
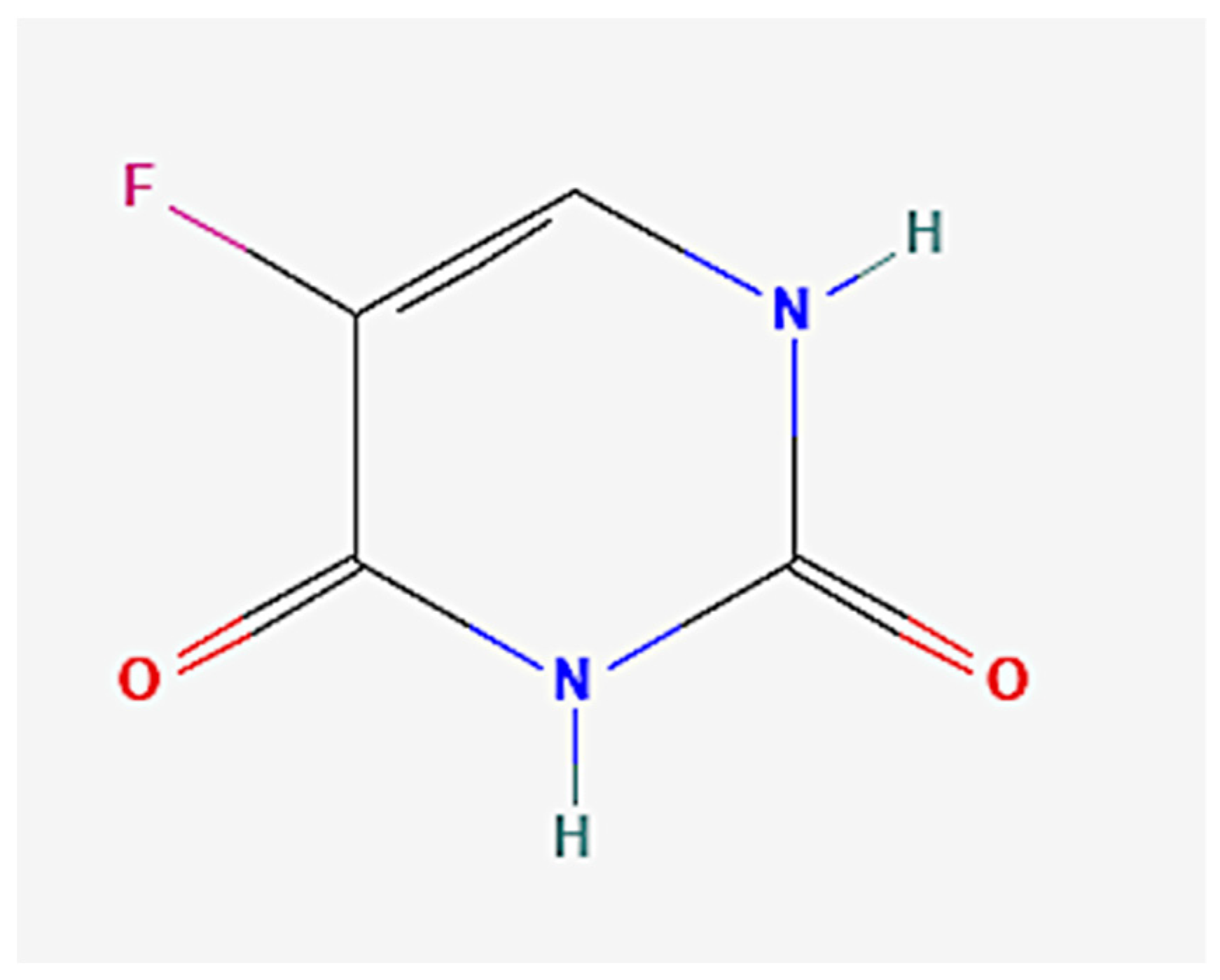
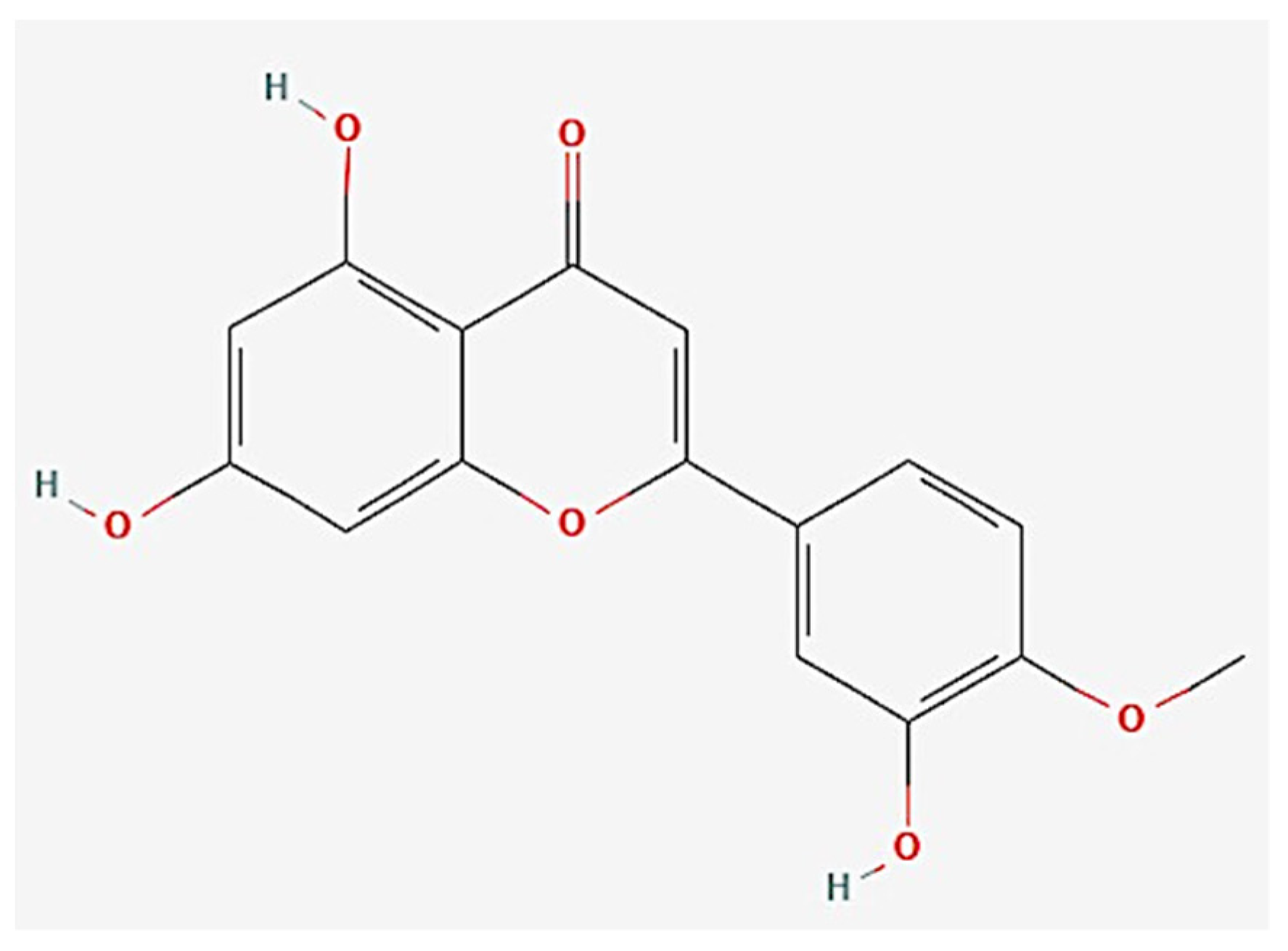
Diosmetin Exerts Synergistic Effects in Combination with 5-Fluorouracil in Colorectal Cancer Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Condition
2.2. MTT Assay to Determine Cell Viability
2.3. Synergistic Effect Analysis
2.4. AO/PI Double Staining Assay
2.5. Annexin V/PI Assay for the Detection of Cell Apoptosis
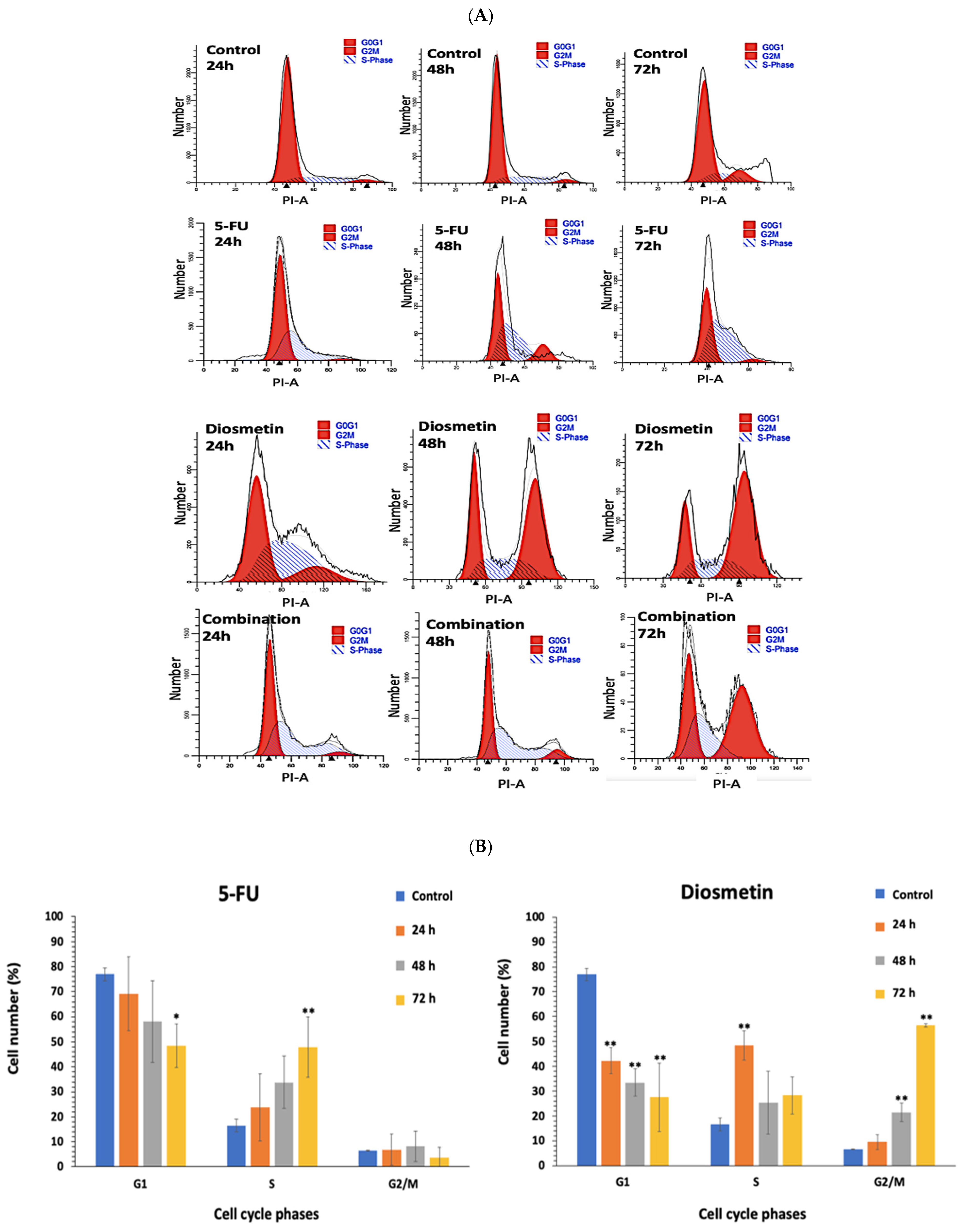
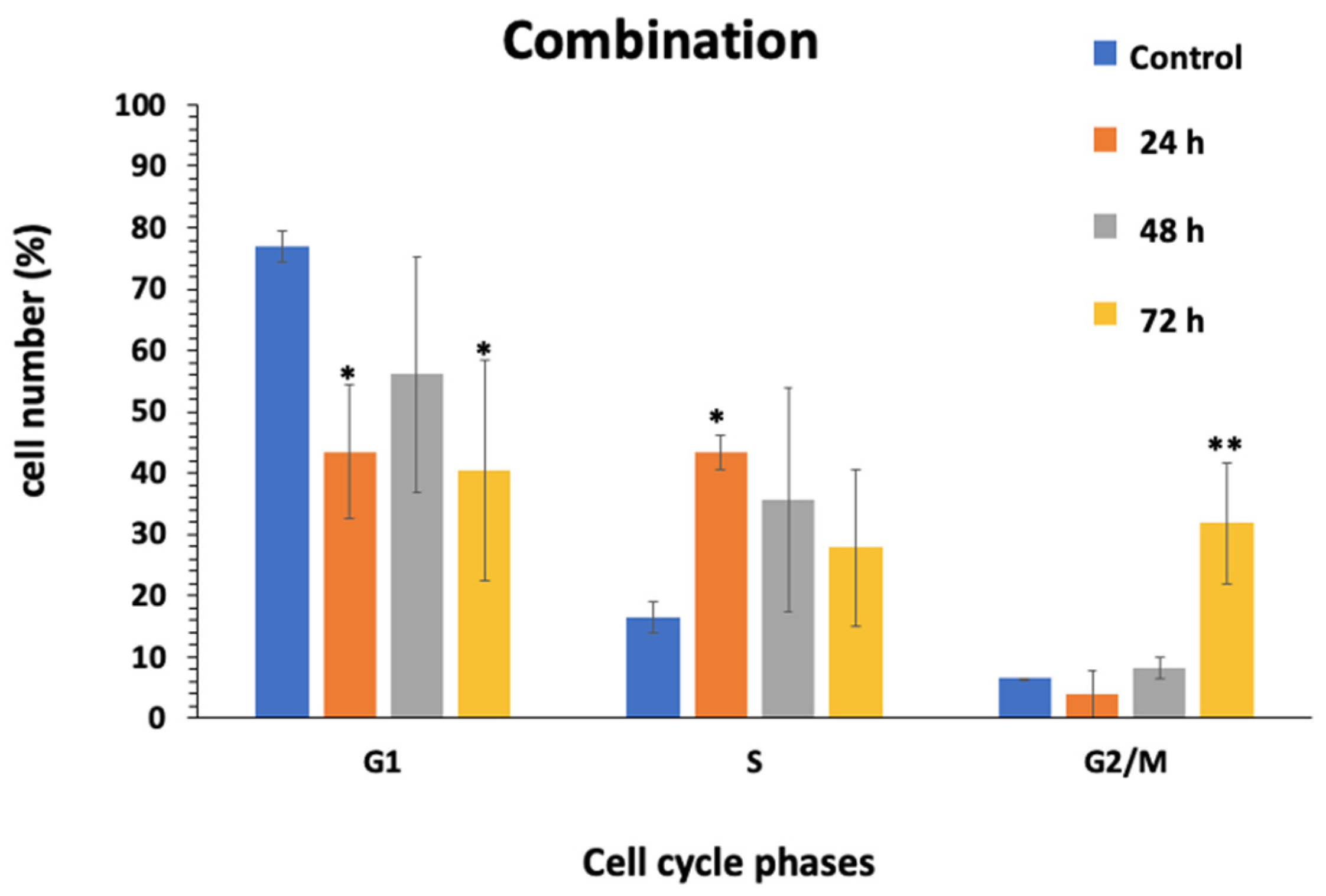
2.6. Cell Cycle Assessment
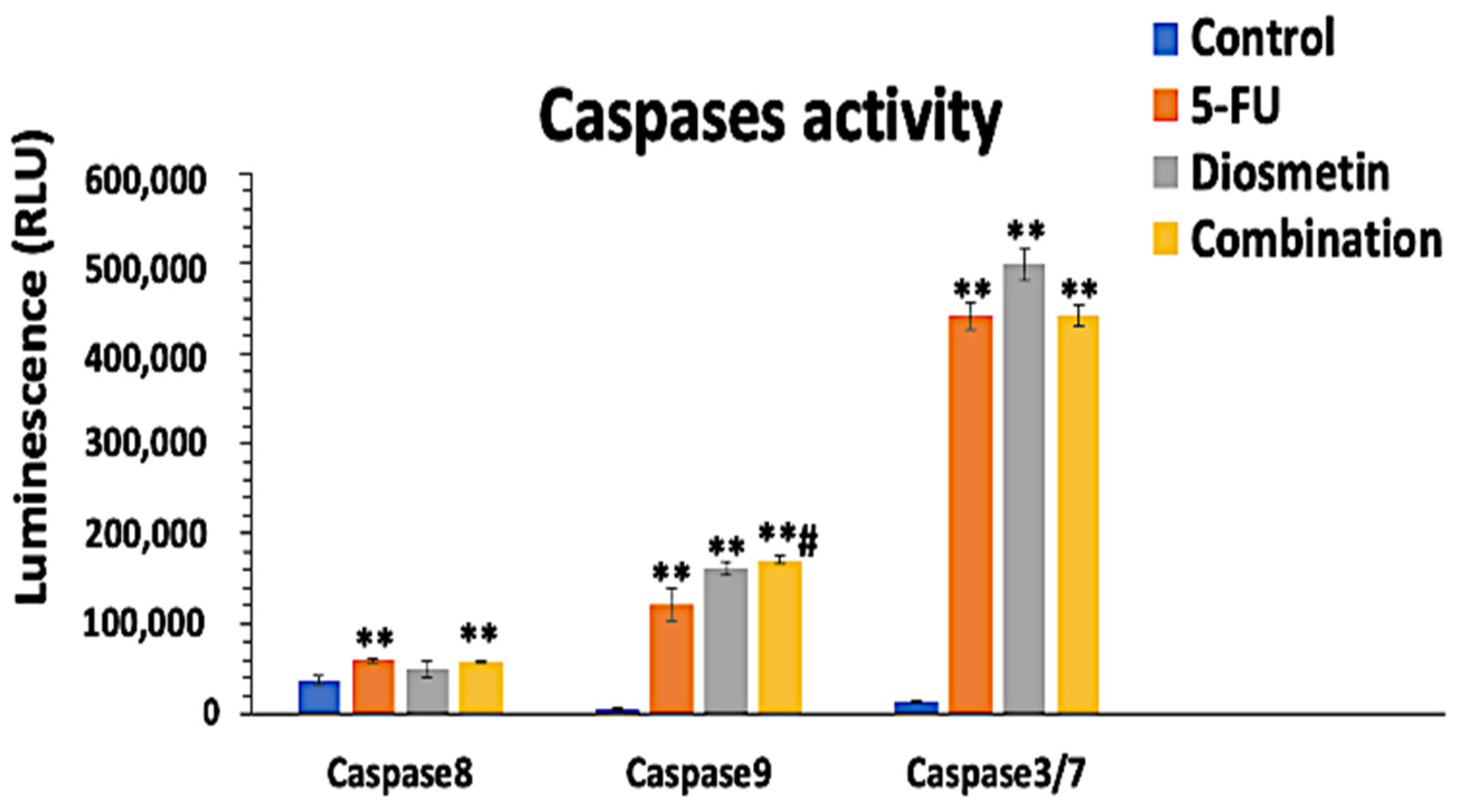
2.7. Caspases Activity Detection
2.8. Statistical Analysis
3. Results
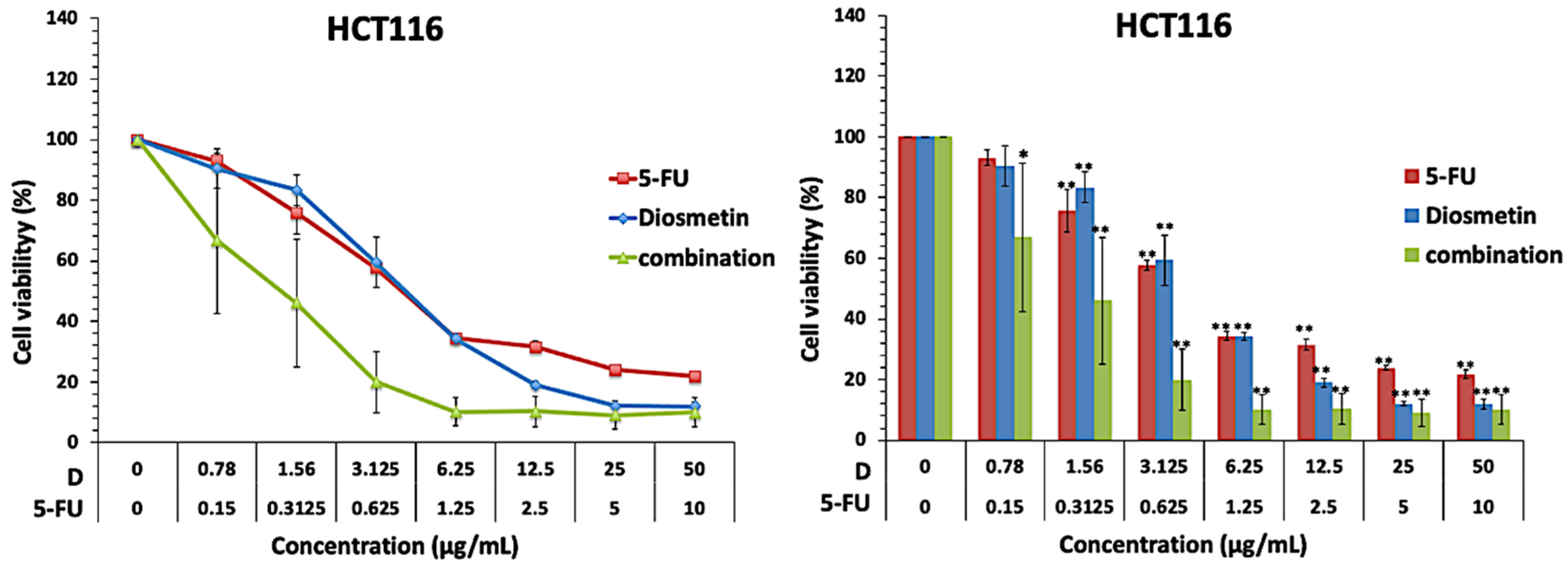
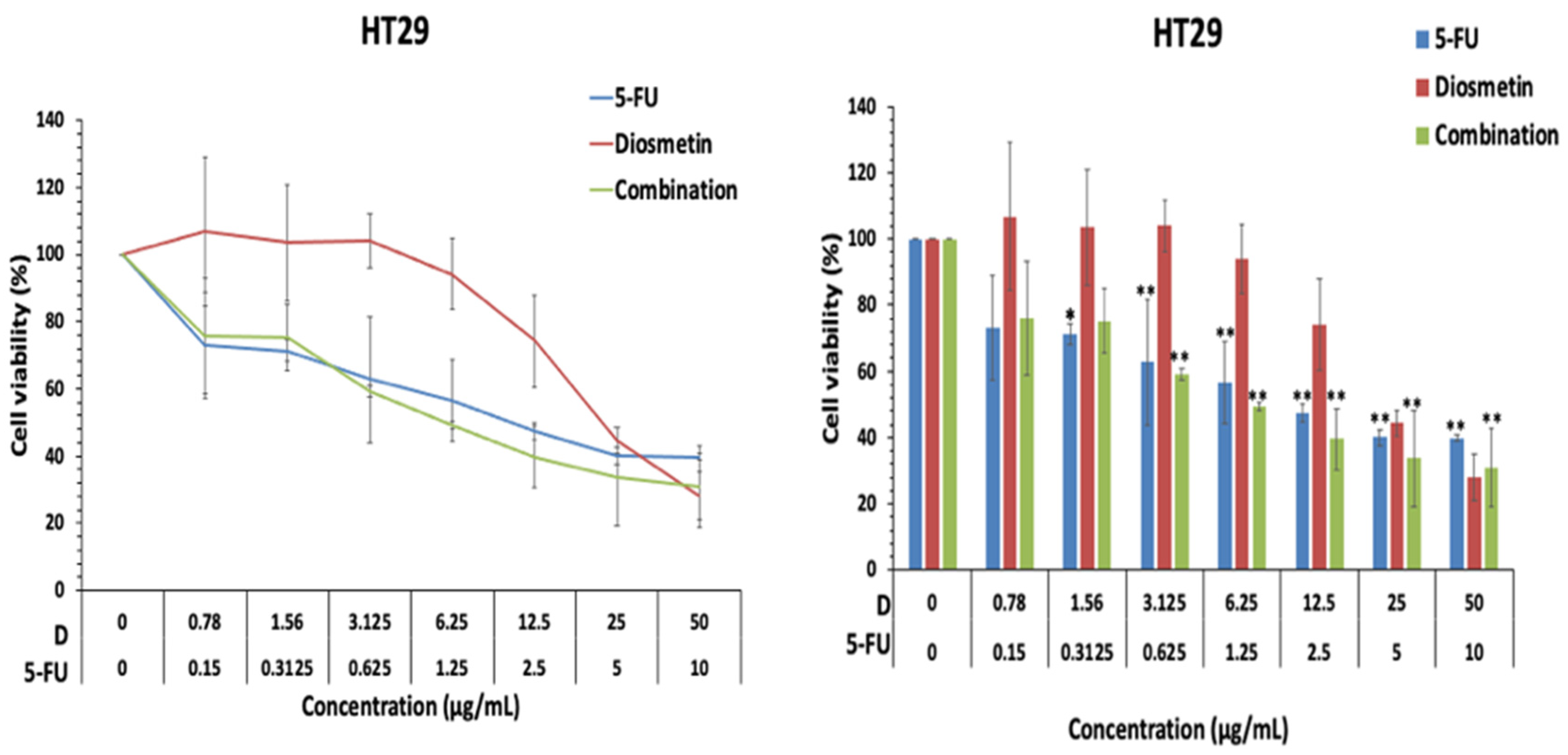
3.1. Effect of Combination on Cell Proliferation
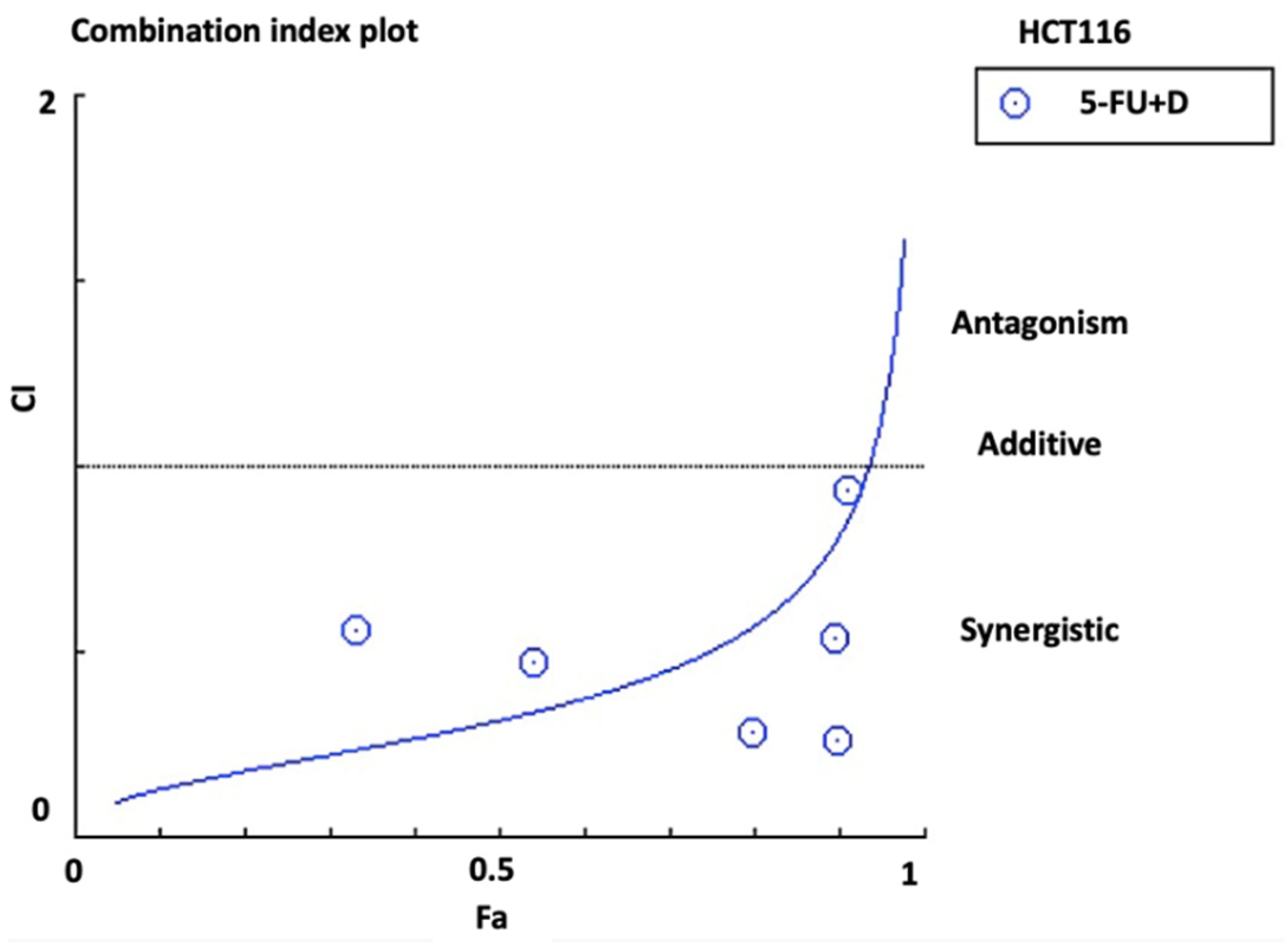
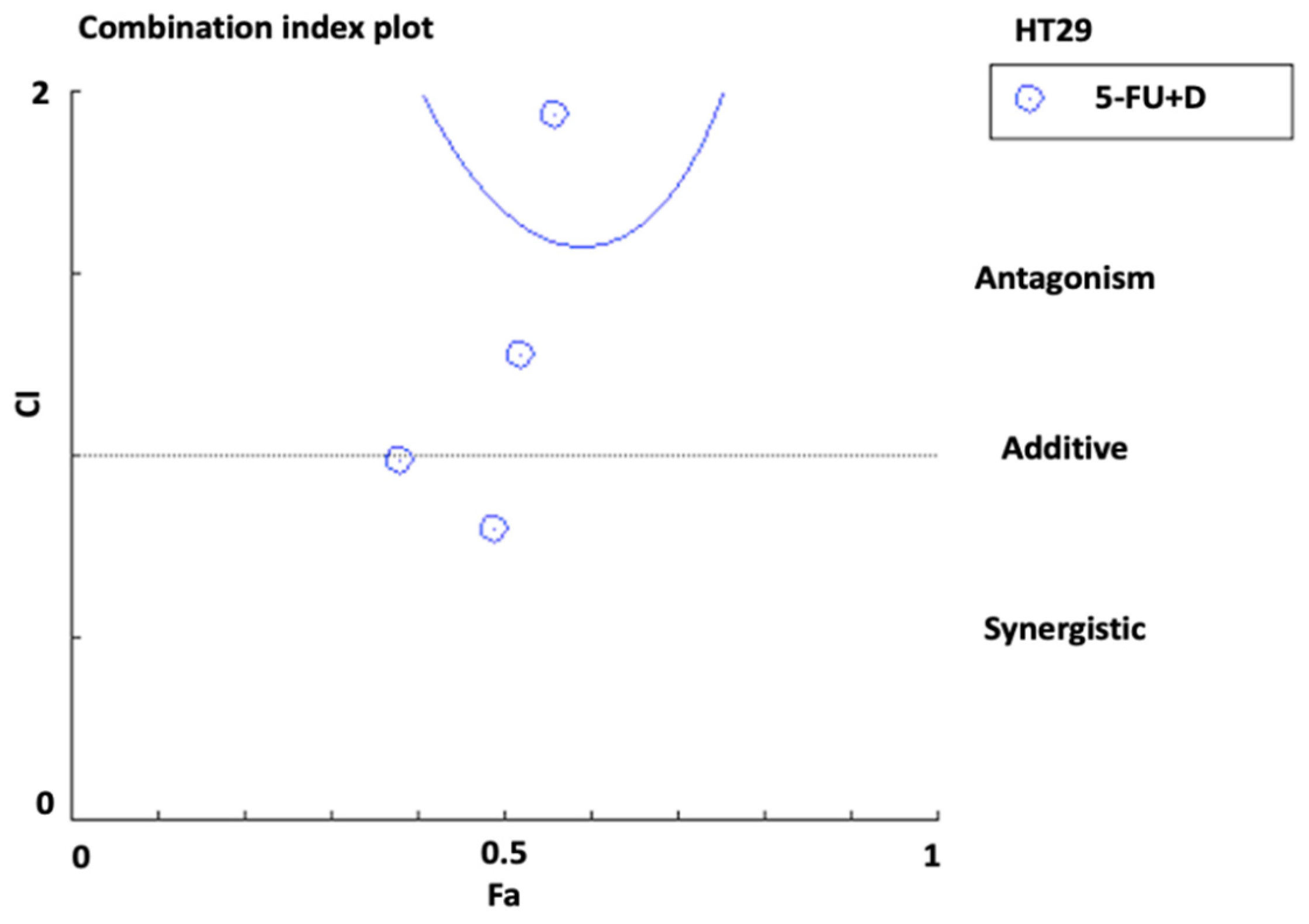
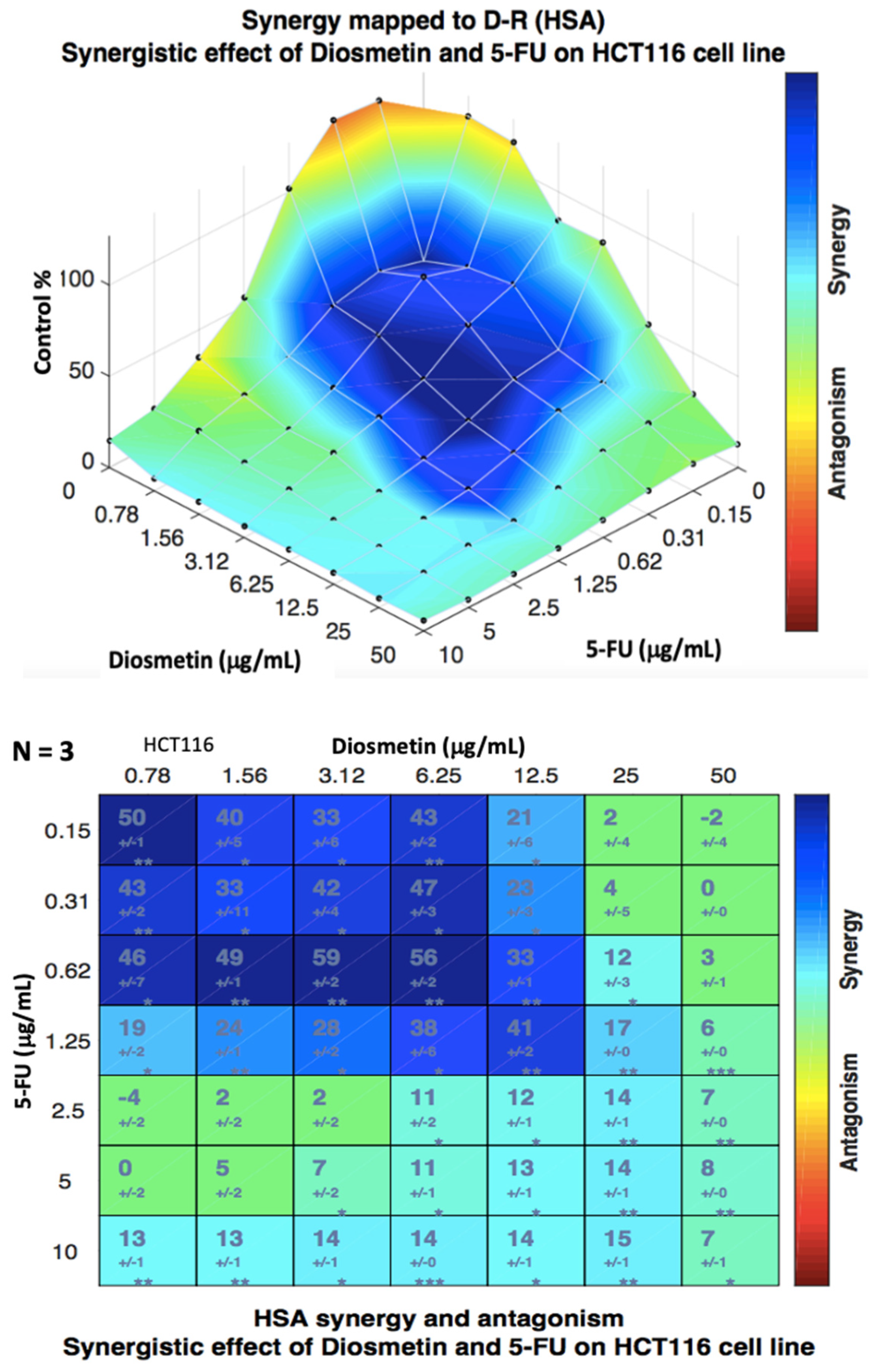
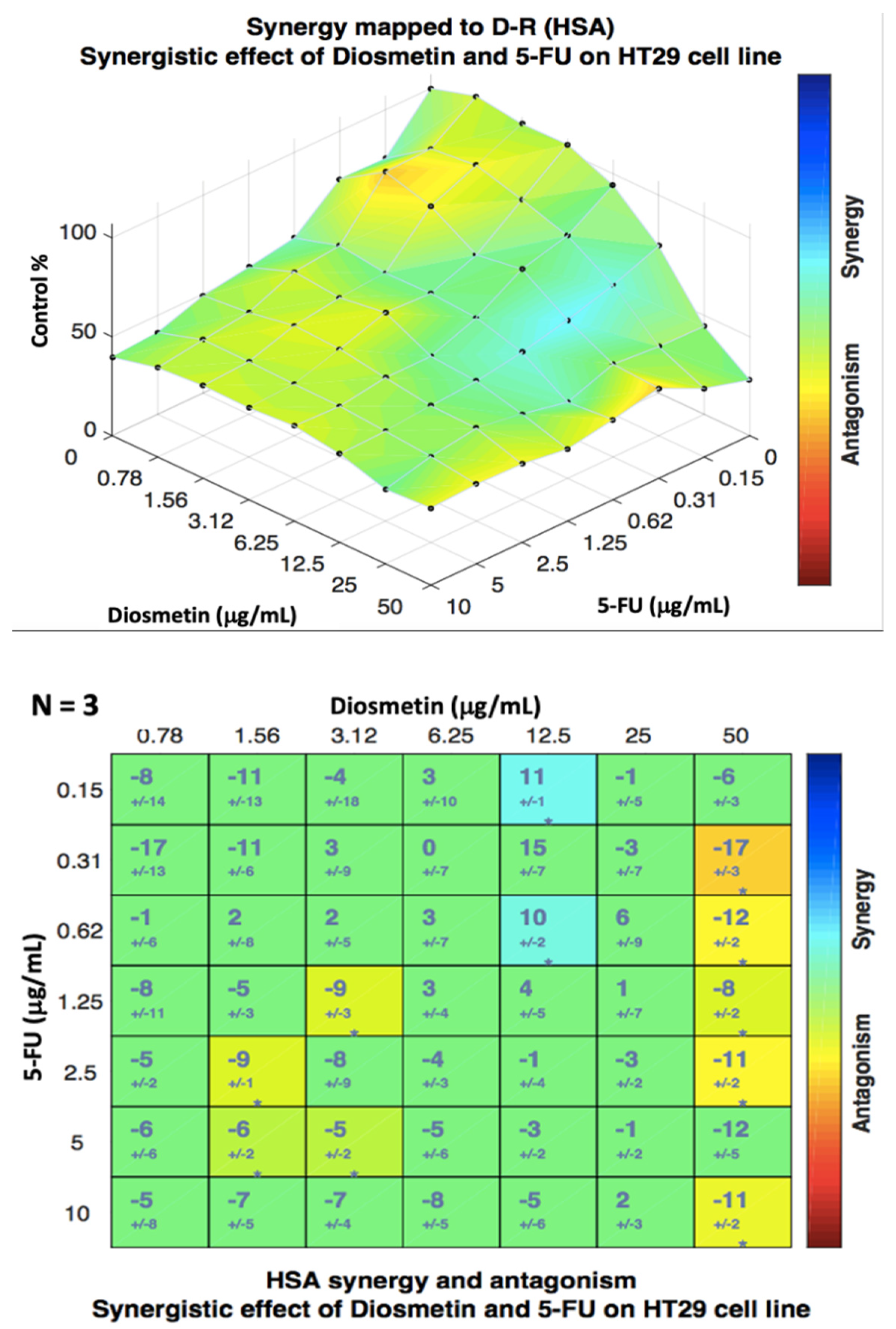
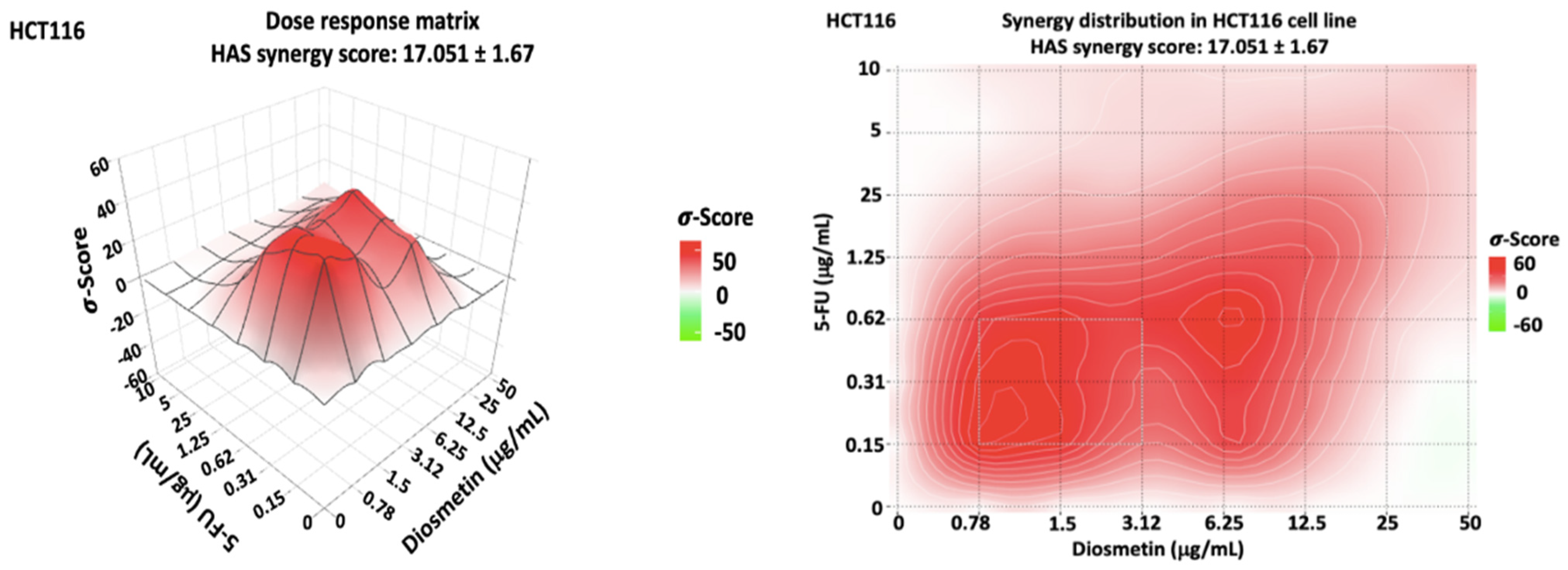
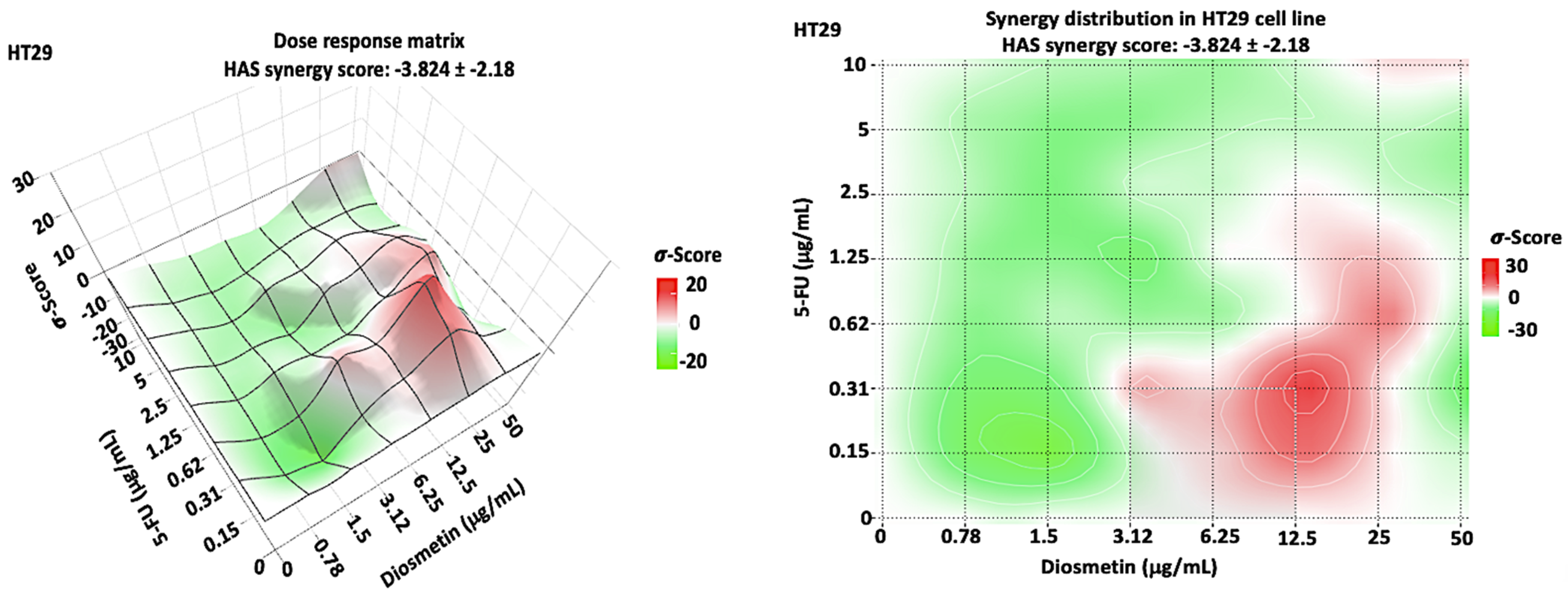
3.2. Synergistic Effect of Combination
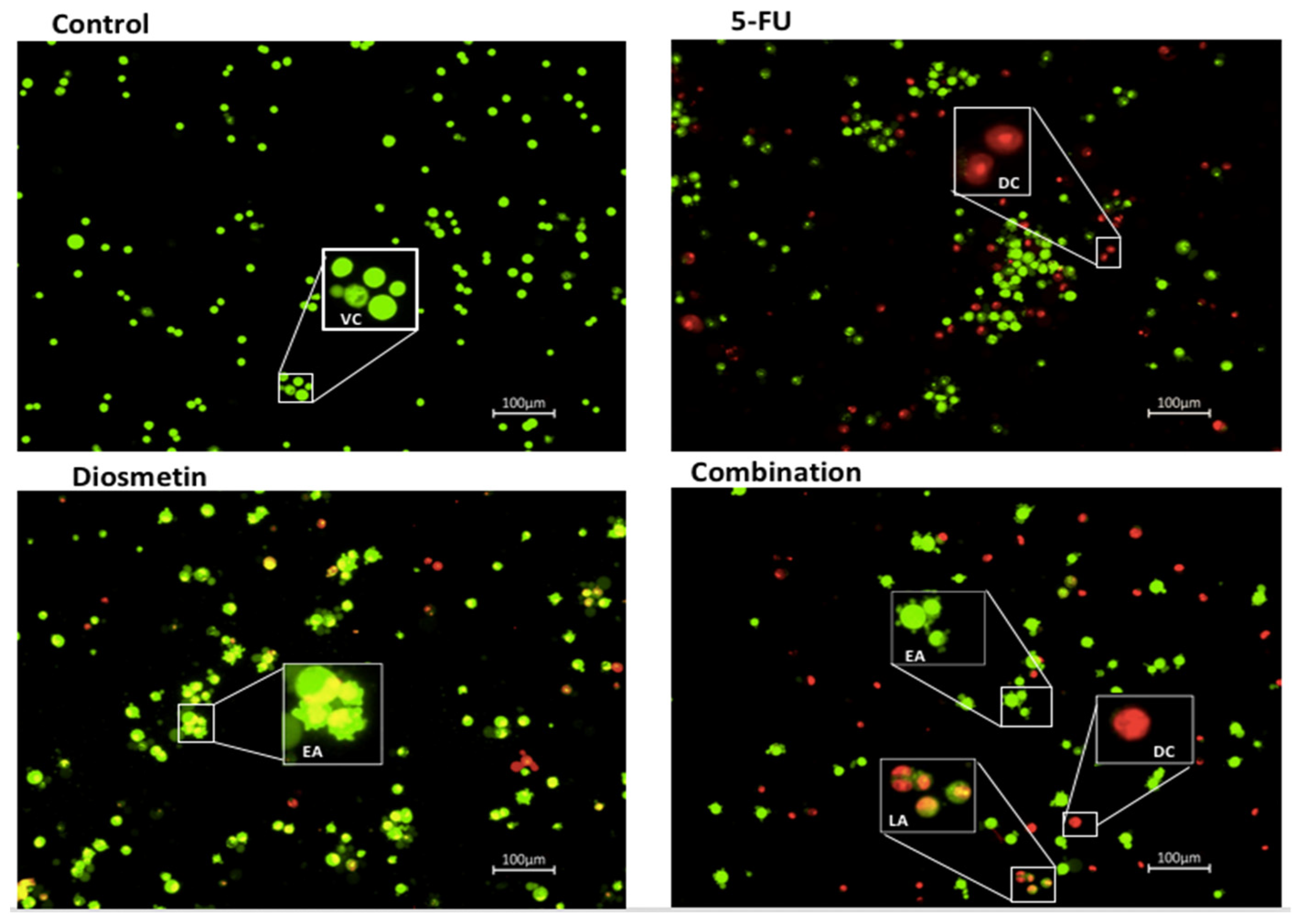
3.3. AO/PI Double Staining Assay
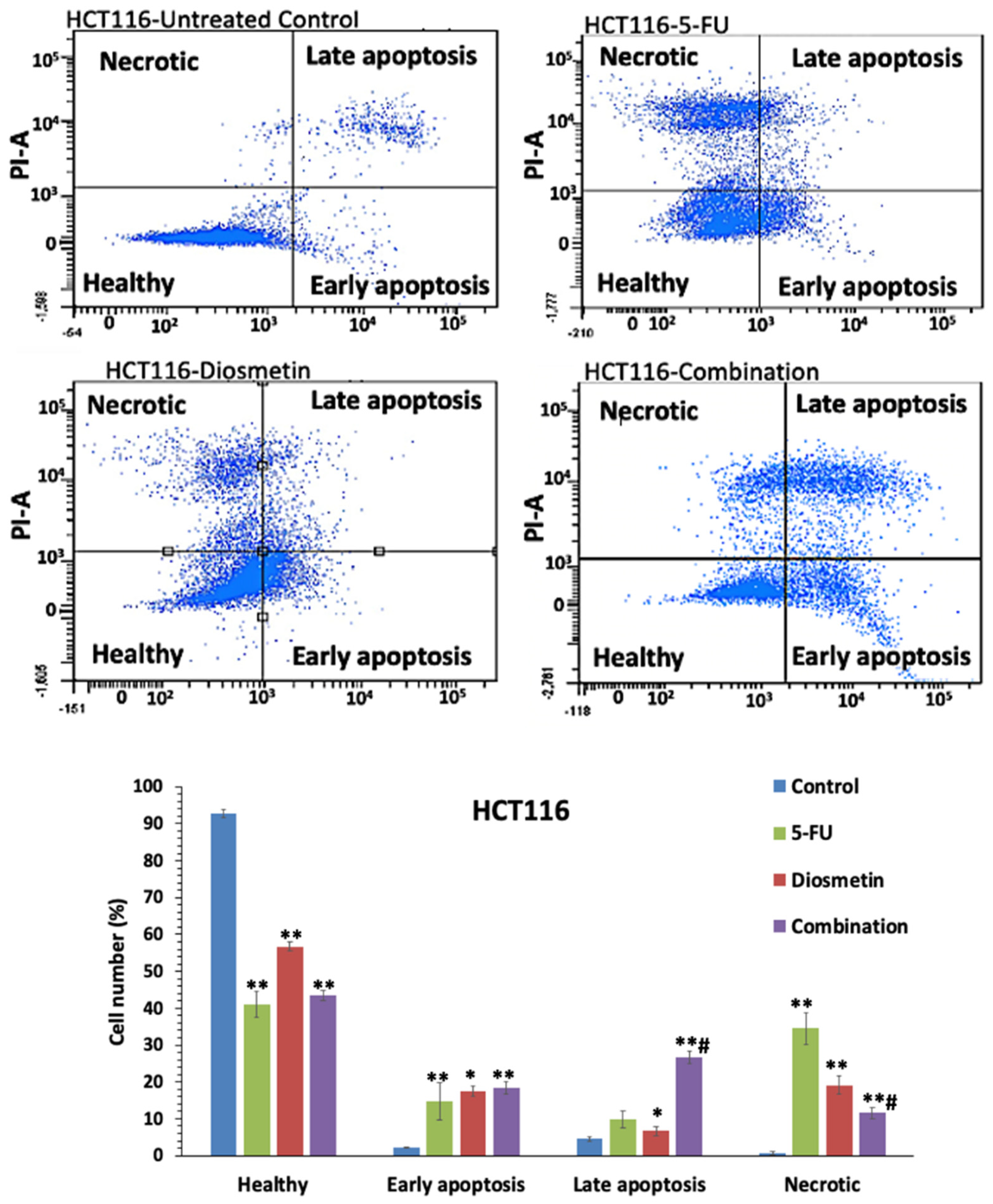
3.4. Annexin V-FITC Assay
3.5. Effect of Combination on Cell Cycle of HCT116
3.6. Effect of Combination on Caspases Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ciardiello, D.; Vitiello, P.P.; Cardone, C.; Martini, G.; Troiani, T.; Martinelli, E.; Ciardiello, F. Immunotherapy of colorectal cancer: Challenges for therapeutic efficacy. Cancer Treat. Rev. 2019, 76, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Wei, L.; Lin, S.; Chen, Y.; Lin, J.; Peng, J. Synergistic effect of kaempferol and 5-fluorouracil on the growth of colorectal cancer cells by regulating the PI3K/Akt signaling pathway. Mol. Med. Rep. 2019, 20, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Ţigu, A.B.; Toma, V.A.; Mot, A.C.; Jurj, A.; Moldovan, C.S.; Fischer-Fodor, E.; Berindan-Neagoe, I.; Pârvu, M. The synergistic antitumor effect of 5-fluorouracil combined with allicin against lung and colorectal carcinoma cells. Molecules 2020, 25, 1947. [Google Scholar] [CrossRef] [PubMed]
- Milczarek, M.; Pogorzelska, A.; Wiktorska, K. Synergistic interaction between 5-fu and an analog of sulforaphane—2-oxohexyl isothiocyanate—in an in vitro colon cancer model. Molecules 2021, 26, 3019. [Google Scholar] [CrossRef]
- Chen, S.J.; Chung, Y.C.; Chang, H.L.; Chang, H.P.; Chou, J.L.; Lin, C.C.; Chen, C.H.; Hsu, C.P. Synergistic Effect of Combined Treatment with Longan Flower Extract and 5-Fluorouracil on Colorectal Cancer Cells. Nutr. Cancer 2020, 72, 209–217. [Google Scholar] [CrossRef]
- Dehghan, R.; Bahreini, F.; Najafi, R.; Saidijam, M.; Amini, R. The Combination of Zerumbone and 5-FU: A Significant Therapeutic Strategy in Sensitizing Colorectal Cancer Cells to Treatment. BioMed Res. Int. 2021, 2021, 6635874. [Google Scholar] [CrossRef]
- Ghosh, S.; Pal, A.; Ray, M. Methylglyoxal in combination with 5-Fluorouracil elicits improved chemosensitivity in breast cancer through apoptosis and cell cycle inhibition. Biomed. Pharmacother. 2019, 114, 108855. [Google Scholar] [CrossRef]
- Xie, Y.H.; Chen, Y.X.; Fang, J.Y. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct. Target. Ther. 2020, 5, 22. [Google Scholar] [CrossRef]
- He, L.; Zhu, H.; Zhou, S.; Wu, T.; Wu, H.; Yang, H.; Mao, H.; SekharKathera, C.; Janardhan, A.; Edick, A.M.; et al. Wnt pathway is involved in 5-FU drug resistance of colorectal cancer cells. Exp. Mol. Med. 2018, 50, 101. [Google Scholar] [CrossRef] [Green Version]
- Kamran, S.; Sinniah, A. Synergistic Effects of 5-Fluorouracil in Combination with Diosmetin in Colorectal Cancer Cells. Biol. Life Sci. Forum 2021, 7, 6. [Google Scholar] [CrossRef]
- Siemiaszko, G.; Niemirowicz-Laskowska, K.; Markiewicz, K.H.; Misztalewska-Turkowicz, I.; Dudź, E.; Milewska, S.; Misiak, P.; Kurowska, I.; Sadowska, A.; Car, H.; et al. Synergistic effect of folate-conjugated polymers and 5-fluorouracil in the treatment of colon cancer. Cancer Nanotechnol. 2021, 12, 31. [Google Scholar] [CrossRef]
- Koosha, S.; Mohamed, Z.; Sinniah, A.; Alshawsh, M.A. Investigation into the Molecular Mechanisms underlying the Anti-proliferative and Anti-tumorigenesis activities of Diosmetin against HCT-116 Human Colorectal Cancer. Sci. Rep. 2019, 9, 5148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Aimaiti, M.; Su, D.; Miao, W.; Zhou, B.; Maimaitiyiming, D.; Yusup, A.; Upur, H.; Aikemu, A. Enhanced efficacy with reduced toxicity of chemotherapy drug 5-fluorouracil by synergistic treatment with Abnormal Savda Munziq from Uyghur medicine. BMC Complement. Altern. Med. 2017, 17, 201. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.C. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–447. [Google Scholar] [CrossRef] [Green Version]
- Chamberlin, S.R.; Blucher, A.; Wu, G.; Shinto, L.; Choonoo, G.; Kulesz-Martin, M.; McWeeney, S. Natural product target network reveals potential for cancer combination therapies. Front. Pharmacol. 2019, 10, 557. [Google Scholar] [CrossRef]
- Kogan, L.G.; Davis, S.L.; Brooks, G.A. Treatment delays during FOLFOX chemotherapy in patients with colorectal cancer: A multicenter retrospective analysis. J. Gastrointest. Oncol. 2019, 10, 841–846. [Google Scholar] [CrossRef]
- Salehifar, E.; Avan, R.; Janbabaei, G.; Mousavi, S.K.; Faramarzi, F. Comparison the incidence and severity of side effects profile of folfox and dcf regimens in gastric cancer patients. Iran. J. Pharm. Res. 2019, 18, 1032–1039. [Google Scholar] [CrossRef]
- Chen, H.M.; Lai, Z.Q.; Liao, H.J.; Xie, J.H.; Xian, Y.F.; Chen, Y.L.; Ip, S.P.; Lin, Z.X.; Su, Z.R. Synergistic antitumor effect of brusatol combined with cisplatin on colorectal cancer cells. Int. J. Mol. Med. 2018, 41, 1447–1454. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wu, Q.; Chen, Y.; Zhang, J.; Li, H.; Yang, Z.; Yang, Y.; Deng, Y.; Zhang, L.; Liu, B. Diosmetin induces apoptosis and enhances the chemotherapeutic efficacy of paclitaxel in non-small cell lung cancer cells via Nrf2 inhibition. Br. J. Pharmacol. 2019, 176, 2079–2094. [Google Scholar] [CrossRef]
- Androutsopoulos, V.P.; Spandidos, D.A. The flavonoids diosmetin and luteolin exert synergistic cytostatic effects in human hepatoma HepG2 cells via CYP1A-catalyzed metabolism, activation of JNK and ERK and P53/P21 up-regulation. J. Nutr. Biochem. 2013, 24, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Kosakowska-Cholody, T.; Cholody, W.; Hariprakasha, H.; Meyer, C.; Michejda, C. Gene expression profiles in HCT116 and HT29 cells exposed to RTA 502 lead to insights into the mechanism of action. Exp. Mol. Ther. 2007, 67, 4895. [Google Scholar]
- Yadav, B.; Wennerberg, K.; Aittokallio, T.; Tang, J. Searching for Drug Synergy in Complex Dose-Response Landscapes Using an Interaction Potency Model. Comput. Struct. Biotechnol. J. 2015, 13, 504–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroemer, G.; Reed, J.C. Mitochondrial control of cell death. Nat. Med. 2000, 6, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.C. Warner-Lambert/Parke Davis award lecture: Mechanisms of apoptosis. Am. J. Pathol. 2000, 157, 1415–1430. [Google Scholar] [CrossRef]
- Fadok, V.A.; De Cathelineau, A.; Daleke, D.L.; Henson, P.M.; Bratton, D.L. Loss of phospholipid asymmetry and surface exposure of phosphatidylserine is required for phagocytosis of apoptotic cells by macrophages and fibroblasts. J. Biol. Chem. 2001, 276, 1071–1077. [Google Scholar] [CrossRef] [Green Version]
- Vermes, I.; Haanen, C.; Steffens-Nakken, H.; Reutellingsperger, C. A novel assay for apoptosis Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled Annexin V. J. Immunol. Methods 1995, 184, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Ricci, M.S.; Zong, W.-X. Chemotherapeutic Approaches for Targeting Cell Death Pathways. Oncologist 2006, 11, 342–357. [Google Scholar] [CrossRef] [Green Version]
- Nouri, Z.; Fakhri, S.; Nouri, K.; Wallace, C.E.; Farzaei, M.H.; Bishayee, A. Targeting multiple signaling pathways in cancer: The rutin therapeutic approach. Cancers 2020, 12, 2276. [Google Scholar] [CrossRef]
- Zhang, T.; Ma, Y.; Fang, J.; Liu, C.; Chen, L. A Deregulated PI3K-AKT Signaling Pathway in Patients with Colorectal Cancer. J. Gastrointest. Cancer 2019, 50, 35–41. [Google Scholar] [CrossRef]
- Qiao, J.; Liu, J.; Jia, K.; Li, N.; Liu, B.; Zhang, Q.; Zhu, R. Diosmetin triggers cell apoptosis by activation of the p53/Bcl-2 pathway and inactivation of the notch3/Nf-κB pathway in HepG2 cells. Oncol. Lett. 2016, 12, 5122–5128. [Google Scholar] [CrossRef] [PubMed]
- Morfoisse, F.; Kuchnio, A.; Frainay, C.; Gomez-Brouchet, A.; Delisle, M.B.; Marzi, S.; Helfer, A.C.; Hantelys, F.; Pujol, F.; Guillermet-Guibert, J.; et al. Hypoxia induces VEGF-C expression in metastatic tumor cells via a HIF-1α-independent translation-mediated mechanism. Cell Rep. 2014, 6, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Ruan, W.J.; Lai, M.D. Autocrine stimulation in colorectal carcinoma (CRC): Positive autocrine loops in human colorectal carcinoma and applicable significance of blocking the loops. Med. Oncol. 2004, 21, 1–8. [Google Scholar] [CrossRef]
- Xia, H.; Shen, J.; Chen, S.; Huang, H.; Xu, Y.; Ma, H. Overexpression of VEGF-C correlates with a poor prognosis in esophageal cancer patients. Cancer Biomarkers 2016, 17, 165–170. [Google Scholar] [CrossRef]
- Mokhtari, R.B.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Narayanaswamy, R.; Ren, H.; Torchilin, V.P. Combination therapy targeting both cancer stem-like cells and bulk tumor cells for improved efficacy of breast cancer treatment. Cancer Biol. Ther. 2016, 17, 698–707. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Wang, F.; Chen, G.; Zhang, H.; Feng, L.; Wang, L.; Colman, H.; Keating, M.J.; Li, X.; Xu, R.H.; et al. Effective elimination of cancer stem cells by a novel drug combination strategy. Stem Cells 2013, 31, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Eyler, C.E.; Rich, J.N. Survival of the fittest: Cancer stem cells in therapeutic resistance and angiogenesis. J. Clin. Oncol. 2008, 26, 2839–2845. [Google Scholar] [CrossRef] [Green Version]
- Milczarek, M.; Mielczarek, L.; Lubelska, K.; Dąbrowska, A.; Chilmonczyk, Z.; Matosiuk, D.; Wiktorska, K. In vitro evaluation of sulforaphane and a natural analog as potent inducers of 5-fluorouracil anticancer activity. Molecules 2018, 23, 3040. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Shen, W.; Zhang, F.; Fan, H.; Tan, J.; Li, L.; Xu, C.; Zhang, H.; Yang, Y.; Cheng, H. α -Hederin Arrests Cell Cycle at G2/M Checkpoint and Promotes Mitochondrial Apoptosis by Blocking Nuclear Factor- B Signaling in Colon Cancer Cells. Biomed Res. Int. 2018, 2018, 2548378. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, T.; Sun, W.; Wang, Z.; Zuo, D.; Zhou, Z.; Li, S.; Xu, J.; Yin, F.; Hua, Y.; et al. Erianin induces G2/M-phase arrest, apoptosis, and autophagy via the ROS/JNK signaling pathway in human osteosarcoma cells in vitro and in vivo. Cell Death Dis. 2016, 7, e2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stark, G.R.; Taylor, W.R. Analyzing the G2/M checkpoint. In Checkpoint Controls and Cancer; Methods in Molecular Biology™; Humana Press: Totowa, NJ, USA, 2004; Volume 280, pp. 51–82. [Google Scholar] [CrossRef]
- Tummers, B.; Green, D.R. Caspase-8: Regulating life and death. Immunol. Rev. 2017, 277, 76–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, E.; Chiaradia, L.D.; Silva, A.H.; Nunes, R.J.; Yunes, R.A.; Creczynski-Pasa, T.B. Involvement of extrinsic and intrinsic apoptotic pathways together with endoplasmic reticulum stress in cell death induced by naphthylchalcones in a leukemic cell line: Advantages of multi-target action. Toxicol. Vitr. 2014, 28, 769–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Drug | HCT116 Cells IC50 (μg/mL) | HT29 Cells IC50 (μg/mL) |
|---|---|---|---|
| Monotherapy | Diosmetin | 4.16 ± 1.3 | 22.06 ± 2.7 |
| 5-FU | 0.83 ± 0.0 | 1.65 ± 0.8 | |
| Combination therapy | Diosmetin | 1.38 ± 0.8 | 6.03 ± 0.3 |
| 5-FU | 0.27 ± 1.1 | 1.20 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamran, S.; Sinniah, A.; Chik, Z.; Alshawsh, M.A. Diosmetin Exerts Synergistic Effects in Combination with 5-Fluorouracil in Colorectal Cancer Cells. Biomedicines 2022, 10, 531. https://doi.org/10.3390/biomedicines10030531
Kamran S, Sinniah A, Chik Z, Alshawsh MA. Diosmetin Exerts Synergistic Effects in Combination with 5-Fluorouracil in Colorectal Cancer Cells. Biomedicines. 2022; 10(3):531. https://doi.org/10.3390/biomedicines10030531
Chicago/Turabian StyleKamran, Sareh, Ajantha Sinniah, Zamri Chik, and Mohammed Abdullah Alshawsh. 2022. "Diosmetin Exerts Synergistic Effects in Combination with 5-Fluorouracil in Colorectal Cancer Cells" Biomedicines 10, no. 3: 531. https://doi.org/10.3390/biomedicines10030531
APA StyleKamran, S., Sinniah, A., Chik, Z., & Alshawsh, M. A. (2022). Diosmetin Exerts Synergistic Effects in Combination with 5-Fluorouracil in Colorectal Cancer Cells. Biomedicines, 10(3), 531. https://doi.org/10.3390/biomedicines10030531