
Dysregulation of lncRNA–miRNA–mRNA Interactome as a Marker of Metastatic Process in Ovarian Cancer

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
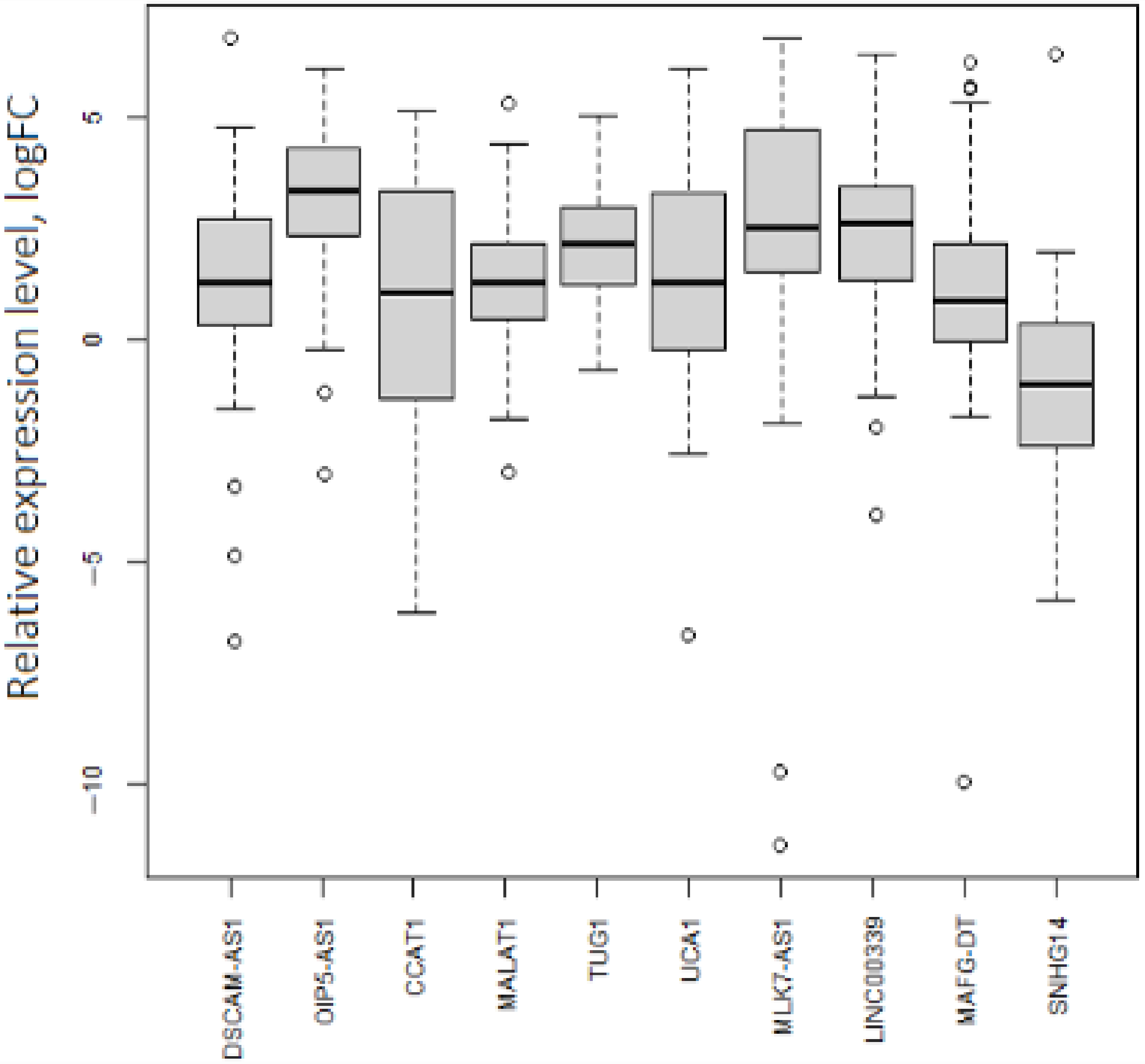
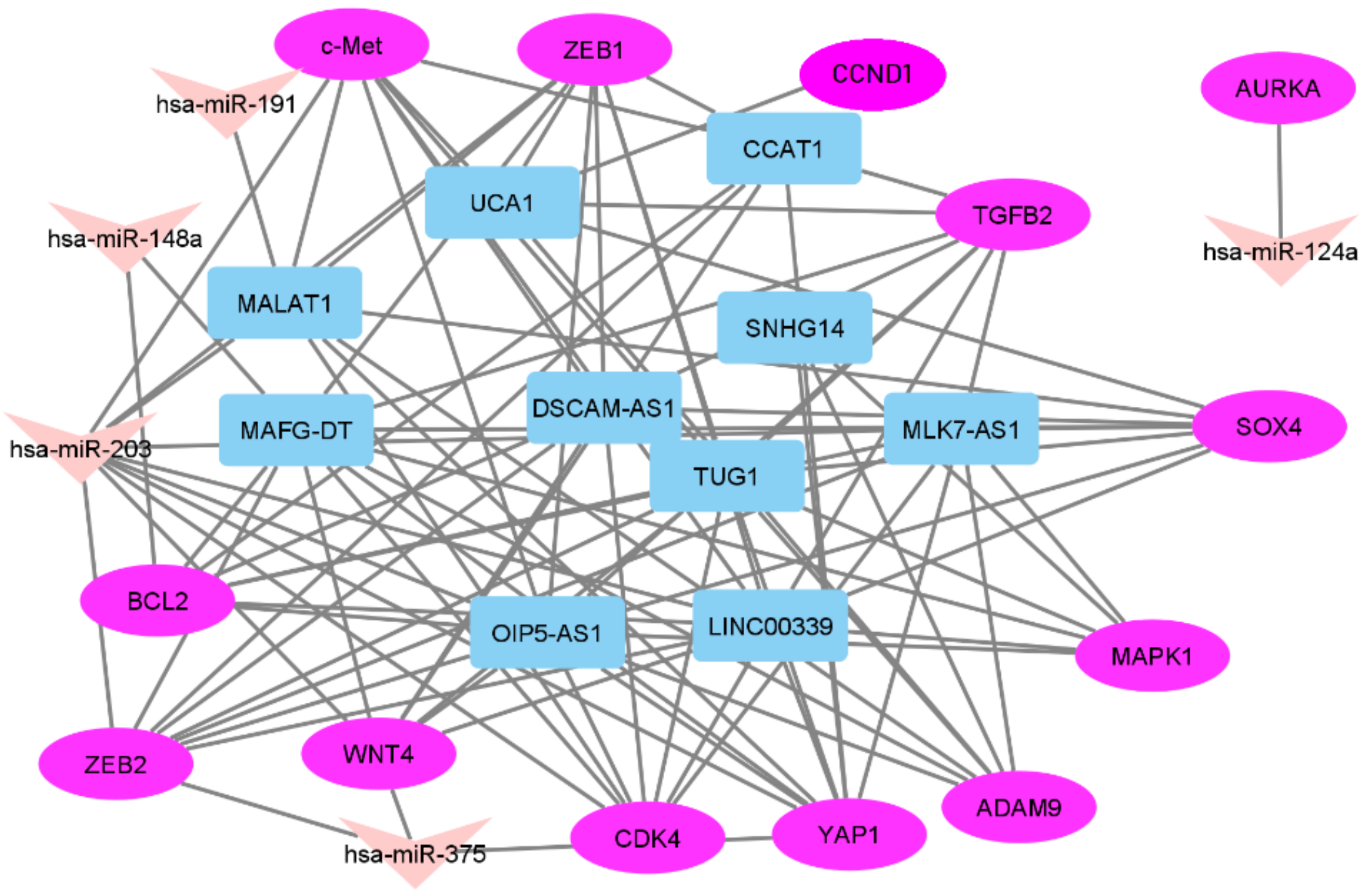
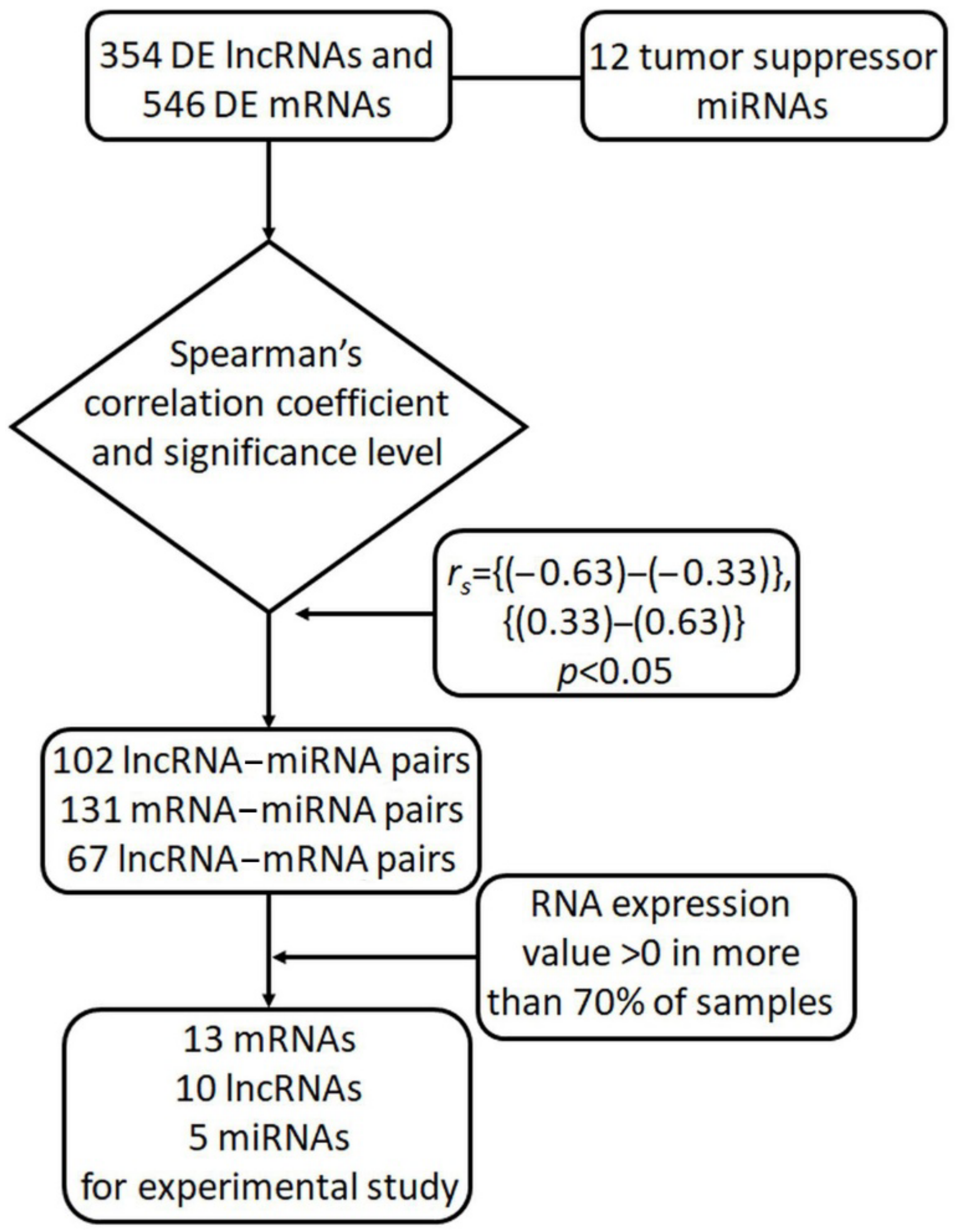
2.1. In Silico Selection of lncRNAs and mRNAs
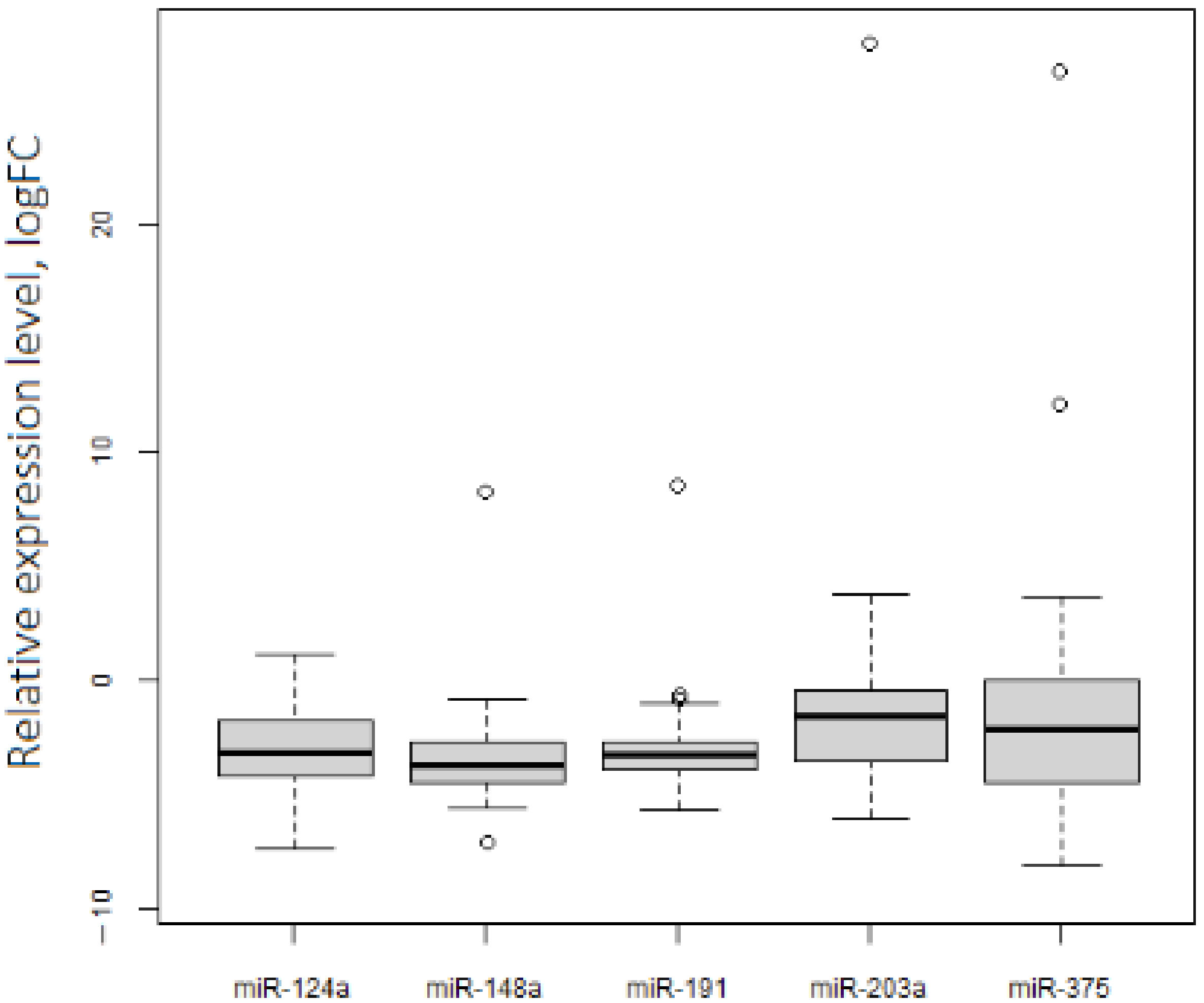
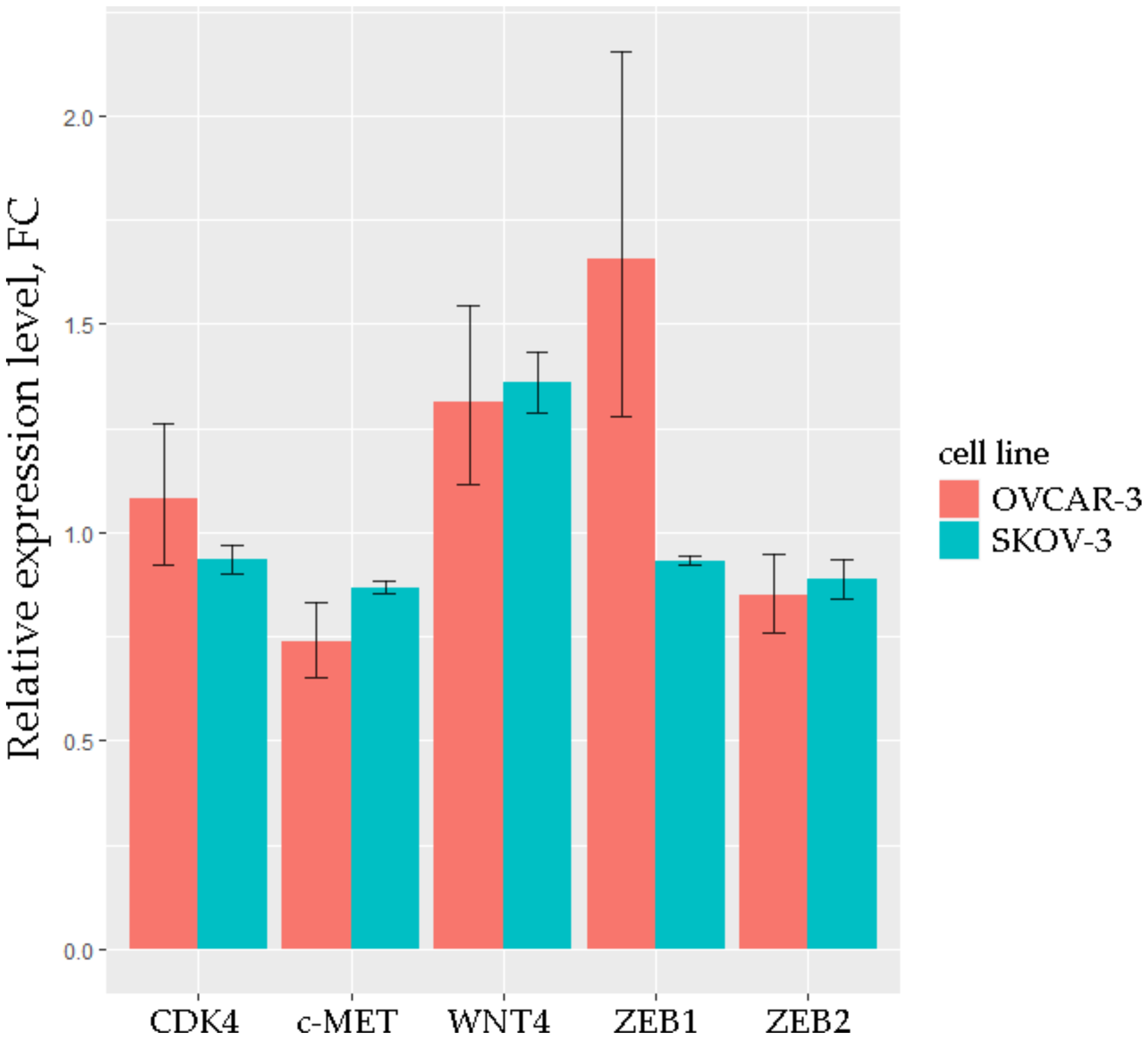
2.2. Experimental Studies of the Level of Selected RNAs on Samples from OC Patients
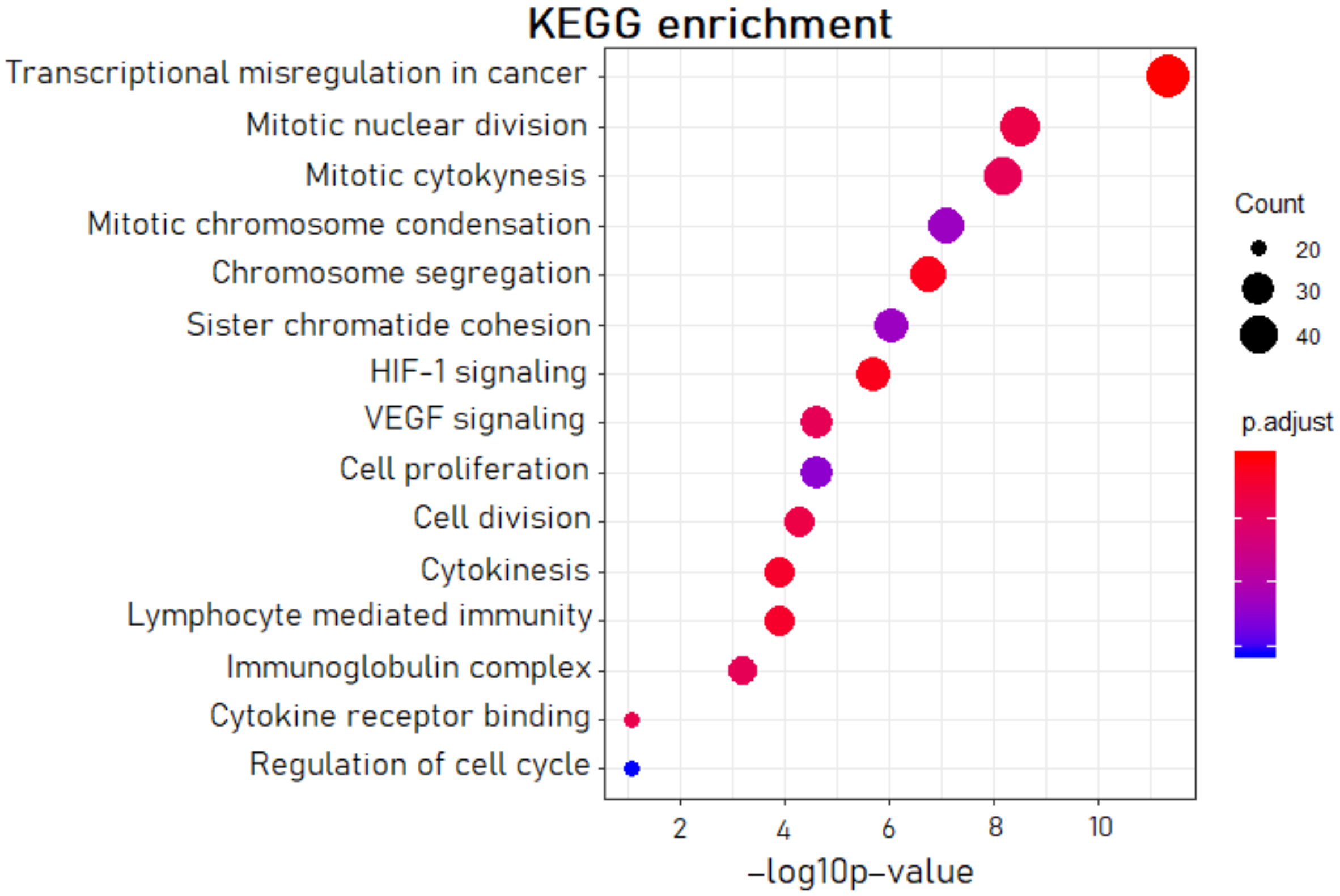
2.3. Experimental Microarray-Based Expression Analysis on Clinical Samples
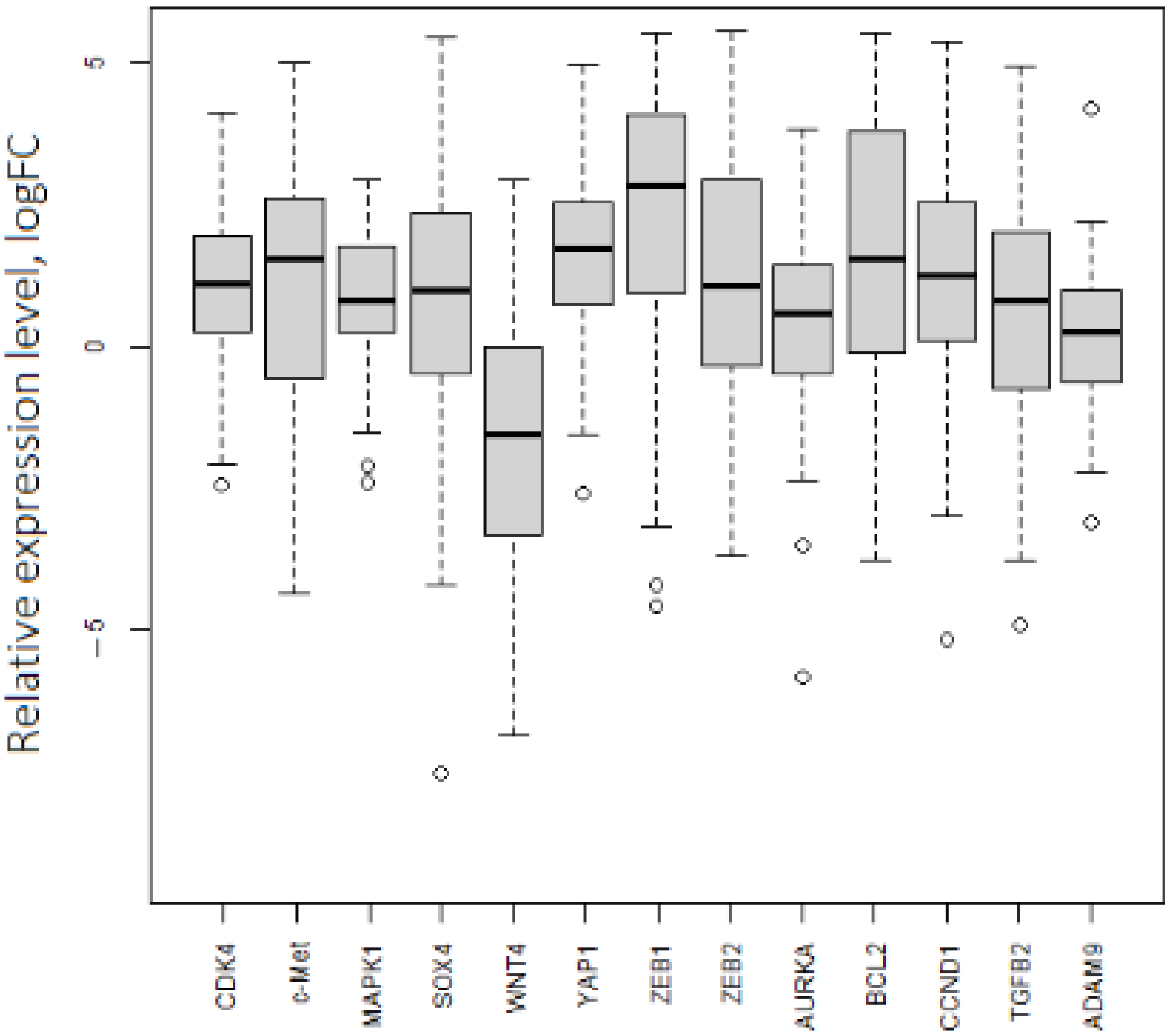
2.4. Experimental Validation of the Interactions between miRNAs and mRNAs on Cell Lines
3. Discussion
4. Materials and Methods
4.1. Bioinformatic Analysis
4.2. Clinical Data
4.3. RNA Isolation
4.4. Reverse Transcription and qPCR
4.5. Cell Lines
4.6. RNA Duplexes and Transfection
4.7. Affymetrix Whole-Transcriptome Gene Expression Analysis
4.8. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carioli, G.; Malvezzi, M.; Bertuccio, P.; Levi, F.; Boffetta, P.; Negri, E.; La Vecchia, C. Cancer mortality and predictions for 2018 in selected Australasian countries and Russia. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2019, 30, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tollefsbol, T.O. DNA methylation methods: Global DNA methylation and methylomic analyses. Methods 2021, 187, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Center, M.M.; DeSantis, C.; Ward, E.M. Global patterns of cancer incidence and mortality rates and trends. Cancer Epidemiol. Prev. Biomark. 2010, 19, 1893–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Luo, G.; Li, M.; Guo, P.; Xiao, Y.; Ji, H.; Hao, Y. Global patterns and trends in ovarian cancer incidence: Age, period and birth cohort analysis. BMC Cancer 2019, 19, 984. [Google Scholar] [CrossRef] [PubMed]
- Malvezzi, M.; Carioli, G.; Rodriguez, T.; Negri, E.; La Vecchia, C. Global trends and predictions in ovarian cancer mortality. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2016, 27, 2017–2025. [Google Scholar] [CrossRef]
- Rauh-Hain, J.A.; Krivak, T.C.; Del Carmen, M.G.; Olawaiye, A.B. Ovarian cancer screening and early detection in the general population. Rev. Obstet. Gynecol. 2011, 4, 15–21. [Google Scholar]
- Smith, C.G. A Resident’s Perspective of Ovarian Cancer. Diagnostics 2017, 7, 24. [Google Scholar] [CrossRef]
- Yang, Z.; Xu, F.; Wang, H.; Teschendorff, A.E.; Xie, F.; He, Y. Pan-cancer characterization of long non-coding RNA and DNA methylation mediated transcriptional dysregulation. EBioMedicine 2021, 68, 103399. [Google Scholar] [CrossRef]
- Kumar, S.; Gonzalez, E.A.; Rameshwar, P.; Etchegaray, J.P. Non-Coding RNAs as Mediators of Epigenetic Changes in Malignancies. Cancers 2020, 12, 3657. [Google Scholar] [CrossRef]
- Saw, P.E.; Xu, X.; Chen, J.; Song, E.W. Non-coding RNAs: The new central dogma of cancer biology. Sci. China Life Sci. 2021, 64, 22–50. [Google Scholar] [CrossRef]
- Miano, V.; Codino, A.; Pandolfini, L.; Barbieri, I. The non-coding epitranscriptome in cancer. Brief. Funct. Genom. 2021, 20, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Loginov, V.I.; Pronina, I.V.; Burdennyy, A.M.; Filippova, E.A.; Kazubskaya, T.P.; Kushlinsky, D.N.; Utkin, D.O.; Khodyrev, D.S.; Kushlinskii, N.E.; Dmitriev, A.A.; et al. Novel miRNA genes deregulated by aberrant methylation in ovarian carcinoma are involved in metastasis. Gene 2018, 662, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Li, J.; Zhao, P. Roles of ncRNAs as ceRNAs in Gastric Cancer. Genes 2021, 12, 1036. [Google Scholar] [CrossRef] [PubMed]
- Su, K.; Wang, N.; Shao, Q.; Liu, H.; Zhao, B.; Ma, S. The role of a ceRNA regulatory network based on lncRNA MALAT1 site in cancer progression. Biomed. Pharmacother. 2021, 137, 111389. [Google Scholar] [CrossRef]
- Braga, E.A.; Fridman, M.V.; Moscovtsev, A.A.; Filippova, E.A.; Dmitriev, A.A.; Kushlinskii, N.E. LncRNAs in Ovarian Cancer Progression, Metastasis, and Main Pathways: ceRNA and Alternative Mechanisms. Int. J. Mol. Sci. 2020, 21, 8855. [Google Scholar] [CrossRef]
- Ala, U. Competing endogenous RNAs and cancer: How coding and non-coding molecules cross-talk can impinge on disease. Int. J. Biochem. Cell Biol. 2021, 130, 105874. [Google Scholar] [CrossRef]
- Huang, M.; Long, Y.; Jin, Y.; Ya, W.; Meng, D.; Qin, T.; Su, L.; Zhou, W.; Wu, J.; Huang, C.; et al. Comprehensive analysis of the lncRNA-miRNA-mRNA regulatory network for bladder cancer. Transl. Androl. Urol. 2021, 10, 1286–1301. [Google Scholar] [CrossRef]
- Gao, L.; Zhao, Y.; Ma, X.; Zhang, L. Integrated analysis of lncRNA-miRNA-mRNA ceRNA network and the potential prognosis indicators in sarcomas. BMC Med. Genom. 2021, 14, 67. [Google Scholar] [CrossRef]
- Mittempergher, L. Genomic Characterization of High-Grade Serous Ovarian Cancer: Dissecting Its Molecular Heterogeneity as a Road Towards Effective Therapeutic Strategies. Curr. Oncol. Rep. 2016, 18, 44. [Google Scholar] [CrossRef]
- Labidi-Galy, S.I.; Papp, E.; Hallberg, D.; Niknafs, N.; Adleff, V.; Noe, M.; Bhattacharya, R.; Novak, M.; Jones, S.; Phallen, J.; et al. High grade serous ovarian carcinomas originate in the fallopian tube. Nat. Commun. 2017, 8, 1093. [Google Scholar] [CrossRef] [PubMed]
- Lisio, M.A.; Fu, L.; Goyeneche, A.; Gao, Z.H.; Telleria, C. High-Grade Serous Ovarian Cancer: Basic Sciences, Clinical and Therapeutic Standpoints. Int. J. Mol. Sci. 2019, 20, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loginov, V.I.; Pronina, I.V.; Filippova, E.A.; Burdennyy, A.M.; Lukina, S.S.; Kazubskaya, T.P.; Uroshlev, L.A.; Fridman, M.V.; Brovkina, O.I.; Apanovich, N.V.; et al. Aberrant Methylation of 20 miRNA Genes Specifically Involved in Various Steps of Ovarian Carcinoma Spread: From Primary Tumors to Peritoneal Macroscopic Metastases. Int. J. Mol. Sci. 2022, 23, 1300. [Google Scholar] [CrossRef]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Ricciardi, M.; Zanotto, M.; Malpeli, G.; Bassi, G.; Perbellini, O.; Chilosi, M.; Bifari, F.; Krampera, M. Epithelial-to-mesenchymal transition (EMT) induced by inflammatory priming elicits mesenchymal stromal cell-like immune-modulatory properties in cancer cells. Br. J. Cancer 2015, 112, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Georgakopoulos-Soares, I.; Chartoumpekis, D.V.; Kyriazopoulou, V.; Zaravinos, A. EMT Factors and Metabolic Pathways in Cancer. Front. Oncol. 2020, 10, 499. [Google Scholar] [CrossRef]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantel, K.; Brakenhoff, R.H. Dissecting the metastatic cascade. Nat. Rev. Cancer 2004, 4, 448–456. [Google Scholar] [CrossRef]
- Van Baal, J.; van Noorden, C.J.F.; Nieuwland, R.; Van de Vijver, K.K.; Sturk, A.; van Driel, W.J.; Kenter, G.G.; Lok, C.A.R. Development of Peritoneal Carcinomatosis in Epithelial Ovarian Cancer: A Review. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2018, 66, 67–83. [Google Scholar] [CrossRef]
- Zhao, G.; Guo, Y.; Chen, Z.; Wang, Y.; Yang, C.; Dudas, A.; Du, Z.; Liu, W.; Zou, Y.; Szabo, E.; et al. miR-203 Functions as a Tumor Suppressor by Inhibiting Epithelial to Mesenchymal Transition in Ovarian Cancer. J. Cancer Sci. Ther. 2015, 7, 34–43. [Google Scholar] [CrossRef]
- Wang, B.; Li, X.; Zhao, G.; Yan, H.; Dong, P.; Watari, H.; Sims, M.; Li, W.; Pfeffer, L.M.; Guo, Y.; et al. miR-203 inhibits ovarian tumor metastasis by targeting BIRC5 and attenuating the TGFbeta pathway. J. Exp. Clin. Cancer Res. 2018, 37, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.T. Epigenetic regulation by long noncoding RNAs. Science 2012, 338, 1435–1439. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Fu, Z.; Gao, L.; Zhou, J.; Deng, X.; Luo, X.; Fang, W.; Luo, R. Direct interaction between miR-203 and ZEB2 suppresses epithelial-mesenchymal transition signaling and reduces lung adenocarcinoma chemoresistance. Acta Biochim. Biophys. Sin. 2016, 48, 1042–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q.; Zhou, Y.; Yang, H.; Li, L.; Deng, X.; Cheng, C.; Xie, Y.; Luo, X.; Fang, W.; Liu, Z. A directly negative interaction of miR-203 and ZEB2 modulates tumor stemness and chemotherapy resistance in nasopharyngeal carcinoma. Oncotarget 2016, 7, 67288–67301. [Google Scholar] [CrossRef] [PubMed]
- Raj, S.; Kesari, K.K.; Kumar, A.; Rathi, B.; Sharma, A.; Gupta, P.K.; Jha, S.K.; Jha, N.K.; Slama, P.; Roychoudhury, S.; et al. Molecular mechanism(s) of regulation(s) of c-MET/HGF signaling in head and neck cancer. Mol. Cancer 2022, 21, 31. [Google Scholar] [CrossRef] [PubMed]
- Park, K.C.; Richardson, D.R. The c-MET oncoprotein: Function, mechanisms of degradation and its targeting by novel anti-cancer agents. Biochim. Et Biophys. Acta Gen. Subj. 2020, 1864, 129650. [Google Scholar] [CrossRef]
- Yang, G.; Fu, Y.; Lu, X.; Wang, M.; Dong, H.; Li, Q. LncRNA HOTAIR/miR-613/c-met axis modulated epithelial-mesenchymal transition of retinoblastoma cells. J. Cell. Mol. Med. 2018, 22, 5083–5096. [Google Scholar] [CrossRef] [Green Version]
- Qu, W.; Chen, X.; Wang, J.; Lv, J.; Yan, D. MicroRNA-1 inhibits ovarian cancer cell proliferation and migration through c-Met pathway. Clin. Chim. Acta Int. J. Clin. Chem. 2017, 473, 237–244. [Google Scholar] [CrossRef]
- Wang, W.; Dong, J.; Wang, M.; Yao, S.; Tian, X.; Cui, X.; Fu, S.; Zhang, S. miR-148a-3p suppresses epithelial ovarian cancer progression primarily by targeting c-Met. Oncol. Lett. 2018, 15, 6131–6136. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Ni, M.; Li, J.; Cheng, J.; Zhao, H.; Zhao, J.; Huang, S.; Wu, X. Circ0004390 promotes cell proliferation through sponging miR-198 in ovarian cancer. Biochem. Biophys. Res. Commun. 2020, 526, 14–20. [Google Scholar] [CrossRef]
- Pronina, I.V.; Loginov, V.I.; Khodyrev, D.S.; Kazubskaya, T.P.; Braga, E.A. RASSF1A expression level in primary epithelial tumors of various locations. Mol. Biol. 2012, 46, 236–243. [Google Scholar] [CrossRef]
- Kroeger, P.T., Jr.; Drapkin, R. Pathogenesis and heterogeneity of ovarian cancer. Curr. Opin. Obstet. Gynecol. 2017, 29, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, K.S.; Blackburn, J.M. Quality assessment and data handling methods for Affymetrix Gene 1.0 ST arrays with variable RNA integrity. BMC Genom. 2013, 14, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNA | Chromosome (Position) * | Localization in a Cell ** | LogFC (TCGA) | LogFC ((qPCR) |
|---|---|---|---|---|
| lncRNAs | ||||
| MALAT1 | Chr11 (65,497,738–65,506,516) | Nucleus | 5.0 | 1.1 |
| MAFG-DT | Chr17 (81,927,829–81,930,753) | Extracellular space | 2.9 | 0.8 |
| OIP5-AS1 | Chr15 (41,282,697–41,313,338) | Nucleus | 5.0 | 3.3 |
| MLK7-AS1 | Chr2 (173,197,712–173,282,036) | No information | 1.5 | 2.5 |
| LINC00339 | Chr1 (22,025,191–22,031,224) | Extracellular space, cytosol, and nucleus | 1.5 | 2.5 |
| TUG1 | Chr22 (30,969,211–30,979,395) | Extracellular space, cytosol, and nucleus | 38.8 | 2.1 |
| UCA1 | Chr19 (15,828,947–15,836,321) | Extracellular space, cytosol, and nucleus | 1.6 | 1.4 |
| SNHG14 | Chr15 (24,823,608–25,419,462) | Nucleus | 2.3 | −1.1 |
| CCAT1 | Chr8 (127,207,382–127,219,268) | Nucleus | 11.6 | 1.3 |
| DSCAM-AS1 | Chr21 (40,383,083–40,385,358) | Extracellular space, cytosol, and nucleus | 27.7 | 1.4 |
| miRNAs | ||||
| miR-148a | Chr7 (25,949,919–25,949,986) | Extracellular space and vesicles | N/A | −4.0 |
| miR-191 | Chr3 (49,020,618–49,020,709) | Extracellular space and vesicles | N/A | −3.8 |
| miR-203a | Chr14 (104,117,405–104,117,514) | Nucleus | N/A | −2.1 |
| miR-124a | Chr8 (9,903,388–9,903,472) | Cytoplasm and extracellular exosomes | N/A | −3.4 |
| miR-375 | Chr2 (219,001,645–219,001,708) | Extracellular space | N/A | −2.5 |
| mRNAs | ||||
| AURKA | Chr20 (56,369,390–56,392,308) | Nucleus | 1.8 | 1.2 |
| BCL2 | Chr18 (63,123,346–63,320,280) | Nucleus | 3.2 | 1.8 |
| CDK4 | Chr12 (57,747,727–57,752,310) | Nucleus | 2.1 | 1.3 |
| c-MET | Chr7 (116,672,196–116,798,386) | Extracellular space | 3.2 | 1.6 |
| WNT4 | Chr1 (22,117,308–22,143,981) | Extracellular space | 2.7 | −1.6 |
| YAP1 | Chr11 (102,109,957–102,233,424) | Nucleus and cytosol | 2.2 | 2.0 |
| ZEB1 | Chr10 (31,318,417–31,529,804) | Nucleus and cytosol | 2.1 | 2.9 |
| ZEB2 | Chr2 (144,384,081–144,520,119) | Nucleus and cytosol | 1.7 | 1.1 |
| CCND1 | Chr11 (69,641,156–69,654,474) | Nucleus and cytosol | 3.2 | 1.5 |
| ADAM9 | Chr8 (38,996,767–39,105,261) | Extracellular space | 8.0 | 0.3 |
| SOX4 | Chr6 (21,593,751–21,598,619) | Nucleus | 1.8 | 1.2 |
| TGFB | Chr19 (41,330,323–41,353,922) | Extracellular space, cytosol, and nucleus | 1.8 | 0.9 |
| MAPK1 | Chr22 (21,759,657–21,867,680) | Extracellular space, cytosol, and nucleus | 1.9 | 0.9 |
| Type of RNA | rs * | |
|---|---|---|
| miRNA | lncRNA | |
| miR-148a | MAFG-DT | −0.37 |
| miR-203a | MALAT1 | −0.42 |
| OIP5-AS1 | −0.42 | |
| MLK7-AS1 | −0.41 | |
| LINC00339 | −0.39 | |
| miRNA | mRNA | |
| miR-148a | BCL2 | −0.44 |
| miR-203a | CDK4 | −0.54 |
| c-MET | −0.42 | |
| ZEB1 | −0.34 | |
| ZEB2 | −0.41 | |
| WNT4 | −0.41 | |
| YAP1 | −0.53 | |
| lncRNA | mRNA | |
| MAFG-DT | BCL2 | 0.45 |
| MALAT1 | CDK4 | 0.52 |
| c-MET | 0.39 | |
| ZEB1 | 0.38 | |
| OIP5-AS1 | CDK4 | 0.53 |
| c-MET | 0.37 | |
| WNT4 | 0.56 | |
| YAP1 | 0.61 | |
| ZEB1 | 0.57 | |
| ZEB2 | 0.65 | |
| MLK7-AS1 | CDK4 | 0.40 |
| YAP1 | 0.51 | |
| ZEB2 | 0.49 | |
| LINC00339 | CDK4 | 0.53 |
| YAP1 | 0.55 | |
| ZEB1 | 0.43 | |
| RNA | Sequence (GRCh37/hg19) | Binding Site |
|---|---|---|
| lncRNAs | ||
| MALAT1 | no binding site | - |
| OIP5-AS1 | chr15: 41,593,028–41,593,127 | 7-mer-m8 |
| MLK7-AS1 | chr2: 174,080,356–174,080,455 | 7-mer-m8 |
| LINC00339 | no binding site | - |
| mRNAs | ||
| CDK4 | no binding site | - |
| c-MET | chr7: 116,415,099–116,415,198 | 8-mer |
| WNT4 | no binding site | - |
| YAP1 | no binding site | - |
| ZEB1 | chr10: 31,817,289–31,817,388 | 7-mer-m8 |
| ZEB2 | chr2: 145,184,392–145,184,491 | 8-mer |
| Clinical Characteristics | N (%) | |
|---|---|---|
| Age, years | <40 | 7 (15) |
| 40–60 | 22 (48) | |
| >60 | 17 (37) | |
| Histological type | Borderline serous adenocarcinoma | 3 (6) |
| Serous adenocarcinoma | 33 (72) | |
| Endometrioid adenocarcinoma | 5 (12) | |
| Clear cell adenocarcinoma | 1 (2) | |
| Mixed epithelial tumors | 2 (4) | |
| Undifferentiated carcinoma | 2 (4) | |
| Stage | I | 12 (26) |
| II | 10 (22) | |
| III | 22 (48) | |
| IV | 2 (4) | |
| Primary tumor site and size | T1 | 12 (26) |
| T2 | 10 (22) | |
| T3 | 24 (52) | |
| Lymph node | Nx | 16 (34) |
| N0 | 30 (65) | |
| N1 | 10 (21) | |
| Peritoneal metastases | Absent | 32 (69) |
| Present | 14 (31) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pronina, I.V.; Uroshlev, L.A.; Moskovtsev, A.A.; Zaichenko, D.M.; Filippova, E.A.; Fridman, M.V.; Burdennyy, A.M.; Loginov, V.I.; Kazubskaya, T.P.; Kushlinskii, N.E.; et al. Dysregulation of lncRNA–miRNA–mRNA Interactome as a Marker of Metastatic Process in Ovarian Cancer. Biomedicines 2022, 10, 824. https://doi.org/10.3390/biomedicines10040824
Pronina IV, Uroshlev LA, Moskovtsev AA, Zaichenko DM, Filippova EA, Fridman MV, Burdennyy AM, Loginov VI, Kazubskaya TP, Kushlinskii NE, et al. Dysregulation of lncRNA–miRNA–mRNA Interactome as a Marker of Metastatic Process in Ovarian Cancer. Biomedicines. 2022; 10(4):824. https://doi.org/10.3390/biomedicines10040824
Chicago/Turabian StylePronina, Irina V., Leonid A. Uroshlev, Alexey A. Moskovtsev, Danila M. Zaichenko, Elena A. Filippova, Marina V. Fridman, Alexey M. Burdennyy, Vitaly I. Loginov, Tatiana P. Kazubskaya, Nikolay E. Kushlinskii, and et al. 2022. "Dysregulation of lncRNA–miRNA–mRNA Interactome as a Marker of Metastatic Process in Ovarian Cancer" Biomedicines 10, no. 4: 824. https://doi.org/10.3390/biomedicines10040824
APA StylePronina, I. V., Uroshlev, L. A., Moskovtsev, A. A., Zaichenko, D. M., Filippova, E. A., Fridman, M. V., Burdennyy, A. M., Loginov, V. I., Kazubskaya, T. P., Kushlinskii, N. E., Dmitriev, A. A., Braga, E. A., & Brovkina, O. I. (2022). Dysregulation of lncRNA–miRNA–mRNA Interactome as a Marker of Metastatic Process in Ovarian Cancer. Biomedicines, 10(4), 824. https://doi.org/10.3390/biomedicines10040824