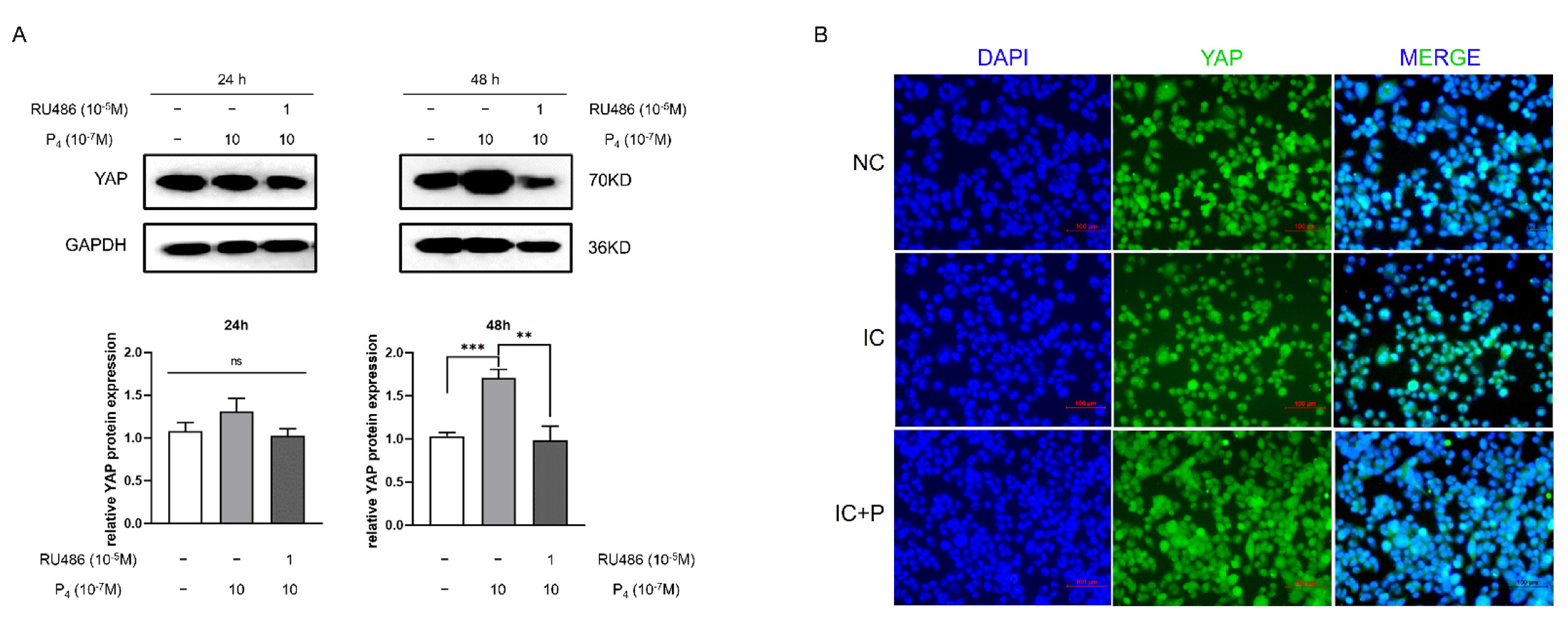
3.3. Progesterone Enhances the Expression of YAP and Is Partly Reversed by RU486 in HTR-8 Cells
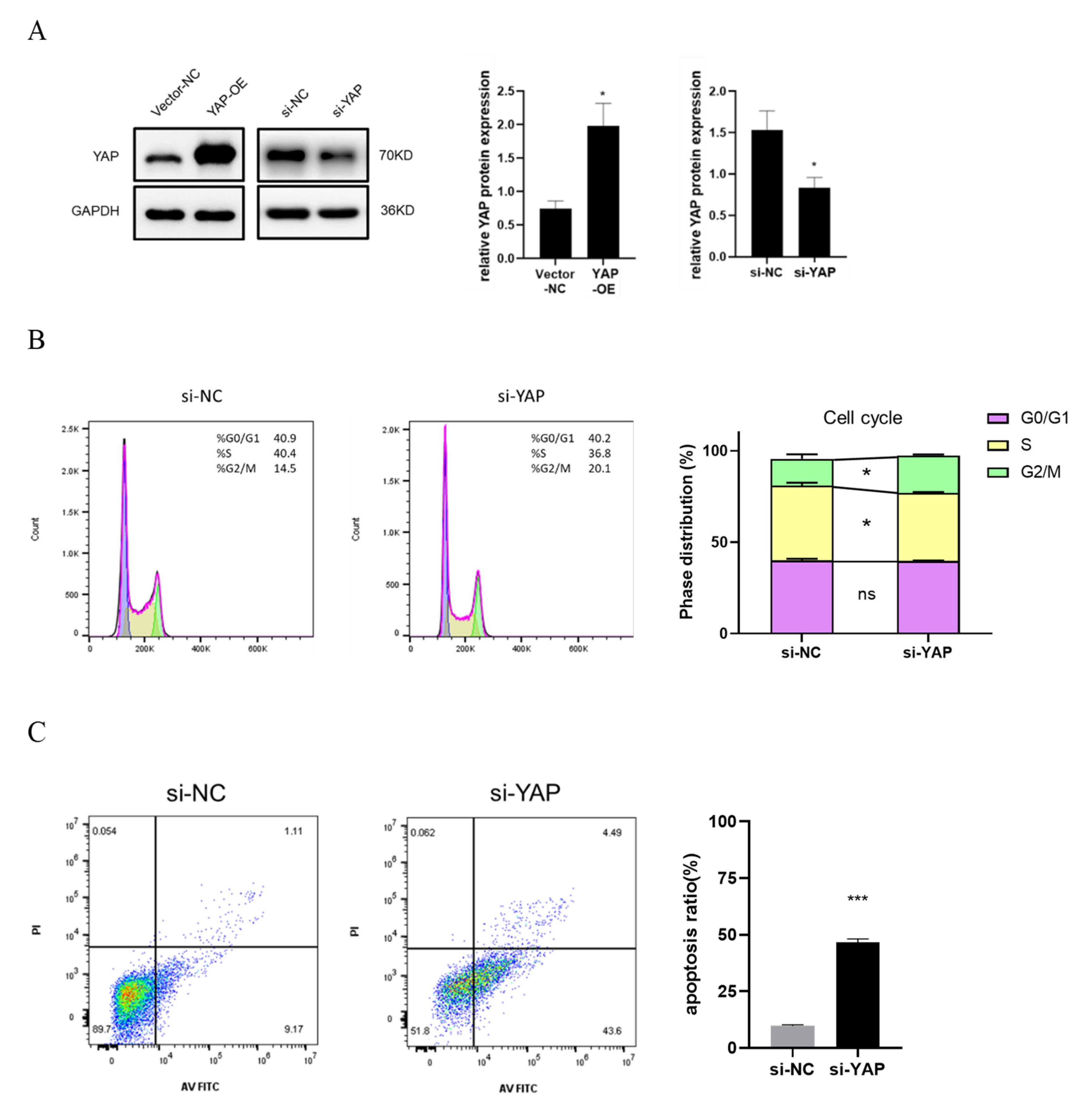
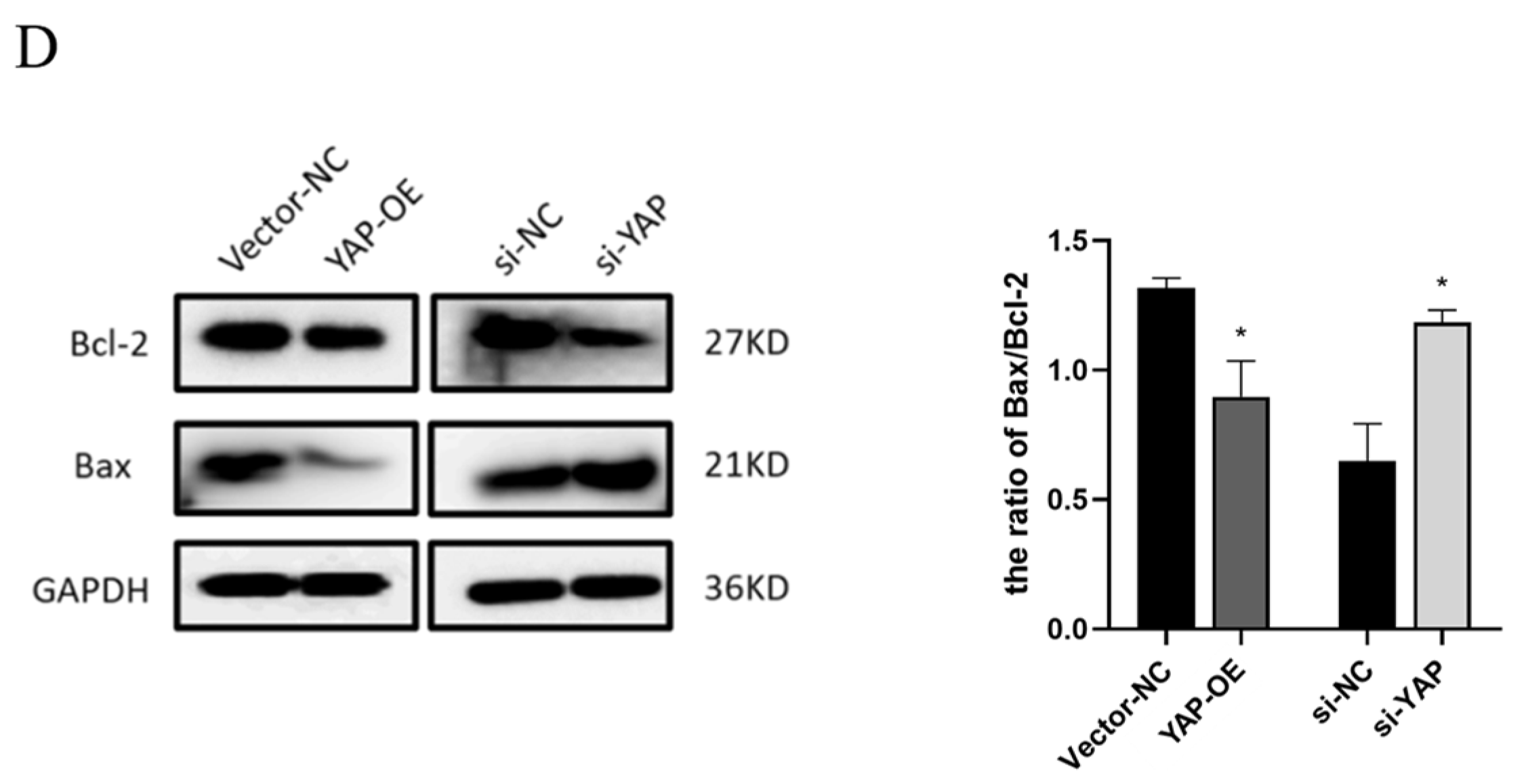
3.4. YAP Depletion Induces Cell Cycle Arrest and Cell Apoptosis of Trophoblast
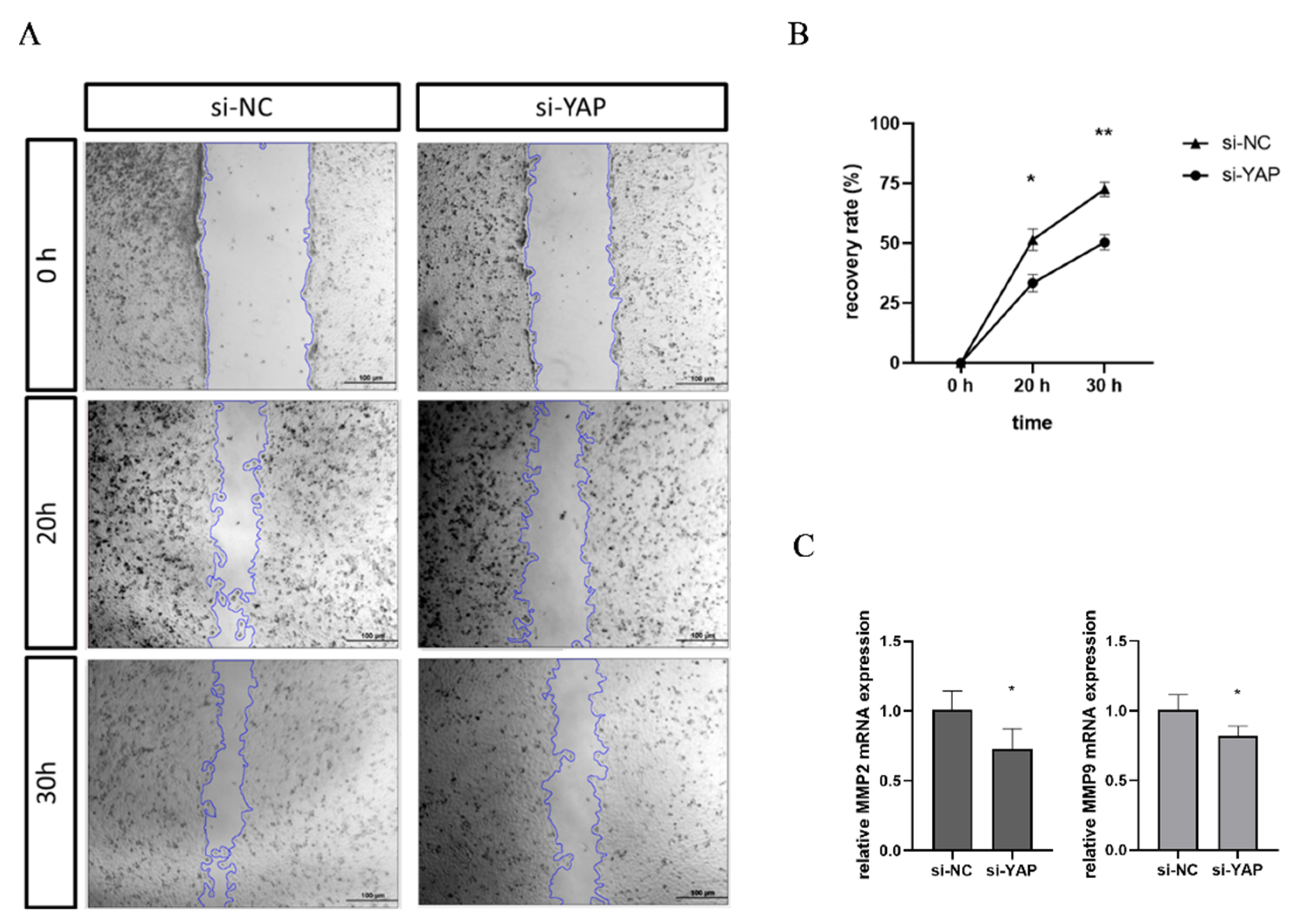
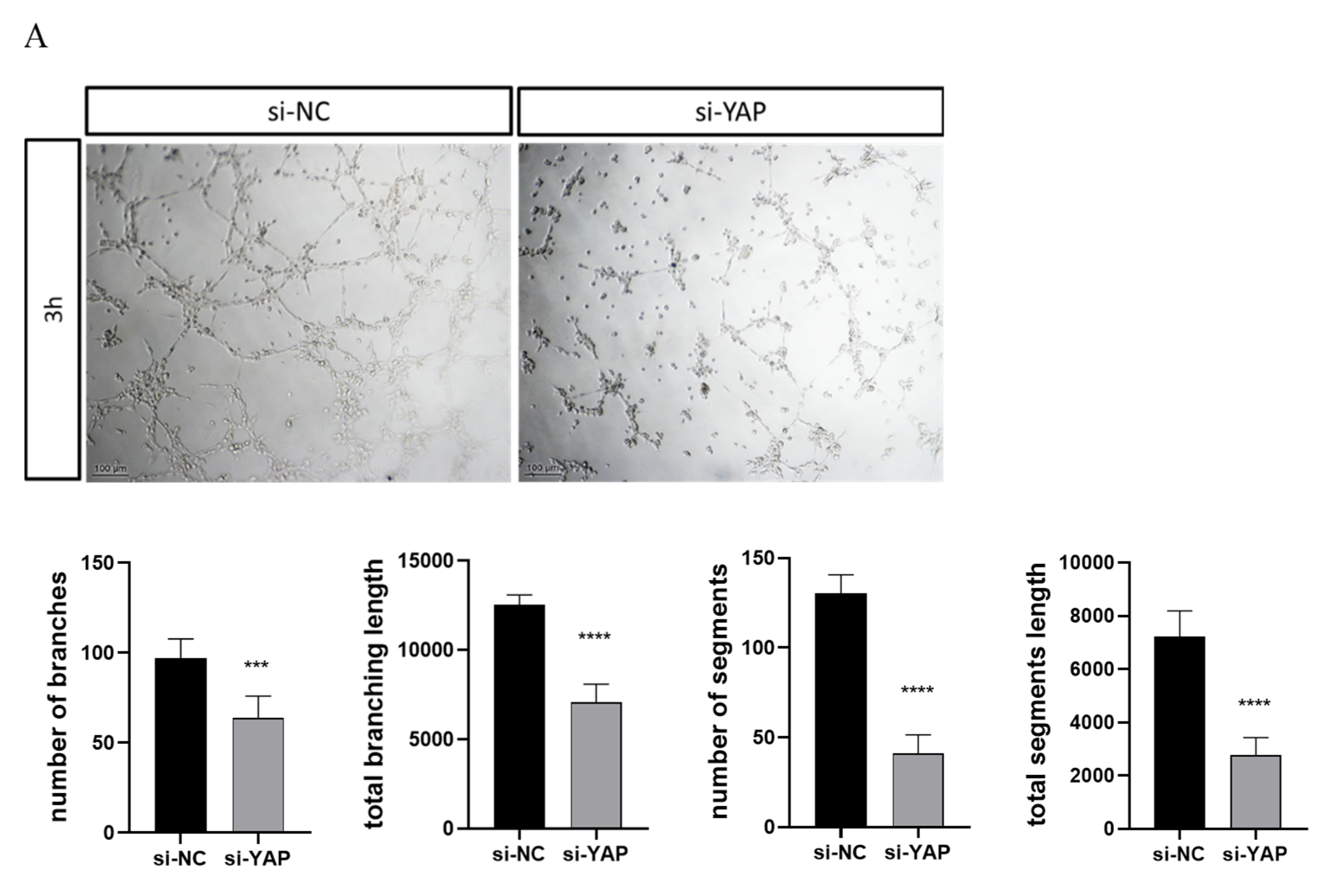
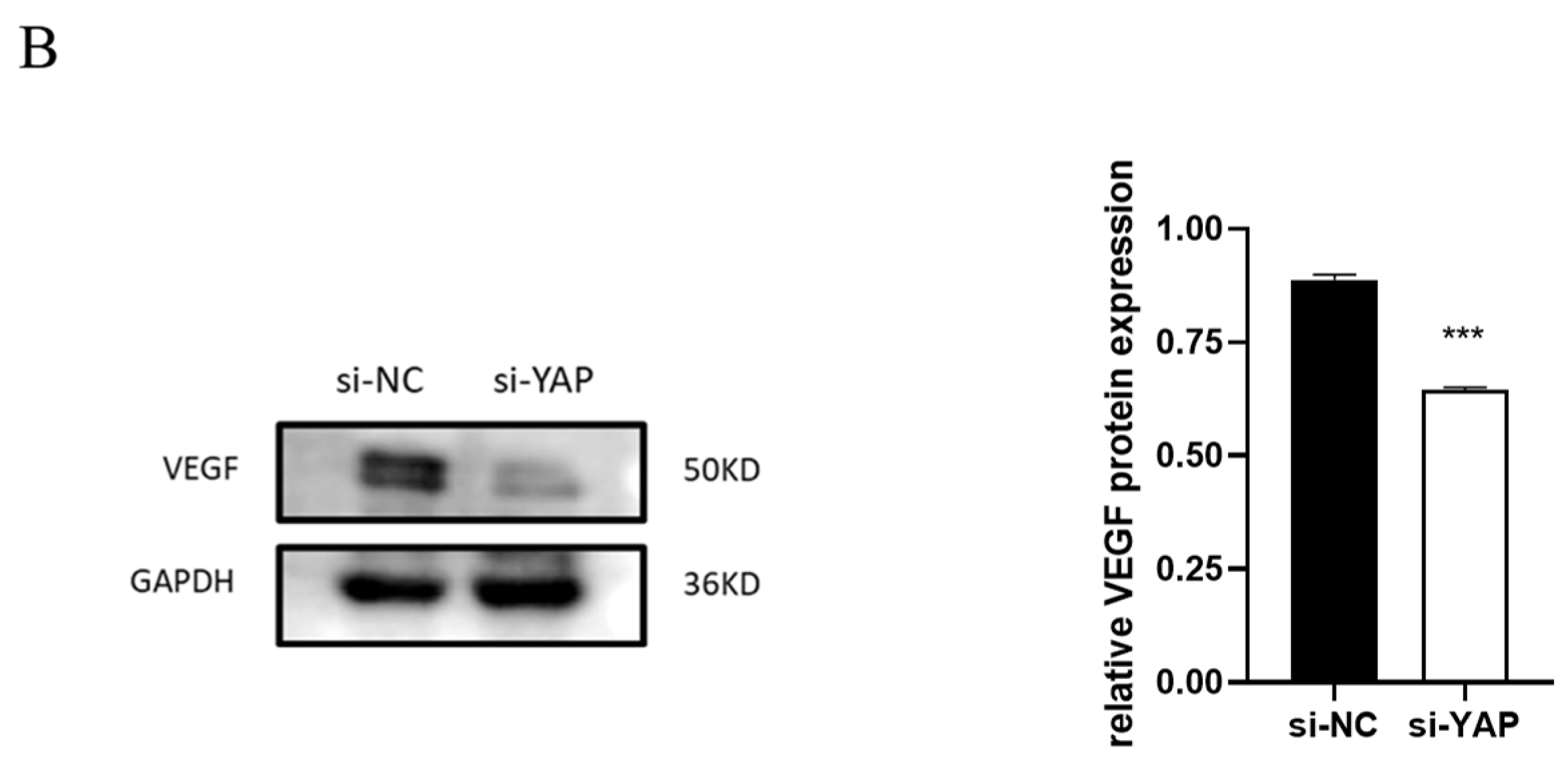
3.5. Downregulated YAP in Trophoblast Impairs Cell Migration and Tube Formation
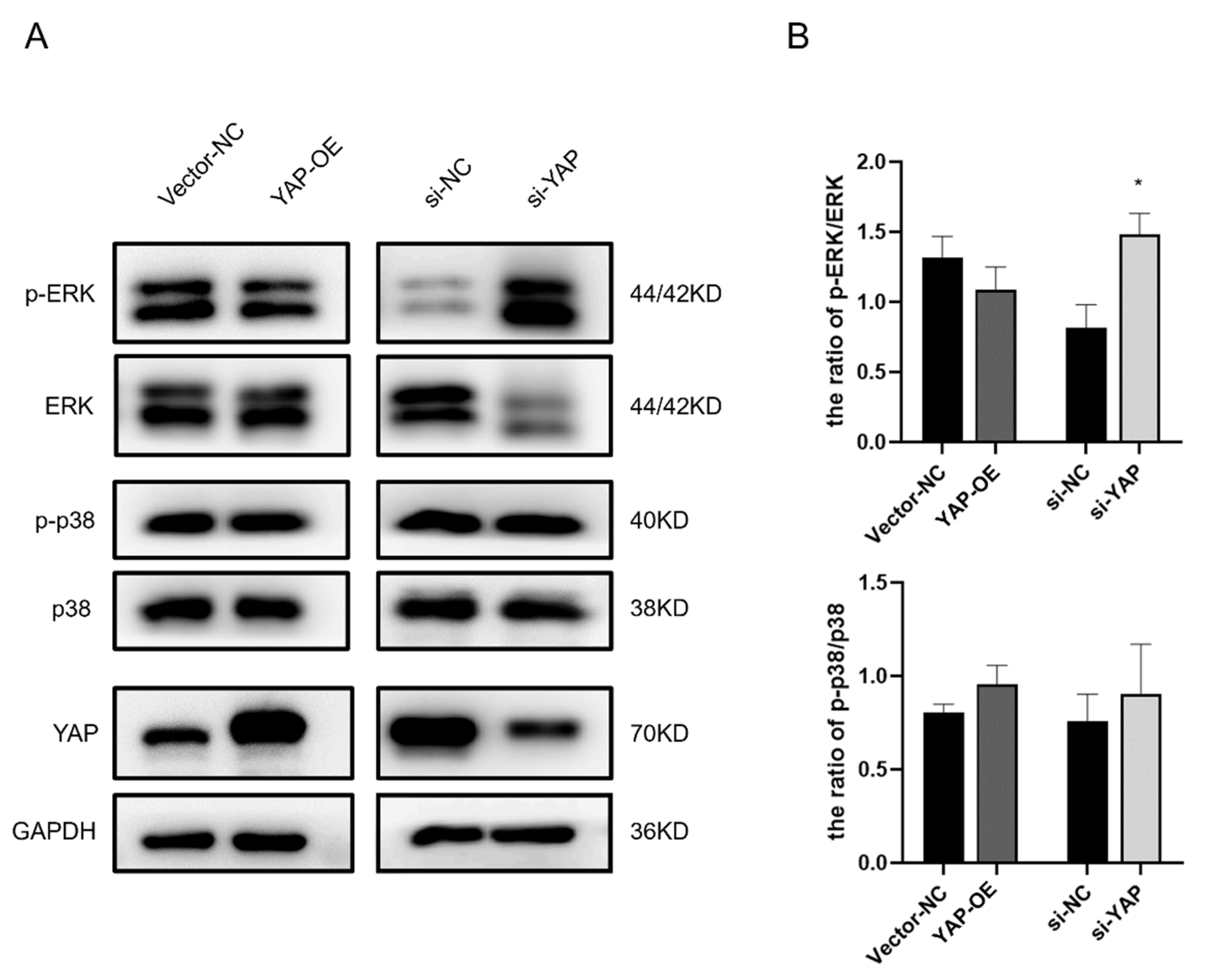
3.6. YAP Knockdown Results in the Activation of ERK1/2 in HTR-8 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nalli, C.; Somma, V.; Andreoli, L.; Buttner, T.; Schierack, P.; Mahler, M.; Roggenbuck, D.; Tincani, A. Anti-Phospholipid IgG Antibodies Detected by Line Immunoassay Differentiate Patients with Anti-Phospholipid Syndrome and Other Autoimmune Diseases. Auto Immun. Highlights 2018, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Saccone, G.; Berghella, V.; Maruotti, G.M.; Ghi, T.; Rizzo, G.; Simonazzi, G.; Rizzo, N.; Facchinetti, F.; Dall’Asta, A.; Visentin, S.; et al. Antiphospholipid Antibody Profile Based Obstetric Outcomes of Primary Antiphospholipid Syndrome: The PREGNANTS Study. Am. J. Obs. Gynecol. 2017, 216, 525.e1–525.e12. [Google Scholar] [CrossRef]
- Garcia, D.; Erkan, D. Diagnosis and Management of the Antiphospholipid Syndrome. N. Engl. J. Med. 2018, 378, 2010–2021. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.A.; Silver, R.M.; Branch, D.W. Do Antiphospholipid Antibodies Cause Preeclampsia and HELLP Syndrome? Curr. Rheumatol. Rep. 2007, 9, 219–225. [Google Scholar] [CrossRef]
- Santos, T.D.S.; Ieque, A.L.; de Carvalho, H.C.; Sell, A.M.; Lonardoni, M.V.C.; Demarchi, I.G.; de Lima Neto, Q.A.; Teixeira, J.J.V. Antiphospholipid Syndrome and Recurrent Miscarriage: A Systematic Review and Meta-Analysis. J. Reprod. Immunol. 2017, 123, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Opatrny, L.; David, M.; Kahn, S.R.; Shrier, I.; Rey, E. Association between Antiphospholipid Antibodies and Recurrent Fetal Loss in Women without Autoimmune Disease: A Metaanalysis. J. Rheumatol. 2006, 33, 2214–2221. [Google Scholar] [PubMed]
- Ofer-Shiber, S.; Molad, Y. Frequency of Vascular and Pregnancy Morbidity in Patients with Low vs. Moderate-to-High Titers of Antiphospholipid Antibodies. Blood Coagul. Fibrinolysis 2015, 26, 261–266. [Google Scholar] [CrossRef] [PubMed]
- De Carolis, S.; Tabacco, S.; Rizzo, F.; Giannini, A.; Botta, A.; Salvi, S.; Garufi, C.; Panici, P.B.; Lanzone, A. Antiphospholipid Syndrome: An Update on Risk Factors for Pregnancy Outcome. Autoimmun. Rev. 2018, 17, 956–966. [Google Scholar] [CrossRef] [PubMed]
- Viall, C.A.; Chamley, L.W. Histopathology in the Placentae of Women with Antiphospholipid Antibodies: A Systematic Review of the Literature. Autoimmun. Rev. 2015, 14, 446–471. [Google Scholar] [CrossRef]
- Ripoll, V.M.; Pregnolato, F.; Mazza, S.; Bodio, C.; Grossi, C.; McDonnell, T.; Pericleous, C.; Meroni, P.L.; Isenberg, D.A.; Rahman, A.; et al. Gene Expression Profiling Identifies Distinct Molecular Signatures in Thrombotic and Obstetric Antiphospholipid Syndrome. J. Autoimmun. 2018, 93, 114–123. [Google Scholar] [CrossRef]
- Poulton, K.; Ripoll, V.M.; Pericleous, C.; Meroni, P.L.; Gerosa, M.; Ioannou, Y.; Rahman, A.; Giles, I.P. Purified IgG from Patients with Obstetric but Not IgG from Non-Obstetric Antiphospholipid Syndrome Inhibit Trophoblast Invasion. Am. J. Reprod. Immunol. 2015, 73, 390–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Han, J.; Zhang, R.; Tang, Z.; Yi, G.; Gong, W.; Wan, L.; Hu, Q.; Teng, J.; Liu, H.; et al. Characteristics of Purified Anti-Beta2GPI IgG N-Glycosylation Associate with Thrombotic, Obstetric and Catastrophic Antiphospholipid Syndrome. Rheumatology (Oxford) 2022, 61, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, S.; McCrae, K.R. Diagnosis and Management of the Antiphospholipid Syndrome. Blood Rev. 2017, 31, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Fierro, J.J.; Velasquez, M.; Cadavid, A.P.; de Leeuw, K. Effects of Anti-Beta 2-Glycoprotein 1 Antibodies and Its Association with Pregnancy-Related Morbidity in Antiphospholipid Syndrome. Am. J. Reprod. Immunol. 2021, 87, e13509. [Google Scholar] [CrossRef]
- Ulrich, V.; Gelber, S.E.; Vukelic, M.; Sacharidou, A.; Herz, J.; Urbanus, R.T.; de Groot, P.G.; Natale, D.R.; Harihara, A.; Redecha, P.; et al. ApoE Receptor 2 Mediation of Trophoblast Dysfunction and Pregnancy Complications Induced by Antiphospholipid Antibodies in Mice. Arthritis Rheumatol. 2016, 68, 730–739. [Google Scholar] [CrossRef] [Green Version]
- Gysler, S.M.; Mulla, M.J.; Guerra, M.; Brosens, J.J.; Salmon, J.E.; Chamley, L.W.; Abrahams, V.M. Antiphospholipid Antibody-Induced MiR-146a-3p Drives Trophoblast Interleukin-8 Secretion through Activation of Toll-like Receptor 8. Mol. Hum. Reprod. 2016, 22, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Mulla, M.J.; Weel, I.C.; Potter, J.A.; Gysler, S.M.; Salmon, J.E.; Peracoli, M.T.S.; Rothlin, C.V.; Chamley, L.W.; Abrahams, V.M. Antiphospholipid Antibodies Inhibit Trophoblast Toll-Like Receptor and Inflammasome Negative Regulators. Arthritis Rheumatol. 2018, 70, 891–902. [Google Scholar] [CrossRef] [Green Version]
- Krivokuća, M.J.; Abu Rabi, T.; Stefanoska, I.; Vrzić-Petronijević, S.; Petronijević, M.; Vićovac, L. Immunoglobulins from sera of APS patients bind HTR-8/SVneo trophoblast cell line and reduce additional mediators of cell invasion. Reprod. Biol. 2017, 17, 389–395. [Google Scholar] [CrossRef]
- Marchetti, T.; Ruffatti, A.; Wuillemin, C.; de Moerloose, P.; Cohen, M. Hydroxychloroquine Restores Trophoblast Fusion Affected by Antiphospholipid Antibodies. J. Thromb. Haemost. 2014, 12, 910–920. [Google Scholar] [CrossRef]
- Di Simone, N.; D’Ippolito, S.; Marana, R.; di Nicuolo, F.; Castellani, R.; Pierangeli, S.S.; Chen, P.; Tersigni, C.; Scambia, G.; Meroni, P.L. Antiphospholipid Antibodies Affect Human Endometrial Angiogenesis: Protective Effect of a Synthetic Peptide (TIFI) Mimicking the Phospholipid Binding Site of Beta(2) Glycoprotein I. Am. J. Reprod. Immunol. 2013, 70, 299–308. [Google Scholar] [CrossRef]
- Tong, M.; Viall, C.A.; Chamley, L.W. Antiphospholipid Antibodies and the Placenta: A Systematic Review of Their in Vitro Effects and Modulation by Treatment. Hum. Reprod. Update 2015, 21, 97–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meroni, P.L.; Borghi, M.O.; Raschi, E.; Tedesco, F. Pathogenesis of Antiphospholipid Syndrome: Understanding the Antibodies. Nat. Rev. Rheumatol. 2011, 7, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Chen, A.C.H.; Tian, S.; Fong, S.W.; Lee, K.C.; Zhang, J.; Ng, E.H.Y.; Lee, K.F.; Yeung, W.S.B.; Lee, Y.L. Human Embryonic Stem Cell-Derived Blastocyst-like Spheroids Resemble Human Trophectoderm during Early Implantation Process. Fertil. Steril. 2020, 114, 653–664.e6. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, N.; Inoue, K.; Adachi, K.; Kiyonari, H.; Ota, M.; Ralston, A.; Yabuta, N.; Hirahara, S.; Stephenson, R.O.; Ogonuki, N.; et al. The Hippo Signaling Pathway Components Lats and Yap Pattern Tead4 Activity to Distinguish Mouse Trophectoderm from Inner Cell Mass. Dev. Cell 2009, 16, 398–410. [Google Scholar] [CrossRef] [Green Version]
- Rayon, T.; Menchero, S.; Nieto, A.; Xenopoulos, P.; Crespo, M.; Cockburn, K.; Canon, S.; Sasaki, H.; Hadjantonakis, A.K.; de la Pompa, J.L.; et al. Notch and Hippo Converge on Cdx2 to Specify the Trophectoderm Lineage in the Mouse Blastocyst. Dev. Cell 2014, 30, 410–422. [Google Scholar] [CrossRef] [Green Version]
- Valis, K.; Novak, P. Targeting ERK-Hippo Interplay in Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 3236. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Zhang, R.; Lu, M.; Liu, S.; Baba, H.A.; Gerken, G.; Wedemeyer, H.; Broering, R. Hippo Pathway Counter-Regulates Innate Immunity in Hepatitis B Virus Infection. Front. Immunol. 2021, 12, 684424. [Google Scholar] [CrossRef]
- Yu, F.X.; Zhao, B.; Guan, K.L. Hippo Pathway in Organ Size Control, Tissue Homeostasis, and Cancer. Cell 2015, 163, 811–828. [Google Scholar] [CrossRef] [Green Version]
- Koo, J.H.; Guan, K.L. Interplay between YAP/TAZ and Metabolism. Cell Metab. 2018, 28, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Anani, S.; Bhat, S.; Honma-Yamanaka, N.; Krawchuk, D.; Yamanaka, Y. Initiation of Hippo Signaling Is Linked to Polarity Rather than to Cell Position in the Pre-Implantation Mouse Embryo. Development 2014, 141, 2813–2824. [Google Scholar] [CrossRef]
- Meng, Z.; Moroishi, T.; Guan, K.L. Mechanisms of Hippo Pathway Regulation. Genes Dev. 2016, 30, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Saha, B.; Ganguly, A.; Home, P.; Bhattacharya, B.; Ray, S.; Ghosh, A.; Rumi, M.A.K.; Marsh, C.; French, V.A.; Gunewardena, S.; et al. TEAD4 Ensures Postimplantation Development by Promoting Trophoblast Self-Renewal: An Implication in Early Human Pregnancy Loss. Proc. Natl. Acad. Sci. USA 2020, 117, 17864–17875. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, G.; Haider, S.; Kunihs, V.; Saleh, L.; Pollheimer, J.; Fiala, C.; Hetey, S.; Feher, Z.; Szilagyi, A.; Than, N.G.; et al. Pivotal Role of the Transcriptional Co-Activator YAP in Trophoblast Stemness of the Developing Human Placenta. Proc. Natl. Acad. Sci. USA 2020, 117, 13562–13570. [Google Scholar] [CrossRef]
- Wang, H.; Xu, P.; Luo, X.; Hu, M.; Liu, Y.; Yang, Y.; Peng, W.; Bai, Y.; Chen, X.; Tan, B.; et al. Phosphorylation of Yes-Associated Protein Impairs Trophoblast Invasion and Migration: Implications for the Pathogenesis of Fetal Growth Restrictiondagger. Biol. Reprod. 2020, 103, 866–879. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wei, C.; Ma, Q.; Wang, W. Hippo-YAP1 Signaling Pathway and Severe Preeclampsia (SPE) in the Chinese Population. Pregnancy Hypertens. 2020, 19, 1–10. [Google Scholar] [CrossRef]
- Sun, M.; Na, Q.; Huang, L.; Song, G.; Jin, F.; Li, Y.; Hou, Y.; Kang, D.; Qiao, C. YAP Is Decreased in Preeclampsia and Regulates Invasion and Apoptosis of HTR-8/SVneo. Reprod. Sci. 2018, 25, 1382–1393. [Google Scholar] [CrossRef]
- Gill, M.K.; Christova, T.; Zhang, Y.Y.; Gregorieff, A.; Zhang, L.; Narimatsu, M.; Song, S.; Xiong, S.; Couzens, A.L.; Tong, J.; et al. A Feed Forward Loop Enforces YAP/TAZ Signaling during Tumorigenesis. Nat. Commun. 2018, 9, 3510. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Miyasaka, K.Y.; Kubo, A.; Kida, Y.S.; Nakagawa, O.; Hirate, Y.; Sasaki, H.; Ogura, T. Notch and Hippo Signaling Converge on Strawberry Notch 1 (Sbno1) to Synergistically Activate Cdx2 during Specification of the Trophectoderm. Sci. Rep. 2017, 7, 46135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soncin, F.; Parast, M.M. Role of Hippo Signaling Pathway in Early Placental Development. Proc. Natl. Acad. Sci. USA 2020, 117, 20354–20356. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fan, X.; Wang, R.; Lu, X.; Dang, Y.L.; Wang, H.; Lin, H.Y.; Zhu, C.; Ge, H.; Cross, J.C.; et al. Single-Cell RNA-Seq Reveals the Diversity of Trophoblast Subtypes and Patterns of Differentiation in the Human Placenta. Cell Res. 2018, 28, 819–832. [Google Scholar] [CrossRef]
- Ray, S.; Saha, A.; Ghosh, A.; Roy, N.; Kumar, R.P.; Meinhardt, G.; Mukerjee, A.; Gunewardena, S.; Kumar, R.; Knöfler, M.; et al. Hippo Signaling Cofactor, WWTR1, at the Crossroads of Human Trophoblast Progenitor Self-Renewal and Differentiation. Proc. Natl. Acad. Sci. USA 2022, 119, e2204069119. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.-C.; Fang, L.; Li, Y.; Thakur, A.; Hoodless, P.A.; Guo, Y.; Wang, Z.; Wu, Z.; Yan, Y.; Jia, Q.; et al. G Protein-Coupled Estrogen Receptor Stimulates Human Trophoblast Cell Invasion via YAP-Mediated ANGPTL4 Expression. Commun. Biol. 2021, 4, 1285. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Xu, T.; Tong, X.; Wang, Y.; Zhang, D.; Gao, D.; Zhang, L.; Ning, W.; Qi, X.; Ma, Y.; et al. Maternal Yes-Associated Protein Participates in Porcine Blastocyst Development via Modulation of Trophectoderm Epithelium Barrier Function. Cells 2019, 8, 1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totaro, A.; Panciera, T.; Piccolo, S. YAP/TAZ Upstream Signals and Downstream Responses. Nat. Cell Biol. 2018, 20, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Mo, H.-Q.; Tian, F.-J.; Zeng, W.-H.; Liu, X.-R.; Ma, X.-L.; Li, X.; Qin, S.; Fan, C.-F.; Lin, Y. EIF5A1 Promotes Trophoblast Migration and Invasion via ARAF-Mediated Activation of the Integrin/ERK Signaling Pathway. Cell Death Dis. 2018, 9, 926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccinni, M.P.; Raghupathy, R.; Saito, S.; Szekeres-Bartho, J. Cytokines, Hormones and Cellular Regulatory Mechanisms Favoring Successful Reproduction. Front. Immunol. 2021, 12, 717808. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.R.; Verma, P.; Singh, K. Immune-Endocrine Crosstalk during Pregnancy. Gen. Comp. Endocrinol. 2017, 242, 18–23. [Google Scholar] [CrossRef]
- Maliqueo, M.; Echiburu, B.; Crisosto, N. Sex Steroids Modulate Uterine-Placental Vasculature: Implications for Obstetrics and Neonatal Outcomes. Front. Physiol. 2016, 7, 152. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jin, S. Mitigating Placental Injuries through Up-Regulating DAF in Experimental APS Mice: New Mechanism of Progesterone. Clin. Exp. Immunol. 2019, 197, 376–386. [Google Scholar] [CrossRef]
- Lan, C.; Cao, N.; Chen, C.; Qu, S.; Fan, C.; Luo, H.; Zeng, A.; Yu, C.; Xue, Y.; Ren, H.; et al. Progesterone, via Yes-Associated Protein, Promotes Cardiomyocyte Proliferation and Cardiac Repair. Cell Prolif. 2020, 53, e12910. [Google Scholar] [CrossRef]
- Suzuki, D.; Okura, K.; Nagakura, S.; Ogawa, H. CDX2 Downregulation in Mouse Mural Trophectoderm during Peri-implantation Is Heteronomous, Dependent on the YAP-TEAD Pathway and Controlled by Estrogen-induced Factors. Reprod. Med. Biol. 2022, 21, e12446. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, K.; Breen, K.; Cohen, H.; Jacobsen, S.; Middeldorp, S.; Pavord, S.; Regan, L.; Roccatello, D.; Robinson, S.E.; Sciascia, S.; et al. HYdroxychloroquine to Improve Pregnancy Outcome in Women with AnTIphospholipid Antibodies (HYPATIA) Protocol: A Multinational Randomized Controlled Trial of Hydroxychloroquine versus Placebo in Addition to Standard Treatment in Pregnant Women with Antiphospholipid Syndrome or Antibodies. Semin. Thromb. Hemost. 2017, 43, 562–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, A.M.; Mulla, M.J.; Chamley, L.W.; Cadavid, A.P.; Abrahams, V.M. Aspirin-Triggered Lipoxin Prevents Antiphospholipid Antibody Effects on Human Trophoblast Migration and Endothelial Cell Interactions. Arthritis Rheumatol. 2015, 67, 488–497. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence5′-3′ | |
|---|---|
| YAP-F | ATGAACTCGGCTTCAGGTC |
| YAP-R | AGCCAAGAGGTGGTCTTGTT |
| MMP2-F | GATACCCCTTTGACGGTAAGGA |
| MMP2-R | CCTTCTCCCAAGGTCCATAGC |
| MMP9-F | AGACCTGGGCAGATTCCAAAC |
| MMP9-R | CGGCAAGTCTTCCGAGTAGT |
| GAPDH-F | GGAGCGAGATCCCTCCAAAAT |
| GAPDH-R | GGCTGTTGTCATACTTCTCATGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Z.; Tang, Z.; Guan, H.; Leung, W.; Wang, L.; Xia, H.; Zhang, W. Inactivation of Yes-Associated Protein Mediates Trophoblast Dysfunction: A New Mechanism of Pregnancy Loss Associated with Anti-Phospholipid Antibodies? Biomedicines 2022, 10, 3296. https://doi.org/10.3390/biomedicines10123296
Huang Z, Tang Z, Guan H, Leung W, Wang L, Xia H, Zhang W. Inactivation of Yes-Associated Protein Mediates Trophoblast Dysfunction: A New Mechanism of Pregnancy Loss Associated with Anti-Phospholipid Antibodies? Biomedicines. 2022; 10(12):3296. https://doi.org/10.3390/biomedicines10123296
Chicago/Turabian StyleHuang, Zengshu, Zhijing Tang, Haiyun Guan, Wingting Leung, Lu Wang, Hexia Xia, and Wei Zhang. 2022. "Inactivation of Yes-Associated Protein Mediates Trophoblast Dysfunction: A New Mechanism of Pregnancy Loss Associated with Anti-Phospholipid Antibodies?" Biomedicines 10, no. 12: 3296. https://doi.org/10.3390/biomedicines10123296
APA StyleHuang, Z., Tang, Z., Guan, H., Leung, W., Wang, L., Xia, H., & Zhang, W. (2022). Inactivation of Yes-Associated Protein Mediates Trophoblast Dysfunction: A New Mechanism of Pregnancy Loss Associated with Anti-Phospholipid Antibodies? Biomedicines, 10(12), 3296. https://doi.org/10.3390/biomedicines10123296