
Role of NLRP7 in Normal and Malignant Trophoblast Cells

 , , ,
, , ,  ,
,  ,
,  and
and 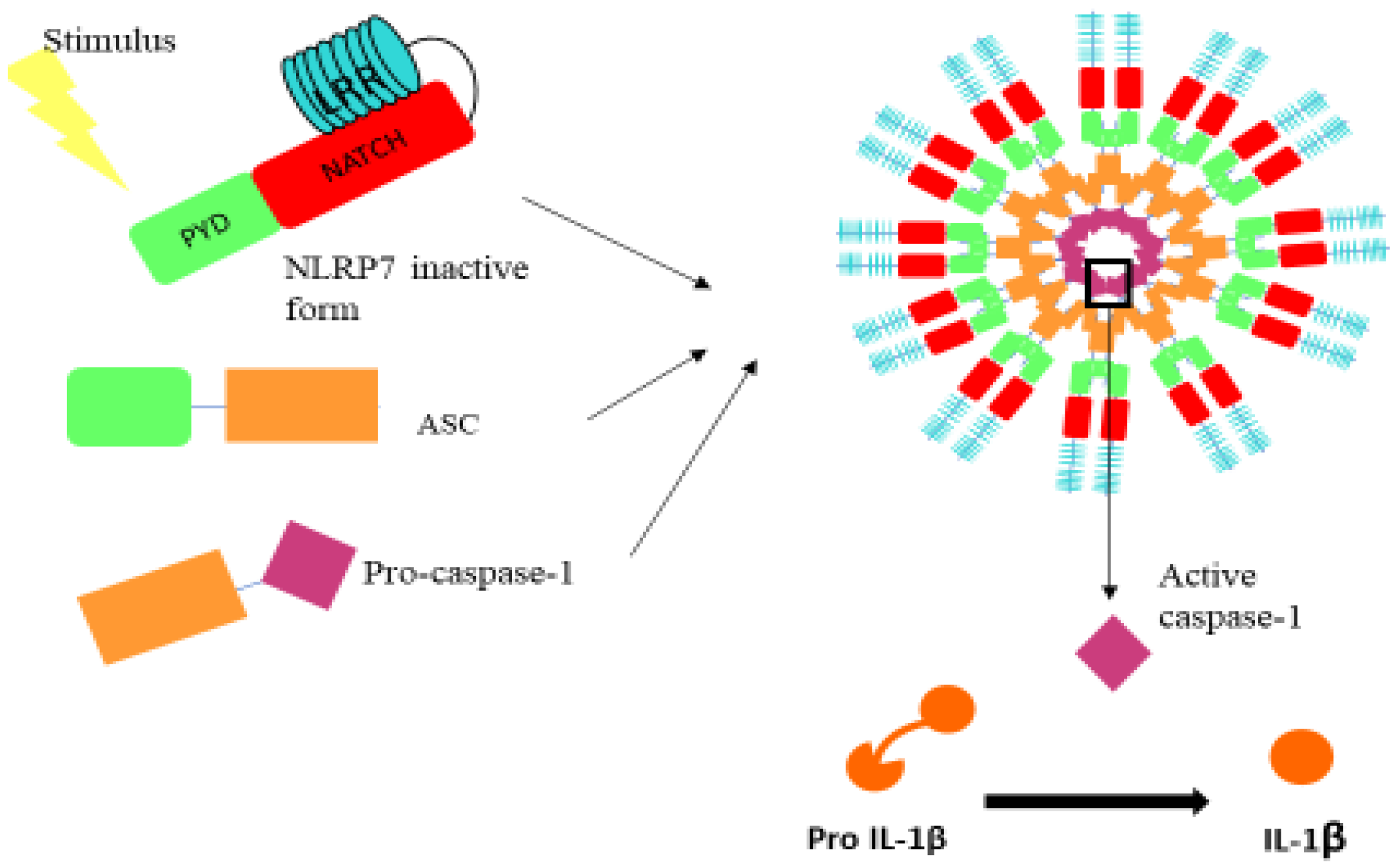{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. NLRP7 Inflammasome: Generalities
NLRP7 Expression Pattern and Functions
3. NLRP7 and Normal Pregnancy
4. NLRP7 in Pregnancy Pathologies
4.1. NLRP7 and Recurrent Hydatidiform Moles
4.2. NLRP7 and Fetal Growth Restriction
4.3. NLRP7 and Preeclampsia
5. NLRs and Cancer
5.1. NLRP7 and Gestational Trophoblastic Diseases
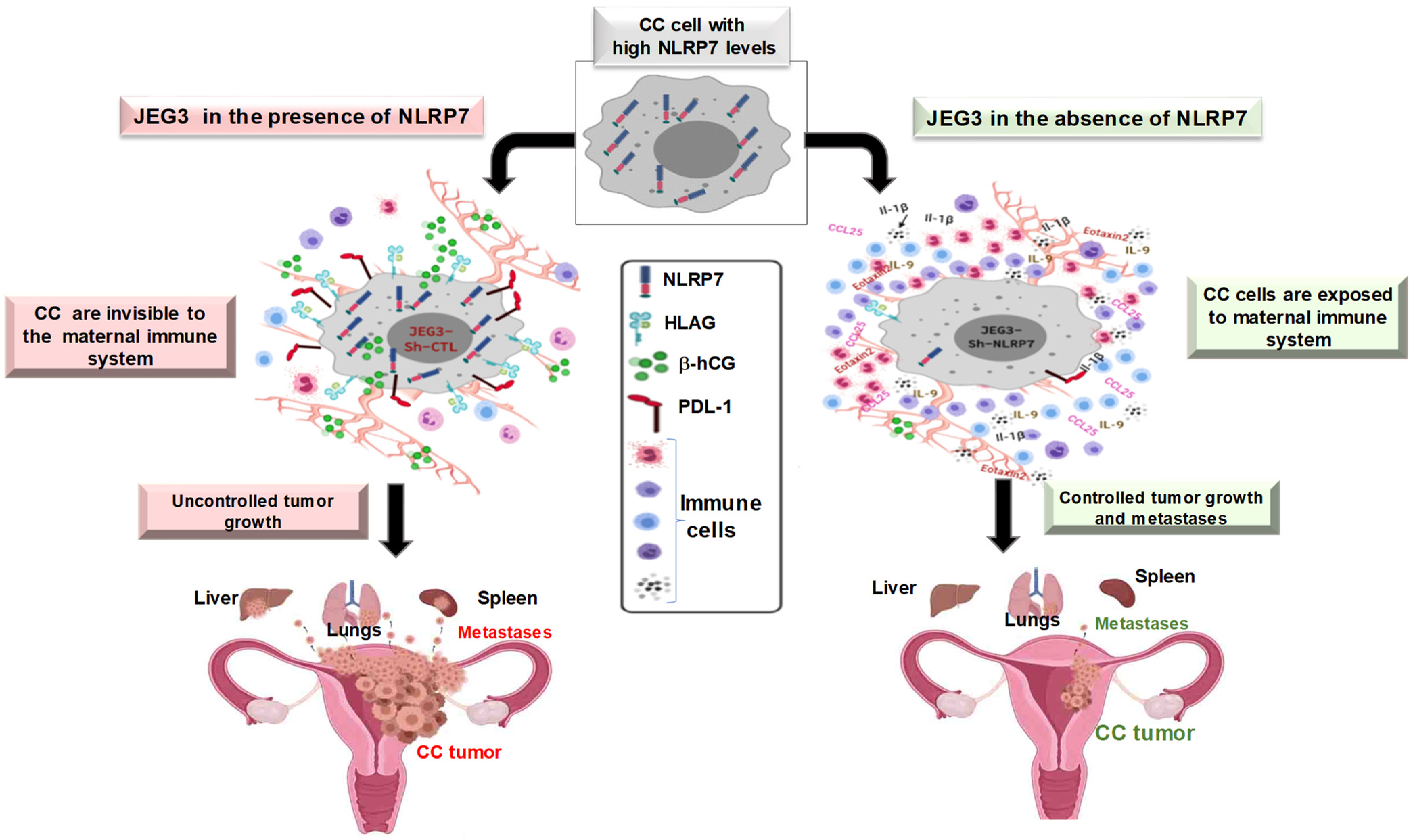
5.2. NLRP7 and Choriocarcinoma
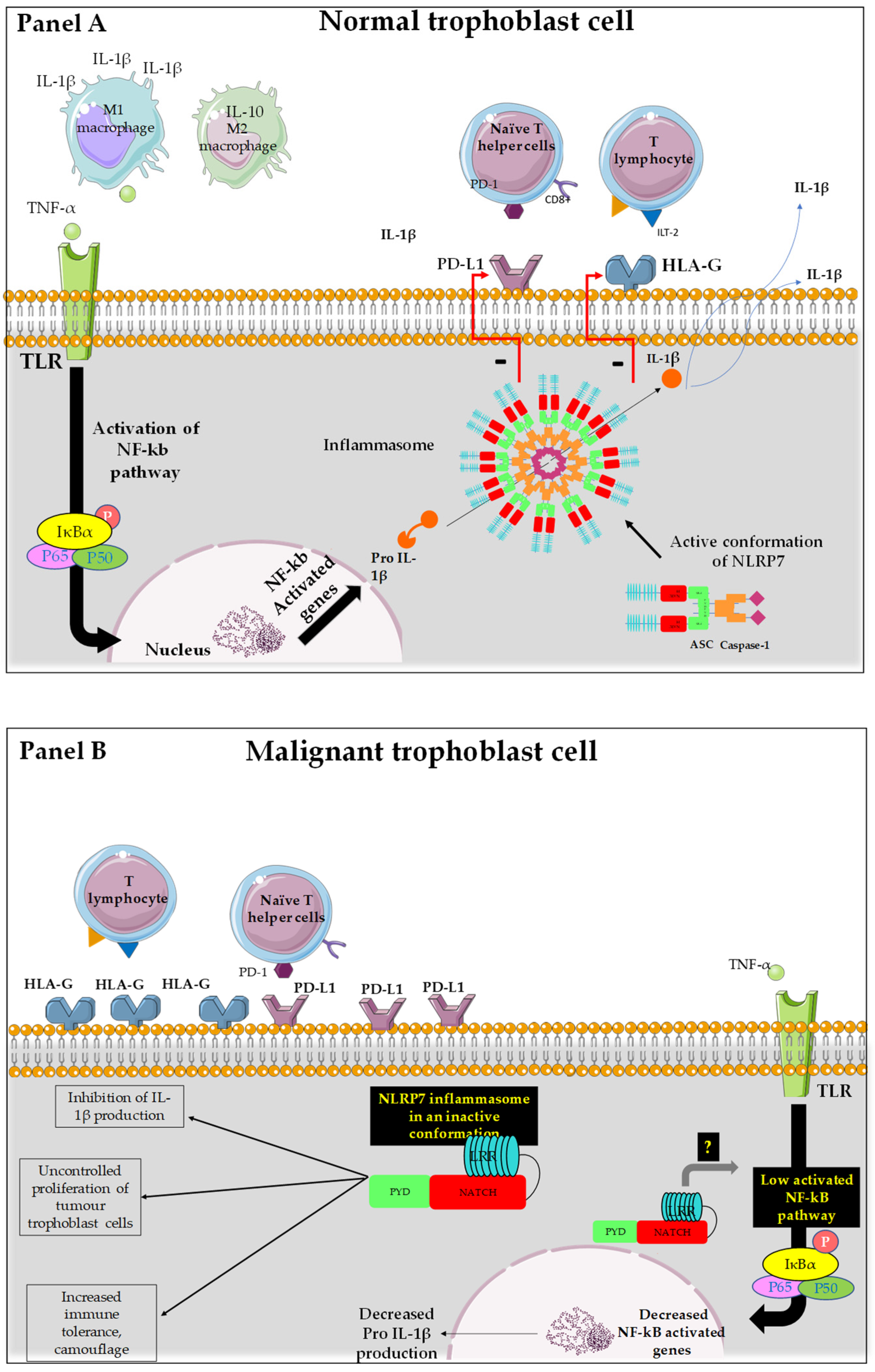
5.3. Proposed Mechanism of NLRP7 Control of Maternal Microenvironment
5.4. HLA Family in Normal and Tumor Placenta
5.5. PDL-1 in Normal and Tumor Trophoblast Cells
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Huang, X.; Wang, L.; Zhao, S.; Liu, H.; Chen, S.; Wu, L.; Liu, L.; Ding, J.; Yang, H.; Maxwell, A.; et al. Pregnancy Induces an Immunological Memory Characterized by Maternal Immune Alterations Through Specific Genes Methylation. Front. Immunol. 2021, 12, 686676. [Google Scholar] [CrossRef] [PubMed]
- Murthi, P.; Pinar, A.A.; Dimitriadis, E.; Samuel, C.S. Inflammasomes-A Molecular Link for Altered Immunoregulation and Inflammation Mediated Vascular Dysfunction in Preeclampsia. Int. J. Mol. Sci. 2020, 21, 1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnier, V.; Traboulsi, W.; Salomon, A.; Brouillet, S.; Fournier, T.; Winkler, C.; Desvergne, B.; Hoffmann, P.; Zhou, Q.Y.; Congiu, C.; et al. PPARgamma controls pregnancy outcome through activation of EG-VEGF: New insights into the mechanism of placental development. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E357–E369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Red-Horse, K.; Drake, P.M.; Fisher, S.J. Human pregnancy: The role of chemokine networks at the fetal-maternal interface. Expert Rev. Mol. Med. 2004, 6, 1–14. [Google Scholar] [CrossRef]
- Pijnenborg, R. Establishment of uteroplacental circulation. Reprod. Nutr. Dev. 1988, 28, 1581–1586. [Google Scholar] [CrossRef]
- Zhou, Y.; Fisher, S.J.; Janatpour, M.; Genbacev, O.; Dejana, E.; Wheelock, M.; Damsky, C.H. Human cytotrophoblasts adopt a vascular phenotype as they differentiate. A strategy for successful endovascular invasion? J. Clin. Investig. 1997, 99, 2139–2151. [Google Scholar] [CrossRef]
- Burton, G.J.; Jaunaiux, E. Maternal vascularisation of the human placenta: Does the embryo develop in a hypoxic environment? Gynecol. Obstet. Fertil. 2001, 29, 503–508. [Google Scholar] [CrossRef]
- Brien, M.E.; Duval, C.; Palacios, J.; Boufaied, I.; Hudon-Thibeault, A.A.; Nadeau-Vallee, M.; Vaillancourt, C.; Sibley, C.P.; Abrahams, V.M.; Jones, R.L.; et al. Uric Acid Crystals Induce Placental Inflammation and Alter Trophoblast Function via an IL-1-Dependent Pathway: Implications for Fetal Growth Restriction. J. Immunol. 2017, 198, 443–451. [Google Scholar] [CrossRef] [Green Version]
- Messaed, C.; Akoury, E.; Djuric, U.; Zeng, J.; Saleh, M.; Gilbert, L.; Seoud, M.; Qureshi, S.; Slim, R. NLRP7, a nucleotide oligomerization domain-like receptor protein, is required for normal cytokine secretion and co-localizes with Golgi and the microtubule-organizing center. J. Biol. Chem. 2011, 286, 43313–43323. [Google Scholar] [CrossRef] [Green Version]
- Alfaidy, N.; Gupta, S.; DeMarco, C.; Caniggia, I.; Challis, J.R. Oxygen regulation of placental 11 beta-hydroxysteroid dehydrogenase 2: Physiological and pathological implications. J. Clin. Endocrinol. Metab. 2002, 87, 4797–4805. [Google Scholar] [CrossRef] [Green Version]
- Bardin, N.; Murthi, P.; Alfaidy, N. Normal and pathological placental angiogenesis. BioMed Res. Int. 2015, 2015, 354359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouillet, S.; Hoffmann, P.; Chauvet, S.; Salomon, A.; Chamboredon, S.; Sergent, F.; Benharouga, M.; Feige, J.J.; Alfaidy, N. Revisiting the role of hCG: New regulation of the angiogenic factor EG-VEGF and its receptors. Cell Mol. Life Sci. 2012, 69, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Brouillet, S.; Murthi, P.; Hoffmann, P.; Salomon, A.; Sergent, F.; De Mazancourt, P.; Dakouane-Giudicelli, M.; Dieudonne, M.N.; Rozenberg, P.; Vaiman, D.; et al. EG-VEGF controls placental growth and survival in normal and pathological pregnancies: Case of fetal growth restriction (FGR). Cell Mol. Life Sci. 2013, 70, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Murthi, P.; Brouillet, S.; Pratt, A.; Borg, A.; Kalionis, B.; Goffin, F.; Tsatsaris, V.; Munaut, C.; Feige, J.; Benharouga, M.; et al. An EG-VEGF-dependent decrease in homeobox gene NKX3.1 contributes to cytotrophoblast dysfunction: A possible mechanism in human fetal growth restriction. Mol. Med. 2015, 21, 645–656. [Google Scholar] [CrossRef]
- Bolze, P.A.; Riedl, C.; Massardier, J.; Lotz, J.P.; You, B.; Schott, A.M.; Hajri, T.; Golfier, F. Mortality rate of gestational trophoblastic neoplasia with a FIGO score of ≥13. Am. J. Obstet. Gynecol. 2016, 214, 390.e1–390.e8. [Google Scholar] [CrossRef]
- Nguyen, N.M.; Slim, R. Genetics and Epigenetics of Recurrent Hydatidiform Moles: Basic Science and Genetic Counselling. Curr. Obstet. Gynecol. Rep. 2014, 3, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Seckl, M.J.; Fisher, R.A.; Salerno, G.; Rees, H.; Paradinas, F.J.; Foskett, M.; Newlands, E.S. Choriocarcinoma and partial hydatidiform moles. Lancet 2000, 356, 36–39. [Google Scholar] [CrossRef]
- Seckl, M.J.; Sebire, N.J.; Berkowitz, R.S. Gestational trophoblastic disease. Lancet 2010, 376, 717–729. [Google Scholar] [CrossRef]
- Collet, C.; Lopez, J.; Battail, C.; Allias, F.; Devouassoux-Shisheboran, M.; Patrier, S.; Lemaitre, N.; Hajri, T.; Massardier, J.; You, B.; et al. Transcriptomic Characterization of Postmolar Gestational Choriocarcinoma. Biomedicines 2021, 9, 1474. [Google Scholar] [CrossRef]
- Abi Nahed, R.; Reynaud, D.; Borg, A.J.; Traboulsi, W.; Wetzel, A.; Sapin, V.; Brouillet, S.; Dieudonne, M.N.; Dakouane-Giudicelli, M.; Benharouga, M.; et al. NLRP7 is increased in human idiopathic fetal growth restriction and plays a critical role in trophoblast differentiation. J. Mol. Med. 2019, 97, 355–367. [Google Scholar] [CrossRef]
- Slim, R.; Coullin, P.; Diatta, A.L.; Chebaro, W.; Courtin, D.; Abdelhak, S.; Garcia, A. NLRP7 and the genetics of post-molar choriocarcinomas in Senegal. Mol. Hum. Reprod. 2012, 18, 52–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khare, S.; Dorfleutner, A.; Bryan, N.B.; Yun, C.; Radian, A.D.; de Almeida, L.; Rojanasakul, Y.; Stehlik, C. An NLRP7-containing inflammasome mediates recognition of microbial lipopeptides in human macrophages. Immunity 2012, 36, 464–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radian, A.D.; de Almeida, L.; Dorfleutner, A.; Stehlik, C. NLRP7 and related inflammasome activating pattern recognition receptors and their function in host defense and disease. Microbes Infect. 2013, 15, 630–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesavardhana, S.; Malireddi, R.K.S.; Kanneganti, T.D. Caspases in Cell Death, Inflammation, and Pyroptosis. Annu. Rev. Immunol. 2020, 38, 567–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, M.T.; Sceneay, J.; Paget, C.; Wong, C.S.; Duret, H.; Tschopp, J.; Moller, A.; Smyth, M.J. NLRP3 suppresses NK cell-mediated responses to carcinogen-induced tumors and metastases. Cancer Res. 2012, 72, 5721–5732. [Google Scholar] [CrossRef] [Green Version]
- Terlizzi, M.; Casolaro, V.; Pinto, A.; Sorrentino, R. Inflammasome: Cancer’s friend or foe? Pharmacol. Ther. 2014, 143, 24–33. [Google Scholar] [CrossRef]
- Lupfer, C.; Kanneganti, T.D. Unsolved Mysteries in NLR Biology. Front. Immunol. 2013, 4, 285. [Google Scholar] [CrossRef] [Green Version]
- Carriere, J.; Dorfleutner, A.; Stehlik, C. NLRP7: From inflammasome regulation to human disease. Immunology 2021, 163, 363–376. [Google Scholar] [CrossRef]
- Tsai, P.Y.; Chen, K.R.; Li, Y.C.; Kuo, P.L. NLRP7 Is Involved in the Differentiation of the Decidual Macrophages. Int. J. Mol. Sci. 2019, 20, 5994. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Wang, Y.; Hasegawa, M.; Imamura, R.; Suda, T. PYPAF3, a PYRIN-containing APAF-1-like protein, is a feedback regulator of caspase-1-dependent interleukin-1beta secretion. J. Biol. Chem. 2005, 280, 21720–21725. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.; Mount, S.; Reddy, R.; Slim, R.; Wong, C.; Jobanputra, V.; Clifford, P.; Merrill, L.; Brown, S. Recurrent pregnancy loss in a woman with NLRP7 mutation: Not all molar pregnancies can be easily classified as either “partial” or “complete” hydatidiform moles. Int. J. Gynecol. Pathol. 2013, 32, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.M.; Zhang, L.; Reddy, R.; Dery, C.; Arseneau, J.; Cheung, A.; Surti, U.; Hoffner, L.; Seoud, M.; Zaatari, G.; et al. Comprehensive genotype-phenotype correlations between NLRP7 mutations and the balance between embryonic tissue differentiation and trophoblastic proliferation. J. Med. Genet. 2014, 51, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S.; Kinoshita, T.; Ohno, Y.; Minamoto, T.; Suzuki, N.; Inoue, M.; Suda, T. Expression of NLRP7 (PYPAF3, NALP7) protein in endometrial cancer tissues. Anticancer Res. 2008, 28, 2493–2497. [Google Scholar]
- Sills, E.S.; Obregon-Tito, A.J.; Gao, H.; McWilliams, T.K.; Gordon, A.T.; Adams, C.A.; Slim, R. Pathogenic variant in NLRP7 (19q13.42) associated with recurrent gestational trophoblastic disease: Data from early embryo development observed during in vitro fertilization. Clin. Exp. Reprod. Med. 2017, 44, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Dixon, M.; Zucchelli, M.; Hambiliki, F.; Levkov, L.; Hovatta, O.; Kere, J. Expression analysis of the NLRP gene family suggests a role in human preimplantation development. PLoS ONE 2008, 3, e2755. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Y.; Yu, P.H.; Li, Y.C.; Kuo, P.L. NLRP7 contributes to in vitro decidualization of endometrial stromal cells. Reprod. Biol. Endocrinol. 2017, 15, 66. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Tian, X.; Lv, D.; Zhang, L.; Zhang, Z.; Wang, J.; Yang, M.; Tao, J.; Ma, T.; Wu, H.; et al. NLRP7 is expressed in the ovine ovary and associated with in vitro pre-implantation embryo development. Reproduction 2019, 158, 415–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mor, G.; Cardenas, I. The immune system in pregnancy: A unique complexity. Am. J. Reprod. Immunol. 2010, 63, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Tong, M.; Abrahams, V.M. Immunology of the Placenta. Obstet. Gynecol. Clin. N. Am. 2020, 47, 49–63. [Google Scholar] [CrossRef]
- Stodle, G.S.; Silva, G.B.; Tangeras, L.H.; Gierman, L.M.; Nervik, I.; Dahlberg, U.E.; Sun, C.; Aune, M.H.; Thomsen, L.C.V.; Bjorge, L.; et al. Placental inflammation in pre-eclampsia by Nod-like receptor protein (NLRP)3 inflammasome activation in trophoblasts. Clin. Exp. Immunol. 2018, 193, 84–94. [Google Scholar] [CrossRef] [Green Version]
- I., C.W.; Romao-Veiga, M.; Matias, M.L.; Fioratti, E.G.; Peracoli, J.C.; Borges, V.T.; Araujo, J.P., Jr.; Peracoli, M.T. Increased expression of NLRP3 inflammasome in placentas from pregnant women with severe preeclampsia. J. Reprod. Immunol. 2017, 123, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.N.; Hay, D.P. A clear and present danger: Inflammasomes DAMPing down disorders of pregnancy. Hum. Reprod. Update 2015, 21, 388–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Lopez, N.; Romero, R.; Panaitescu, B.; Leng, Y.; Xu, Y.; Tarca, A.L.; Faro, J.; Pacora, P.; Hassan, S.S.; Hsu, C.D. Inflammasome activation during spontaneous preterm labor with intra-amniotic infection or sterile intra-amniotic inflammation. Am. J. Reprod. Immunol. 2018, 80, e13049. [Google Scholar] [CrossRef] [PubMed]
- Pontillo, A.; Girardelli, M.; Agostinis, C.; Masat, E.; Bulla, R.; Crovella, S. Bacterial LPS differently modulates inflammasome gene expression and IL-1beta secretion in trophoblast cells, decidual stromal cells, and decidual endothelial cells. Reprod. Sci. 2013, 20, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lopez, N.; Motomura, K.; Miller, D.; Garcia-Flores, V.; Galaz, J.; Romero, R. Inflammasomes: Their Role in Normal and Complicated Pregnancies. J. Immunol. 2019, 203, 2757–2769. [Google Scholar] [CrossRef]
- Romero, R.; Xu, Y.; Plazyo, O.; Chaemsaithong, P.; Chaiworapongsa, T.; Unkel, R.; Than, N.G.; Chiang, P.J.; Dong, Z.; Xu, Z.; et al. A Role for the Inflammasome in Spontaneous Labor at Term. Am. J. Reprod. Immunol. 2018, 79, e12440. [Google Scholar] [CrossRef]
- Co, E.C.; Gormley, M.; Kapidzic, M.; Rosen, D.B.; Scott, M.A.; Stolp, H.A.; McMaster, M.; Lanier, L.L.; Barcena, A.; Fisher, S.J. Maternal decidual macrophages inhibit NK cell killing of invasive cytotrophoblasts during human pregnancy. Biol. Reprod. 2013, 88, 155. [Google Scholar] [CrossRef]
- Alici-Garipcan, A.; Ozcimen, B.; Suder, I.; Ulker, V.; Onder, T.T.; Ozoren, N. NLRP7 plays a functional role in regulating BMP4 signaling during differentiation of patient-derived trophoblasts. Cell Death Dis. 2020, 11, 658. [Google Scholar] [CrossRef]
- Slim, R.; Wallace, E.P. NLRP7 and the Genetics of Hydatidiform Moles: Recent Advances and New Challenges. Front. Immunol. 2013, 4, 242. [Google Scholar] [CrossRef] [Green Version]
- Hayward, B.E.; De Vos, M.; Talati, N.; Abdollahi, M.R.; Taylor, G.R.; Meyer, E.; Williams, D.; Maher, E.R.; Setna, F.; Nazir, K.; et al. Genetic and epigenetic analysis of recurrent hydatidiform mole. Hum. Mutat. 2009, 30, E629–E639. [Google Scholar] [CrossRef]
- Estrada, H.; Buentello, B.; Zenteno, J.C.; Fiszman, R.; Aguinaga, M. The p.L750V mutation in the NLRP7 gene is frequent in Mexican patients with recurrent molar pregnancies and is not associated with recurrent pregnancy loss. Prenat. Diagn. 2013, 33, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Alfaidy, N.; Chauvet, S.; Donadio-Andrei, S.; Salomon, A.; Saoudi, Y.; Richaud, P.; Aude-Garcia, C.; Hoffmann, P.; Andrieux, A.; Moulis, J.M.; et al. Prion protein expression and functional importance in developmental angiogenesis: Role in oxidative stress and copper homeostasis. Antioxid. Redox Signal. 2013, 18, 400–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappas, M.; McCracken, S.; McKelvey, K.; Lim, R.; James, J.; Roberts, C.T.; Fournier, T.; Alfaidy, N.; Powell, K.L.; Borg, A.J.; et al. Formyl peptide receptor-2 is decreased in foetal growth restriction and contributes to placental dysfunction. Mol. Hum. Reprod. 2018, 24, 94–109. [Google Scholar] [CrossRef] [PubMed]
- Soellner, L.; Begemann, M.; Degenhardt, F.; Geipel, A.; Eggermann, T.; Mangold, E. Maternal heterozygous NLRP7 variant results in recurrent reproductive failure and imprinting disturbances in the offspring. Eur. J. Hum. Genet. 2017, 25, 924–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karan, D.; Tawfik, O.; Dubey, S. Expression analysis of inflammasome sensors and implication of NLRP12 inflammasome in prostate cancer. Sci. Rep. 2017, 7, 4378. [Google Scholar] [CrossRef]
- Allen, I.C.; TeKippe, E.M.; Woodford, R.M.; Uronis, J.M.; Holl, E.K.; Rogers, A.B.; Herfarth, H.H.; Jobin, C.; Ting, J.P. The NLRP3 inflammasome functions as a negative regulator of tumorigenesis during colitis-associated cancer. J. Exp. Med. 2010, 207, 1045–1056. [Google Scholar] [CrossRef]
- Ungerback, J.; Belenki, D.; Jawad ul-Hassan, A.; Fredrikson, M.; Fransen, K.; Elander, N.; Verma, D.; Soderkvist, P. Genetic variation and alterations of genes involved in NFkappaB/TNFAIP3- and NLRP3-inflammasome signaling affect susceptibility and outcome of colorectal cancer. Carcinogenesis 2012, 33, 2126–2134. [Google Scholar] [CrossRef] [Green Version]
- Verma, D.; Bivik, C.; Farahani, E.; Synnerstad, I.; Fredrikson, M.; Enerback, C.; Rosdahl, I.; Soderkvist, P. Inflammasome polymorphisms confer susceptibility to sporadic malignant melanoma. Pigment. Cell Melanoma Res. 2012, 25, 506–513. [Google Scholar] [CrossRef] [Green Version]
- Voronov, E.; Dotan, S.; Krelin, Y.; Song, X.; Elkabets, M.; Carmi, Y.; Rider, P.; Idan, C.; Romzova, M.; Kaplanov, I.; et al. Unique Versus Redundant Functions of IL-1alpha and IL-1beta in the Tumor Microenvironment. Front. Immunol. 2013, 4, 177. [Google Scholar] [CrossRef]
- Burgener, S.S.; Schroder, K. Placental inflammasome signaling: Protection for mother and baby. J. Exp. Med. 2021, 218, e20201738. [Google Scholar] [CrossRef]
- Okada, K.; Hirota, E.; Mizutani, Y.; Fujioka, T.; Shuin, T.; Miki, T.; Nakamura, Y.; Katagiri, T. Oncogenic role of NALP7 in testicular seminomas. Cancer Sci. 2004, 95, 949–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, F.T.; Katzorke, N.; Tempfer, C.; Kreimer, U.; Bizjak, G.I.; Fleisch, M.C.; Fehm, T.N. Gestational Trophoblastic Disorders: An Update in 2015. Geburtshilfe Frauenheilkd 2015, 75, 1043–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, P.; Winter, M.; Parker, V.; Harding, V.; Sita-Lumsden, A.; Fisher, R.A.; Harvey, R.; Unsworth, N.; Sarwar, N.; Short, D.; et al. Demographics, natural history and treatment outcomes of non-molar gestational choriocarcinoma: A UK population study. BJOG 2020, 127, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Reynaud, D.; Abi Nahed, R.; Lemaitre, N.; Bolze, P.A.; Traboulsi, W.; Sergent, F.; Battail, C.; Filhol, O.; Sapin, V.; Boufettal, H.; et al. NLRP7 Promotes Choriocarcinoma Growth and Progression through the Establishment of an Immunosuppressive Microenvironment. Cancers 2021, 13, 2999. [Google Scholar] [CrossRef]
- Traboulsi, W.; Sergent, F.; Boufettal, H.; Brouillet, S.; Slim, R.; Hoffmann, P.; Benlahfid, M.; Zhou, Q.Y.; Balboni, G.; Onnis, V.; et al. Antagonism of EG-VEGF Receptors as Targeted Therapy for Choriocarcinoma Progression In Vitro and In Vivo. Clin. Cancer Res. 2017, 23, 7130–7140. [Google Scholar] [CrossRef] [Green Version]
- Nilkaeo, A.; Bhuvanath, S. Interleukin-1 modulation of human placental trophoblast proliferation. Mediat. Inflamm. 2006, 2006, 79359. [Google Scholar] [CrossRef] [Green Version]
- Prutsch, N.; Fock, V.; Haslinger, P.; Haider, S.; Fiala, C.; Pollheimer, J.; Knofler, M. The role of interleukin-1beta in human trophoblast motility. Placenta 2012, 33, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Karki, R.; Kanneganti, T.D. Diverging inflammasome signals in tumorigenesis and potential targeting. Nat. Rev. Cancer 2019, 19, 197–214. [Google Scholar] [CrossRef]
- Schumacher, A.; Heinze, K.; Witte, J.; Poloski, E.; Linzke, N.; Woidacki, K.; Zenclussen, A.C. Human chorionic gonadotropin as a central regulator of pregnancy immune tolerance. J. Immunol. 2013, 190, 2650–2658. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, A.; Poloski, E.; Sporke, D.; Zenclussen, A.C. Luteinizing hormone contributes to fetal tolerance by regulating adaptive immune responses. Am. J. Reprod. Immunol. 2014, 71, 434–440. [Google Scholar] [CrossRef]
- Kallenbach, L.R.; Bianchi, D.W.; Peter, I.; Stroh, H.; Johnson, K.L. Maternal background strain influences fetal-maternal trafficking more than maternal immune competence in mice. J. Reprod. Immunol. 2011, 90, 188–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.; Schwartz, R.H. Molecular mechanisms for adaptive tolerance and other T cell anergy models. Semin. Immunol. 2007, 19, 140–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, J.F.; Fisher, S.J. The Placenta—Fast, Loose, and in Control. N. Engl. J. Med. 2021, 385, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Mosaferi, E.; Majidi, J.; Mohammadian, M.; Babaloo, Z.; Monfaredan, A.; Baradaran, B. HLA-G Expression Pattern: Reliable Assessment for Pregnancy Outcome Prediction. Adv. Pharm. Bull. 2013, 3, 443–446. [Google Scholar] [CrossRef] [Green Version]
- Tedesco, F.; Pausa, M.; Bulla, R.; Melazzini, S.; Guaschino, S. The maternal-fetal immune relationship in pregnancy. Recenti. Prog. Med. 1997, 88, 406–414. [Google Scholar]
- Wang, J.M.; Zhao, H.X.; Wang, L.; Gao, Z.Y.; Yao, Y.Q. The human leukocyte antigen G promotes trophoblast fusion and beta-hCG production through the Erk1/2 pathway in human choriocarcinoma cell lines. Biochem. Biophys. Res. Commun. 2013, 434, 460–465. [Google Scholar] [CrossRef]
- Rouas-Freiss, N.; Khalil-Daher, I.; Riteau, B.; Menier, C.; Paul, P.; Dausset, J.; Carosella, E.D. The immunotolerance role of HLA-G. Semin. Cancer Biol. 1999, 9, 3–12. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, W.; Zhang, D. Association of 14-bp insertion/deletion polymorphism of HLA-G gene with unexplained recurrent spontaneous abortion: A meta-analysis. Tissue Antigens 2013, 81, 108–115. [Google Scholar] [CrossRef]
- Xing, Y.F.; Pan, X.; Qian, B.; Shi, M.H. Expression of PD-1 and PD-L1 in the peripheral blood of advanced non-small-cell lung cancer patients and its implications. Zhonghua Yi Xue Za Zhi 2019, 99, 111–114. [Google Scholar] [CrossRef]
- Okuyama, M.; Mezawa, H.; Kawai, T.; Urashima, M. Elevated Soluble PD-L1 in Pregnant Women’s Serum Suppresses the Immune Reaction. Front. Immunol. 2019, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Bolze, P.A.; Patrier, S.; Massardier, J.; Hajri, T.; Abbas, F.; Schott, A.M.; Allias, F.; Devouassoux-Shisheboran, M.; Freyer, G.; Golfier, F.; et al. PD-L1 Expression in Premalignant and Malignant Trophoblasts From Gestational Trophoblastic Diseases Is Ubiquitous and Independent of Clinical Outcomes. Int. J. Gynecol. Cancer 2017, 27, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Sergent, F.; Hoffmann, P.; Brouillet, S.; Garnier, V.; Salomon, A.; Murthi, P.; Benharouga, M.; Feige, J.J.; Alfaidy, N. Sustained Endocrine Gland-Derived Vascular Endothelial Growth Factor Levels Beyond the First Trimester of Pregnancy Display Phenotypic and Functional Changes Associated With the Pathogenesis of Pregnancy-Induced Hypertension. Hypertension 2016, 68, 148–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abi Nahed, R.; Elkhoury Mikhael, M.; Reynaud, D.; Collet, C.; Lemaitre, N.; Michy, T.; Hoffmann, P.; Sergent, F.; Marquette, C.; Murthi, P.; et al. Role of NLRP7 in Normal and Malignant Trophoblast Cells. Biomedicines 2022, 10, 252. https://doi.org/10.3390/biomedicines10020252
Abi Nahed R, Elkhoury Mikhael M, Reynaud D, Collet C, Lemaitre N, Michy T, Hoffmann P, Sergent F, Marquette C, Murthi P, et al. Role of NLRP7 in Normal and Malignant Trophoblast Cells. Biomedicines. 2022; 10(2):252. https://doi.org/10.3390/biomedicines10020252
Chicago/Turabian StyleAbi Nahed, Roland, Maya Elkhoury Mikhael, Deborah Reynaud, Constance Collet, Nicolas Lemaitre, Thierry Michy, Pascale Hoffmann, Frederic Sergent, Christel Marquette, Padma Murthi, and et al. 2022. "Role of NLRP7 in Normal and Malignant Trophoblast Cells" Biomedicines 10, no. 2: 252. https://doi.org/10.3390/biomedicines10020252
APA StyleAbi Nahed, R., Elkhoury Mikhael, M., Reynaud, D., Collet, C., Lemaitre, N., Michy, T., Hoffmann, P., Sergent, F., Marquette, C., Murthi, P., Raia-Barjat, T., Alfaidy, N., & Benharouga, M. (2022). Role of NLRP7 in Normal and Malignant Trophoblast Cells. Biomedicines, 10(2), 252. https://doi.org/10.3390/biomedicines10020252