
Epigenetic Effects of Gender-Affirming Hormone Treatment: A Pilot Study of the ESR2 Promoter’s Methylation in AFAB People

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hormone Profile
2.2. DNA Extraction and AR and ER Polymorphisms
2.3. Epigenetic Analysis
2.4. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fisher, A.D.; Senofonte, G.; Cocchetti, C.; Guercio, G.; Lingiardi, V.; Meriggiola, M.C.; Mosconi, M.; Motta, G.; Ristori, J.; Speranza, A.M.; et al. SIGIS–SIAMS–SIE position statement of gender affirming hormonal treatment in transgender and non-binary people. J. Endocrinol. Investig. 2021, 1–17. [Google Scholar] [CrossRef]
- Mirabella, M.; Giovanardi, G.; Fortunato, A.; Senofonte, G.; Lombardo, F.; Lingiardi, V.; Speranza, A.M. The Body I Live in. Perceptions and Meanings of Body Dissatisfaction in Young Transgender Adults: A Qualitative Study. J. Clin. Med. 2020, 9, 3733. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.B.; Chavez, A.M.; Lipner, E.; Hantsoo, L.; Kornfield, S.L.; Davies, R.D.; Epperson, C.N. Gender-Affirming Hormone Use in Transgender Individuals: Impact on Behavioral Health and Cognition. Curr. Psychiatry Rep. 2018, 20, 110. [Google Scholar] [CrossRef] [PubMed]
- Hembree, W.C.; Cohen-Kettenis, P.T.; Gooren, L.; Hannema, S.E.; Meyer, W.J.; Murad, M.H.; Rosenthal, S.M.; Safer, J.D.; Tangpricha, V.; T’Sjoen, G.G. Endocrine Treatment of Gender-Dysphoric/Gender-Incongruent Persons: An Endocrine Society* Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2017, 102, 3869–3903. [Google Scholar] [CrossRef]
- D’Andrea, S.; Pallotti, F.; Senofonte, G.; Castellini, C.; Paoli, D.; Lombardo, F.; Lenzi, A.; Francavilla, S.; Francavilla, F.; Barbonetti, A. Polymorphic Cytosine-Adenine-Guanine Repeat Length of Androgen Receptor Gene and Gender Incongruence in Trans Women: A Systematic Review and Meta-Analysis of Case-Control Studies. J. Sex. Med. 2020, 17, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Almstrup, K.; Frederiksen, H.; Andersson, A.-M.; Juul, A. Levels of endocrine-disrupting chemicals are associated with changes in the peri-pubertal epigenome. Endocr. Connect. 2020, 9, 845–857. [Google Scholar] [CrossRef]
- Fernández, R.; Ramírez, K.; Gómez-Gil, E.; Cortés-Cortés, J.; Mora, M.; Aranda, G.; Zayas, E.D.; Esteva, I.; Almaraz, M.C.; Guillamon, A.; et al. Gender-Affirming Hormone Therapy Modifies the CpG Methylation Pattern of the ESR1 Gene Promoter After Six Months of Treatment in Transmen. J. Sex. Med. 2020, 17, 1795–1806. [Google Scholar] [CrossRef]
- Aranda, G.; Fernandez-Rebollo, E.; Pradas-Juni, M.; Hanzu, F.A.; Kalko, S.; Halperin, I.; Mora, M. Effects of sex steroids on the pattern of methylation and expression of the promoter region of estrogen and androgen receptors in people with gender dysphoria under cross-sex hormone treatment. J. Steroid Biochem. Mol. Biol. 2017, 172, 20–28. [Google Scholar] [CrossRef]
- Murphy, R.; Ibáñez, L.; Hattersley, A.; Tost, J. IGF2/H19 hypomethylation in a patient with very low birthweight, preocious pubarche and insulin resistance. BMC Med Genet. 2012, 13, 42. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Peterson, K.E.; Sánchez, B.N.; Dolinoy, D.C.; Mercado-Garcia, A.; Rojo, M.T.; Goodrich, J.M. Association of blood leukocyte DNA methylation at LINE-1 and growth-related candidate genes with pubertal onset and progression. Epigenetics 2018, 13, 1222–1233. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, B.O.; Bolour, S.; Woods, K.; Moore, A.; Azziz, R. Visually scoring hirsutism. Hum. Reprod. Updat. 2009, 16, 51–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallotti, F.; Senofonte, G.; Pelloni, M.; Cargnelutti, F.; Carlini, T.; Radicioni, A.F.; Rossi, A.; Lenzi, A.; Paoli, D.; Lombardo, F. Androgenetic alopecia: Effects of oral finasteride on hormone profile, reproduction and sexual function. Endocrinology 2020, 68, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Zitzmann, M.; Depenbusch, M.; Gromoll, J.; Nieschlag, E. X-Chromosome Inactivation Patterns and Androgen Receptor Functionality Influence Phenotype and Social Characteristics as Well as Pharmacogenetics of Testosterone Therapy in Klinefelter Patients. J. Clin. Endocrinol. Metab. 2004, 89, 6208–6217. [Google Scholar] [CrossRef]
- Grassetti, D.; Giannandrea, F.; Paoli, D.; Masciandaro, P.; Figura, V.; Carlini, T.; Rizzo, F.; Lombardo, F.; Lenzi, A.; Gandini, L. Androgen receptor polymorphisms and testicular cancer risk. Andrology 2015, 3, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitzwieser, M.; Entfellner, E.; Werner, B.; Pulverer, W.; Pfeiler, G.; Hacker, S.; Cichna-Markl, M. Hypermethylation of CDKN2A exon 2 in tumor, tumor-adjacent and tumor-distant tissues from breast cancer patients. BMC Cancer 2017, 17, 260. [Google Scholar] [CrossRef] [Green Version]
- Montrose, L.; Padmanabhan, V.; Goodrich, J.M.; Domino, S.E.; Treadwell, M.C.; Meeker, J.D.; Watkins, D.J.; Dolinoy, D.C. Maternal levels of endocrine disrupting chemicals in the first trimester of pregnancy are associated with infant cord blood DNA methylation. Epigenetics 2018, 13, 301–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.-C.; Galati, J.C.; Burrows, S.; Beilin, L.J.; Li, X.; Pennell, C.E.; Van Eekelen, J.; Mori, T.A.; Adams, L.A.; Craig, J.M. DNA methylation of the IGF2/H19 imprinting control region and adiposity distribution in young adults. Clin. Epigenetics 2012, 4, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deodati, A.; Inzaghi, E.; Liguori, A.; Puglianiello, A.; Germani, D.; Brufani, C.; Fintini, D.; Cappa, M.; Barbetti, F.; Cianfarani, S. IGF2Methylation Is Associated with Lipid Profile in Obese Children. Horm. Res. Paediatr. 2013, 79, 361–367. [Google Scholar] [CrossRef]
- Holtzman, L.; Gersbach, C.A. Editing the Epigenome: Reshaping the Genomic Landscape. Annu. Rev. Genom. Hum. Genet. 2018, 19, 43–71. [Google Scholar] [CrossRef]
- Chen, H.; Malentacchi, F.; Fambrini, M.; Harrath, A.H.; Huang, H.; Petraglia, F. Epigenetics of Estrogen and Progesterone Receptors in Endometriosis. Reprod. Sci. 2020, 27, 1967–1974. [Google Scholar] [CrossRef]
- Lomniczi, A.; Wright, H.; Ojeda, S.R. Epigenetic regulation of female puberty. Front. Neuroendocr. 2015, 36, 90–107. [Google Scholar] [CrossRef] [PubMed]
- Shalev, D.; Melamed, P. The role of the hypothalamus and pituitary epigenomes in central activation of the reproductive axis at puberty. Mol. Cell. Endocrinol. 2020, 518, 111031. [Google Scholar] [CrossRef] [PubMed]
- Cargnelutti, F.; Di Nisio, A.; Pallotti, F.; Sabovic, I.; Spaziani, M.; Tarsitano, M.G.; Paoli, D.; Foresta, C. Effects of endocrine disruptors on fetal testis development, male puberty, and transition age. Endocrinology 2021, 72, 358–374. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rodriguez, D.; Franssen, D.; Bakker, J.; Lomniczi, A.; Parent, A.-S. Cellular and molecular features of EDC exposure: Consequences for the GnRH network. Nat. Rev. Endocrinol. 2021, 17, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Sader, M.A.; McGrath, K.; Hill, M.D.; Bradstock, K.F.; Jimenez, M.; Handelsman, D.J.; Celermajer, D.S.; Death, A.K. Androgen receptor gene expression in leucocytes is hormonally regulated: Implications for gender differences in disease pathogenesis. Clin. Endocrinol. 2005, 62, 56–63. [Google Scholar] [CrossRef]
- Ben-Avraham, D.; Muzumdar, R.H.; Atzmon, G. Epigenetic genome-wide association methylation in aging and longevity. Epigenomics 2012, 4, 503–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocklandt, S.; Lin, W.; Sehl, M.E.; Sánchez, F.J.; Sinsheimer, J.S.; Horvath, S.; Vilain, E. Epigenetic Predictor of Age. PLoS ONE 2011, 6, e14821. [Google Scholar] [CrossRef] [Green Version]
- Florath, I.; Butterbach, K.; Müller, H.; Bewerunge-Hudler, M.; Brenner, H. Cross-sectional and longitudinal changes in DNA methylation with age: An epigenome-wide analysis revealing over 60 novel age-associated CpG sites. Hum. Mol. Genet. 2014, 23, 1186–1201. [Google Scholar] [CrossRef]
- Tan, Q.; Heijmans, B.T.; Hjelmborg, J.V.B.; Soerensen, M.; Christensen, K.; Christiansen, L. Epigenetic drift in the aging genome: A ten-year follow-up in an elderly twin cohort. Int. J. Epidemiol. 2016, 45, 1146–1158. [Google Scholar] [CrossRef] [Green Version]
- Ehlert, T.; Simon, P.; Moser, D.A. Epigenetics in Sports. Sports Med. 2013, 43, 93–110. [Google Scholar] [CrossRef]
- McGee, S.L.; Fairlie, E.; Garnham, A.P.; Hargreaves, M. Exercise-induced histone modifications in human skeletal muscle. J. Physiol. 2009, 587, 5951–5958. [Google Scholar] [CrossRef]
- Schwarzenbach, H. Impact of Physical Activity and Doping on Epigenetic Gene Regulation. Drug Test. Anal. 2011, 3, 682–687. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer/Sequence to Analyze | ESR2 | H19 |
|---|---|---|
| Forward PCR Primer | 5′-GGAGGTTGAGAGAAATAATTGTTTTTTGA- 3′ | 5′-TTTGTTGATTTTATTAAGGGAG-3′ |
| Reverse PCR Primer | 5′-[Biotin]-ATAAACACACCCACCTTACCTTCTCTA-3′ | 5′-[Biotin]-CTATAAATAAACCCCAACCAAAC-3′ |
| Sequencing Primer | 5′-GAAATAATTGTTTTTTGAAATTTG-3′ | 5′-GTGTGGAATTAGAAGT-3′ |
| Sequence to Analyze | TAGGGYGAAGAGTAGGYGGYGAGYGTTGGGTYGGGGAGGGATTATTYGAGTTGYGAYGGGTTTTGGGGTTGYGGGGTA | GGTYGYGYGGYGGTAGTGTAGGTTTATATATTATAGTT |
| Annealing Temperature (°C) | 61 °C for 30 s | 51 °C for 30 s |
| Age (Years) | 29.3 ± 12.6 (21) |
| BMI | 26.6 ± 1.4 (27) |
| CAG repeats expression (AR) | 23.1 ± 1.9 (23.5) |
| CA repeats expression (ERβ) | 22.1 ± 1.4 (21.5) |
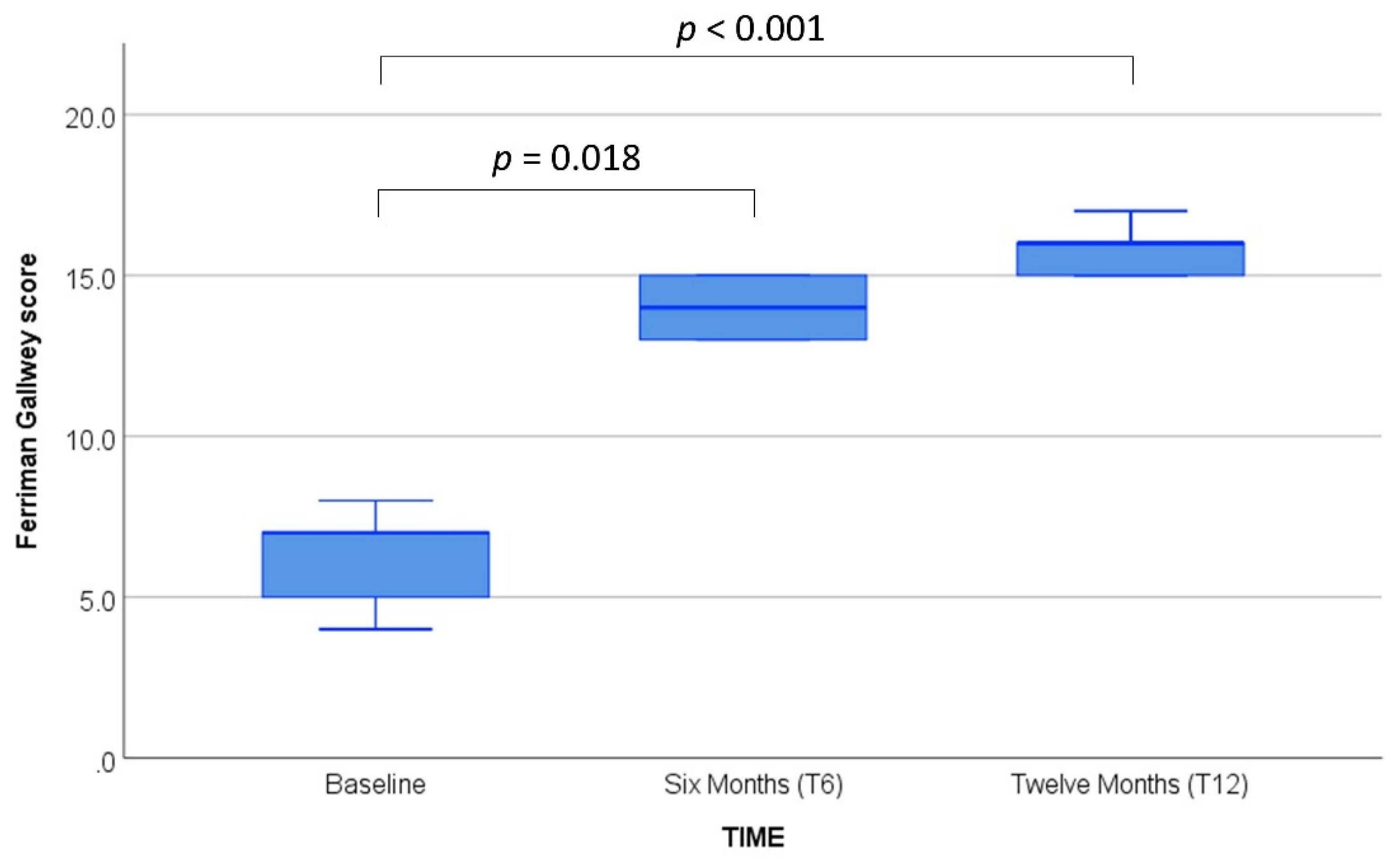
| Ferriman–Gallwey score | 6.4 ± 1.3 (7.0) |
| Creatinine (mg/dL) | 0.6 ± 0.1 (0.6) |
| Red blood cells (×106/mL) | 4.6 ± 0.5 (4.6) |
| Hemoglobin (g/dL) | 12.7 ± 1.2 (12.6) |
| Hematocrit (%) | 39.0 ± 3.1 (39.9) |
| White blood cells (×103/mL) | 7.1 ± 1.7 (7.1) |
| Platelets (×103/mL) | 243.8 ± 80.3 (252.0) |
| Glycemia (mg/dL) | 85.8 ± 11.8 (85.0) |
| HbA1c (%) | 5.1 ± 0.4 (5.1) |
| AST (UI/L) | 23.5 ± 19.8 (19) |
| ALT (UI/L) | 22.1 ± 10.8 (18.0) |
| γGT (UI/L) | 19.5 ± 12.9 (15.0) |
| Total cholesterol (mg/dL) | 153.8 ± 27.2 (150.0) |
| HDL (mg/dL) | 56.3 ± 11.6 (58.0) |
| LDL (mg/dL) | 80.5 ± 23.1 (85.0) |
| Triglycerides (mg/dL) | 83.8 ± 38.3 (71.0) |
| FSH (mUI/mL) | 10.5 ± 13.5 (5.4) |
| LH (mUI/mL) | 7.8 ± 4.6 (6.1) |
| Prolactin (ng/dL) | 13.2 ± 4.7 (12.2) |
| 17β estradiol (pg/mL) | 84.0 ± 86.3 (56.1) |
| Total testosterone (nmol/L) | 1.2 ± 0.5 (1.3) |
| Patient | CAG Repeats (Allele 1) | Percentage of Inactivation (Allele 1) | CAG Repeats (Allele 2) | Percentage of Inactivation (Allele 2) | CA Repeats (Allele 1) | CA Repeats (Allele 2) |
|---|---|---|---|---|---|---|
| #1 | 24 | 41.0 | 25 | 59.0 | 24 | 24 |
| #2 | 23 | 62.1 | 25 | 37.9 | 20 | 24 |
| #3 | 19 | 29.6 | 26 | 70.4 | 18 | 23 |
| #4 | 17 | 50.8 | 24 | 49.3 | 20 | 20 |
| #5 | 20 | 35.8 | 24 | 64.2 | 20 | 23 |
| #6 | 20 | 51.3 | 21 | 48.7 | 23 | 23 |
| #7 | 24 | 56.7 | 26 | 43.3 | 18 | 25 |
| #8 | 21 | 56.6 | 24 | 43.4 | 21 | 24 |
| #9 | 25 | 51.4 | 26 | 48.6 | 21 | 21 |
| #10 | 22 | 58.3 | 25 | 41.7 | 17 | 26 |
| #11 | 20 | / | 20 | / | 24 | 24 |
| #12 | 24 | 45.4 | 26 | 54.6 | 24 | 24 |
| #13 | 24 | 35.8 | 26 | 64.2 | 21 | 21 |
| B | CI 95% | Beta | p | |
|---|---|---|---|---|
| Coefficients from regression models predicting baseline ESR2 methylation. | ||||
| CA repeats | −0.529 | −1.302–0.243 | −0.414 | 0.533 |
| CAG repeats | −0.175 | −0.762–0.413 | −0.193 | 0.527 |
| Total testosterone | 1.143 | −0.852–3.139 | 0.355 | 0.233 |
| Estradiol | −0.007 | −0.019–0.006 | −0.334 | 0.164 |
| Age | 0.026 | −0.103–0.155 | 0.185 | 0.653 |
| Coefficients from regression models predicting T6 ESR2 methylation. | ||||
| CA repeats | 0.437 | −0.428–1.302 | 0.318 | 0.290 |
| CAG repeats | −0.111 | −0.751–0.529 | −0.114 | 0.711 |
| Total testosterone | 0.030 | −0.296–0.357 | 0.062 | 0.841 |
| Estradiol | 0.009 | −0.060–0.077 | 0.083 | 0.788 |
| Age | −0.034 | −0.130–0.062 | −0.230 | 0.450 |
| Coefficients from regression models predicting T12 ESR2 methylation. | ||||
| CA repeats | −0.110 | −1.062–0.841 | −0.077 | 0.803 |
| CAG repeats | 0.110 | −0.560–0.780 | 0.108 | 0.725 |
| Total testosterone | 0.280 | 0.086–0.474 | 0.691 | 0.009 |
| Estradiol | −0.009 | −0.028–0.011 | −0.276 | 0.361 |
| Age | 0.106 | 0.031–0.182 | 0.682 | 0.010 |
| B | CI 95% | Beta | p | Partial Eta Squared | |
|---|---|---|---|---|---|
| Linear regression model coefficients predicting the increase in ESR2 promoter methylation between baseline and T6. | |||||
| CA repeats | 1.400 | 0.357–2.444 | 0.822 | 0.016 | 0.590 |
| CAG repeats | −0.481 | −1.385–0.423 | −0.400 | 0.249 | 0.185 |
| Total testosterone | 0.445 | −0.061–0.850 | 0.728 | 0.076 | 0.382 |
| Estradiol | 0.079 | −0.016–0.073 | 0.609 | 0.090 | 0.356 |
| Age | −0.067 | −0.183–0.050 | −0.361 | 0.220 | 0.206 |
| Linear regression model coefficients predicting the increase in ESR2 promoter methylation between baseline and T12. | |||||
| B | CI 95% | Beta | p | Partial Eta Squared | |
| CA repeats | 0.498 | −0.270–1.266 | 0.304 | 0.169 | 0.251 |
| CAG repeats | 0.285 | −0.421–0.991 | 0.246 | 0.371 | 0.115 |
| Total testosterone | 0.169 | −0.179–0.518 | 0.367 | 0.289 | 0.159 |
| Estradiol | 0.014 | −0.007–0.036 | 0.407 | 0.152 | 0.270 |
| Age | 0.109 | 0.010–0.218 | 0.615 | 0.048 | 0.449 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pallotti, F.; Senofonte, G.; Konstantinidou, F.; Di Chiano, S.; Faja, F.; Rizzo, F.; Cargnelutti, F.; Krausz, C.; Paoli, D.; Lenzi, A.; et al. Epigenetic Effects of Gender-Affirming Hormone Treatment: A Pilot Study of the ESR2 Promoter’s Methylation in AFAB People. Biomedicines 2022, 10, 459. https://doi.org/10.3390/biomedicines10020459
Pallotti F, Senofonte G, Konstantinidou F, Di Chiano S, Faja F, Rizzo F, Cargnelutti F, Krausz C, Paoli D, Lenzi A, et al. Epigenetic Effects of Gender-Affirming Hormone Treatment: A Pilot Study of the ESR2 Promoter’s Methylation in AFAB People. Biomedicines. 2022; 10(2):459. https://doi.org/10.3390/biomedicines10020459
Chicago/Turabian StylePallotti, Francesco, Giulia Senofonte, Fani Konstantinidou, Silvia Di Chiano, Fabiana Faja, Flavio Rizzo, Francesco Cargnelutti, Csilla Krausz, Donatella Paoli, Andrea Lenzi, and et al. 2022. "Epigenetic Effects of Gender-Affirming Hormone Treatment: A Pilot Study of the ESR2 Promoter’s Methylation in AFAB People" Biomedicines 10, no. 2: 459. https://doi.org/10.3390/biomedicines10020459
APA StylePallotti, F., Senofonte, G., Konstantinidou, F., Di Chiano, S., Faja, F., Rizzo, F., Cargnelutti, F., Krausz, C., Paoli, D., Lenzi, A., Stuppia, L., Gatta, V., & Lombardo, F. (2022). Epigenetic Effects of Gender-Affirming Hormone Treatment: A Pilot Study of the ESR2 Promoter’s Methylation in AFAB People. Biomedicines, 10(2), 459. https://doi.org/10.3390/biomedicines10020459