
Elucidation of the Potential Hair Growth-Promoting Effect of Botryococcus terribilis, Its Novel Compound Methylated-Meijicoccene, and C32 Botryococcene on Cultured Hair Follicle Dermal Papilla Cells Using DNA Microarray Gene Expression Analysis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Cells and Cell Culture
2.3. RNA Extraction
2.4. DNA Microarray Analysis
2.5. TaqMan Quantitative RT-PCR Gene Expression Analysis
2.6. Soxhlet Extraction
2.7. Cell Viability Assay
2.8. Statistical Analysis
3. Results
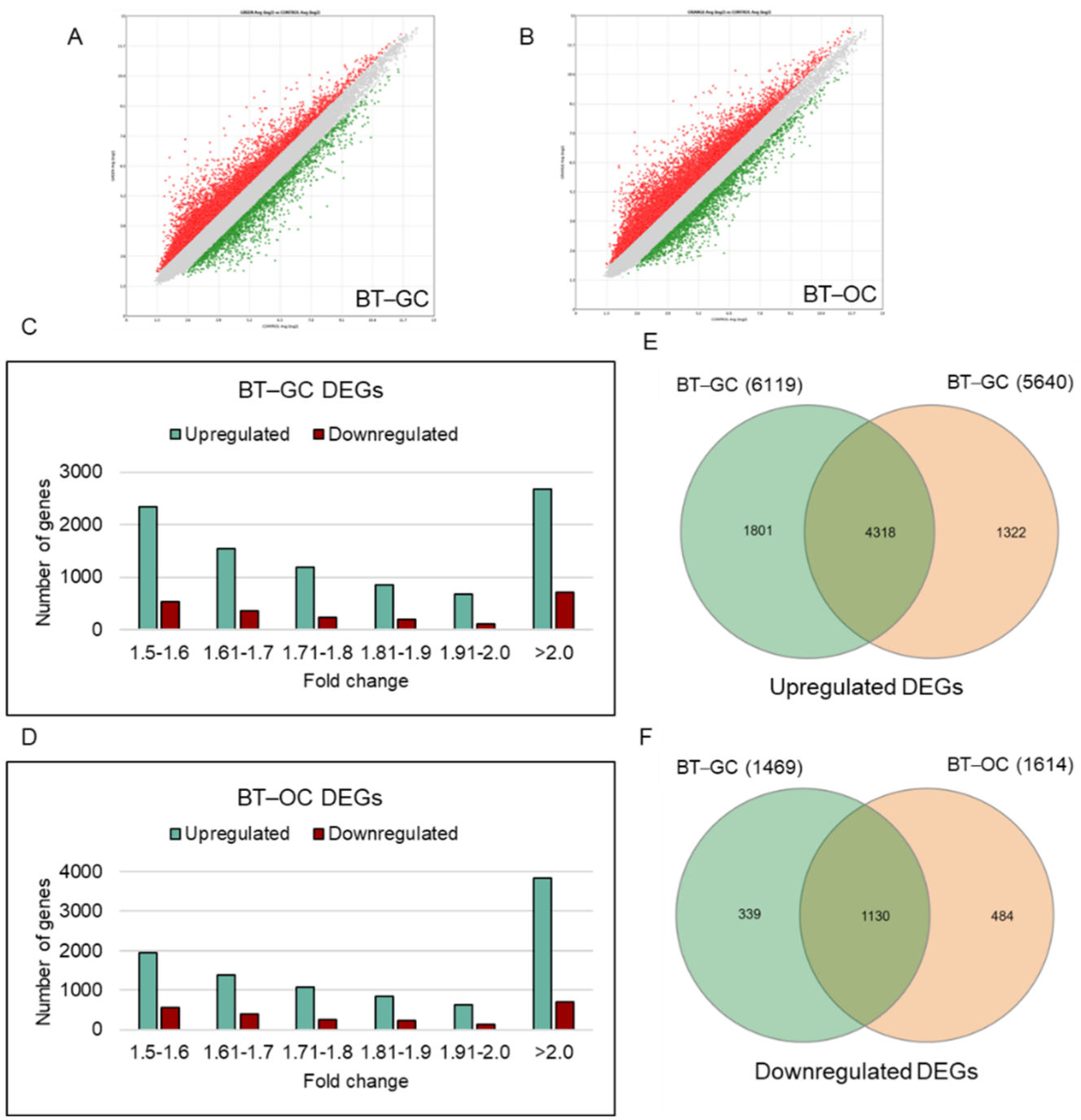
3.1. Comparison of Significantly Enriched DEGs of BT-GC and BT-OC
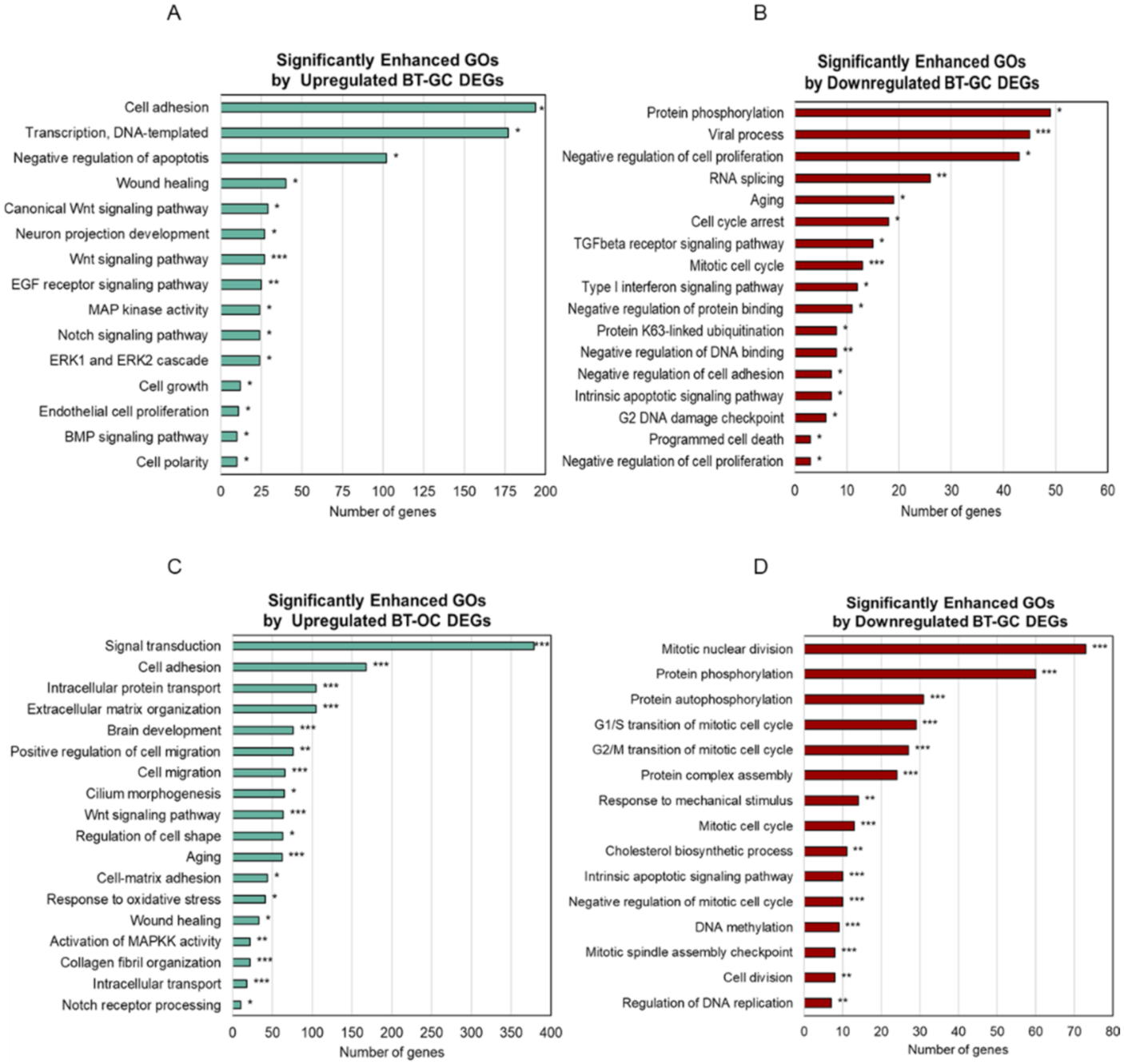
3.2. Significantly Enriched Biological Process and Cellular Components of BT-GC and BT-OC
3.3. Comparison of Significantly Enriched Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathways by BT-GC and BT-OC
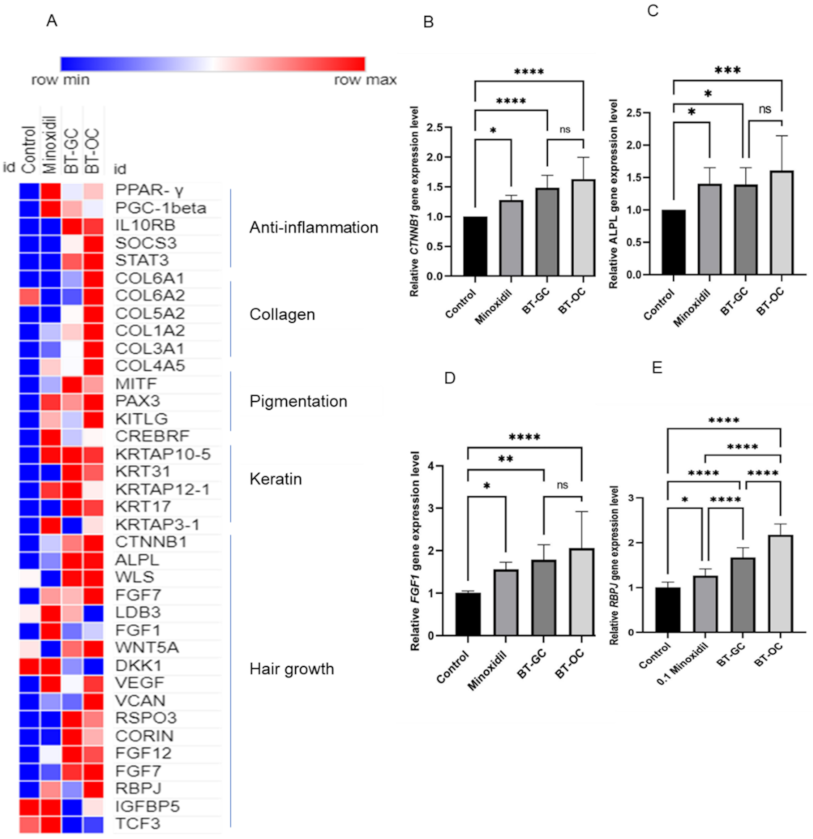
3.4. BT-GC and BT-OC Treatment Regulated Essential Functional Categories and Gene Expression Related to Hair Growth
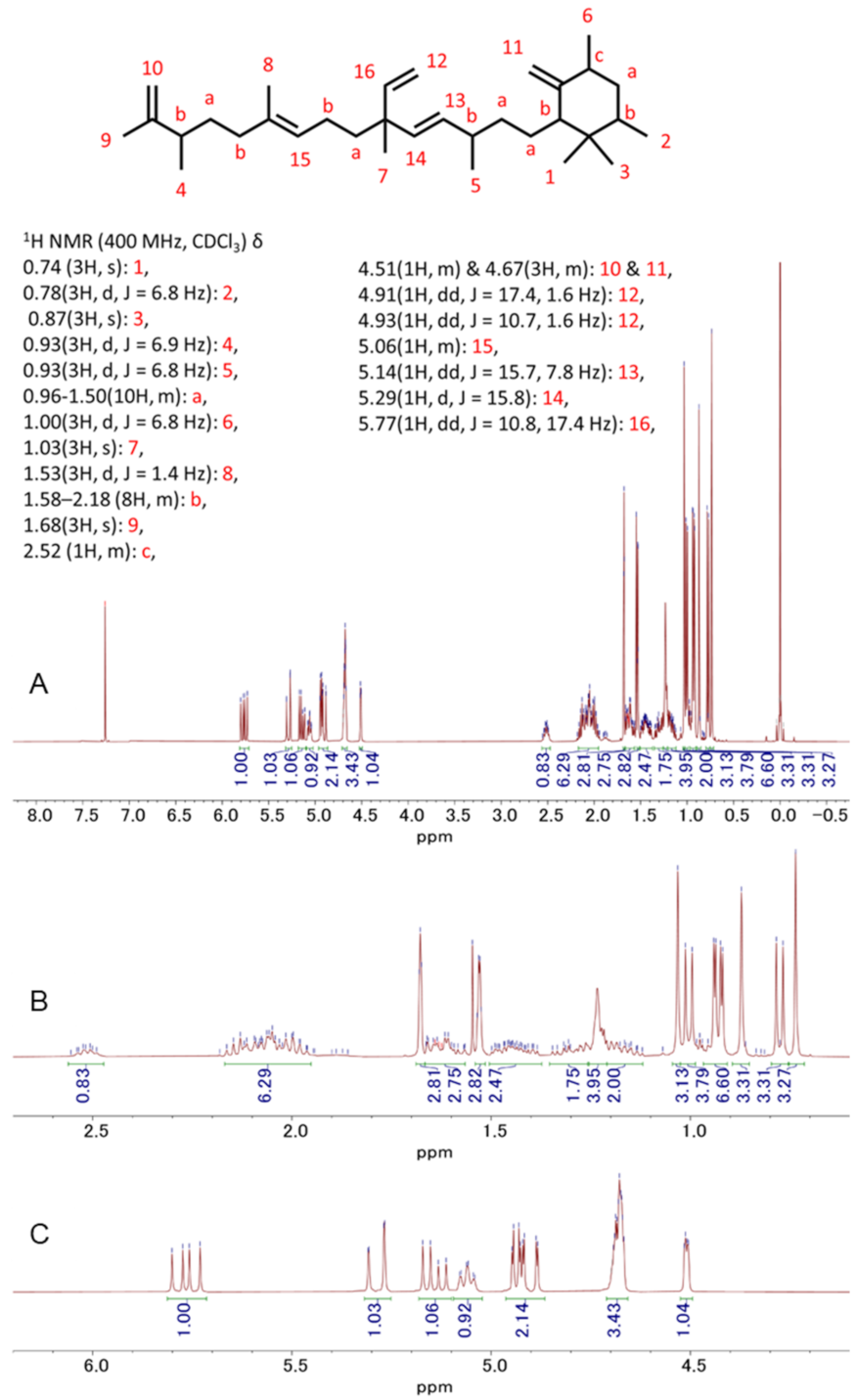
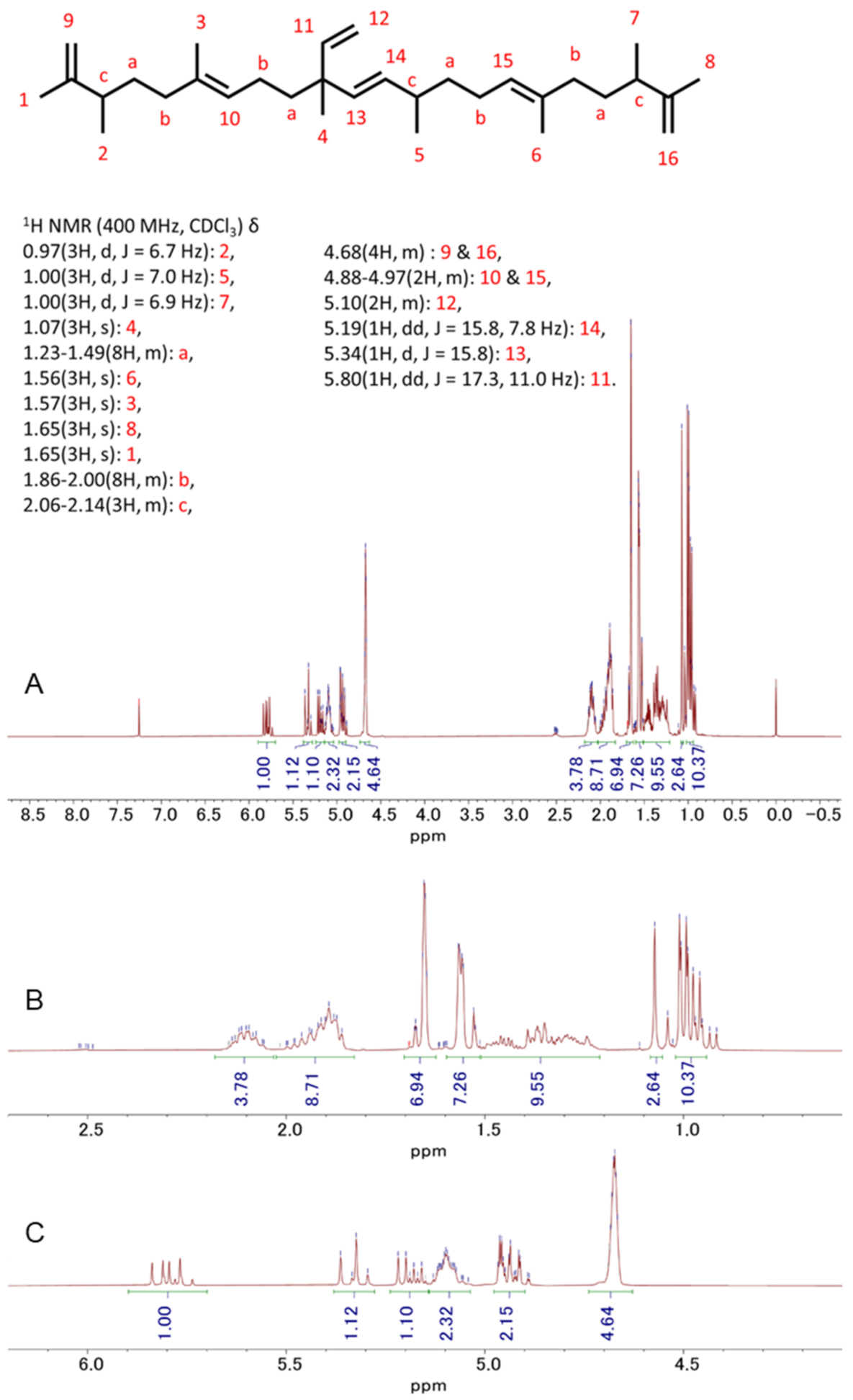
3.5. Detection and Isolation of Two Major Compounds of Botryococcus terribilis—Me-Meijicoccene and C32 Botryococcene
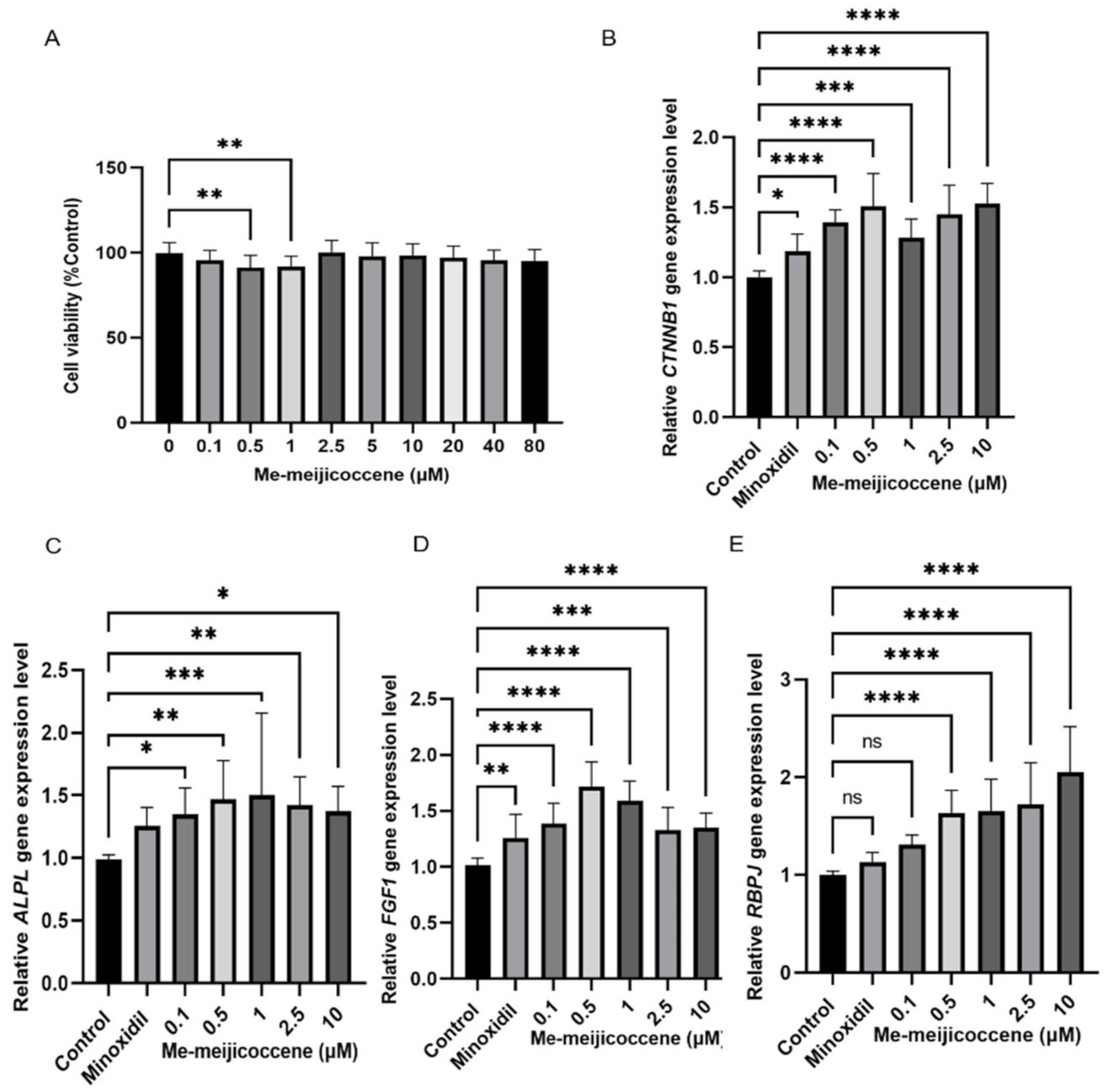
3.6. Me-Meijicoccene and C32 Botryococcene Treatment Regulated the Gene Expression Levels of Hair Growth Markers
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
References
- Semalty, M.; Semalty, A.; Joshi, G.P.; Rawat, M.S.M. Hair growth and rejuvenation: An overview. J. Dermatol. Treat. 2011, 22, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Bejaoui, M.; Villareal, M.O.; Isoda, H. β-catenin-mediated hair growth induction effect of 3,4,5-tri-O-caffeoylquinic acid. Aging 2019, 11, 4216–4237. [Google Scholar] [CrossRef] [PubMed]
- Orasan, M.S.; Roman, I.I.; Coneac, A.; Muresan, A.; Orasan, R.I. Hair loss and regeneration performed on animal models. Clujul Med. 2016, 89, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.R.; Schmidt-Ullrich, R. The hair follicle as a dynamic miniorgan. Curr. Biol. 2009, 19, R132–R142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linayage, D.; Sinclair, R. Telogen Effluvium. Cosmetics 2016, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Enshell-Seijffers, D.; Lindon, C.; Kashiwagi, M.; Morgan, B.A. Article b-catenin activity in the dermal papilla regulates morphogenesis and regeneration of hair. Dev. Cell 2010, 18, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Truong, V.L.; Bak, M.J.; Lee, C.; Jun, M.; Jeong, W.S. Hair regenerative mechanisms of red ginseng oil and its major components in the testosterone-induced delay of anagen entry in C57BL/6 mice. Molecules 2017, 22, 1505. [Google Scholar] [CrossRef] [Green Version]
- Driskell, R.R.; Clavel, C.; Rendl, M.; Watt, F.M. Hair follicle dermal papilla cells at a glance. J. Cell Sci. 2011, 124, 1179–1182. [Google Scholar] [CrossRef] [Green Version]
- Panteleyev, A.A.; Botchkareva, N.V.; Sundberg, J.P.; Christiano, A.M.; Paus, R. The role of the hairless (hr) gene in the regulation of hair follicle catagen transformation. Am. J. Pathol. 1999, 155, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Rishikaysh, P.; Dev, K.; Diaz, D.; Qureshi, W.M.; Filip, S.; Mokry, J. Signaling involved in hair follicle morphogenesis and development. Int. J. Mol. Sci. 2014, 15, 1647–1670. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Caporgno, M.P.; Mathys, A. Trends in microalgae incorporation into innovative food products with potential health benefits. Front. Nutr. 2018, 5, 58. [Google Scholar] [CrossRef]
- Kothari, R.; Pandey, A.; Ahmad, S.; Kumar, A.; Pathak, V.V.; Tyagi, V.V. Microalgal cultivation for value-added products: A critical enviro-economical assessment. 3 Biotech 2017, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Ito, N.; Uno, Y.; Nishii, I.; Kagiwada, S.; Okada, S.; Noguchi, T. Transformation of lipid bodies related to hydrocarbon accumulation in a green alga, Botryococcus braunii (Race B). PLoS ONE 2013, 8, e81626. [Google Scholar] [CrossRef]
- Metzger, P.; Templier, J.; Casadevall, E.; Couté, A. A n-alkatriene and some n-alkadienes from the A race of the green alga Botryococcus braunii. Phytochemistry 1986, 25, 1869–1872. [Google Scholar] [CrossRef]
- Metzger, P.; Allard, B.; Casadevall, E.; Berkaloff, C.; Couté, A. Structure and chemistry of a new chemical race of botryococcus braunii (chlorophyceae) that produces lycopadiene, a tetraterpenoid hydrocarbon. J. Phycol. 1990, 2, 258–266. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.R.; Deepika, G.; Ravishankar, G.A.; Sarada, R.; Panduranga, N.B.; Yuepeng, S.; Bo, L. Botryococcus as an alternative source of carotenoids and its possible applications—An overview. Crit. Rev. Biotechnol. 2017, 38, 541–558. [Google Scholar]
- Sasaki, K.; Othman, M.B.; Demura, M.; Watanabe, M.; Isoda, H. Modulation of neurogenesis through the promotion of energy production activity is behind the antidepressant-like effect of colonial green alga, Botryococcus braunii. Front. Physiol. 2017, 8, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.R.; Sarada, R.; Baskaran, V.; Ravishankar, G.A. Antioxidant activity of Botryococcus braunii extract elucidated in vitro models. J. Agric. Food Chem. 2006, 54, 4593–4599. [Google Scholar] [CrossRef]
- Buono, S.; Langellotti, A.L.; Martello, A.; Bimonte, M.; Tito, A.; Carola, A.; Apone, F.; Colucci, G.; Fogliano, V. Biological activities of dermatological interest by the water extract of the microalga Botryococcus braunii. Arch. Dermatol. Res. 2012, 304, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Tanoi, T.; Kawachi, M.; Watanabe, M. Effects of Carbon source on growth and morphology of Botryococcus braunii. J. Appl. Phycol. 2010, 23, 25–33. [Google Scholar] [CrossRef]
- Gopalasatheeskumar, K. Significant role of soxhlet extraction process in phytochemical research. Mintage J. Pharm. Med. Sci. 2019, 7, 43–47. [Google Scholar]
- Murakami, M.; Nakano, H.; Yamaguchi, K.; Konosu, S.; Nakayama, O.; Matsumoto, Y.; Iwamoto, H. Meijicoccene, a new cyclic hydrocarbon from Botryococcus braunii. Phytochemistry 1988, 27, 455–457. [Google Scholar] [CrossRef]
- Cardozo, K.H.M.; Guaratini, T.; Barros, M.P.; Falcão, V.R.; Tonon, A.P.; Lopes, N.P.; Campos, S.; Torres, M.A.; Souza, A.O.; Colepicolo, P.; et al. Metabolites from algae with economical impact. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2007, 146, 60–78. [Google Scholar] [CrossRef]
- Guihéneuf, F.; Khan, A.; Tran, L.S.P. Genetic engineering: A promising tool to engender physiological, biochemical, and molecular stress resilience in green microalgae. Front. Plant Sci. 2016, 7, 400. [Google Scholar] [CrossRef] [Green Version]
- Khora, S.S.; Navya, P. Bioactive polysaccharides from marine macroalgae. In Encyclopedia of Marine Biotechnology; Kim, S.K., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; Volume 1, pp. 121–145. [Google Scholar]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity screening of microalgae for antioxidant, anti-Inflammatory, anticancer, anti-Diabetes, and antibacterial activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Ji, K.; Kim, Y.; Kim, Y.T. A Study on the tyrosinase inhibitory and antioxidant effect of microalgae extracts. Microbiol. Biotechnol. Lett. 2021, 49, 167–173. [Google Scholar] [CrossRef]
- Abreu, I.N.; Aksmann, A.; Bajhaiya, A.K.; Benlloch, R.; Giordano, M.; Pokora, W.; Selstam, E.; Moritz, T. Changes in lipid and carotenoid metabolism in Chlamydomonas reinhardtii during induction of CO2-concentrating mechanism: Cellular response to low CO2 stress. Algal Res. 2020, 2020. 52, 102099. [Google Scholar] [CrossRef]
- Hrckulak, D.; Kolar, M.; Strnad, H.; Korinek, V. TCF/LEF Transcription Factors: An update from the internet resources. Cancers 2016, 8, 70. [Google Scholar] [CrossRef] [Green Version]
- Cadigan, K.M.; Liu, Y.I. Wnt signaling: Complexity at the surface. J. Cell Sci. 2006, 119, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bejsovec, A. Wnt signaling: An embarrassment of receptors. Curr. Biol. 2000, 10, R919–R922. [Google Scholar] [CrossRef] [Green Version]
- Wodarz, A.; Nusse, R. Mechanisms of Wnt signaling in development. Annu. Rev. Cell Dev. Biol. 1998, 14, 59–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niida, A.; Hiroko, T.; Kasai, M.; Furukawa, Y.; Nakamura, Y.; Suzuki, Y.; Sugano, S.; Akiyama, T. DKK1, a negative regulator of Wnt signaling, is a target of the β-catenin/TCF pathway. Oncogene 2004, 23, 8520–8526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubin-Houzelstein, G. Notch signaling and the developing hair follicle. In Madame Curie Bioscience Database; Reichrath, J., Reichrath, S., Eds.; Landes Biosciences: Austin, TX, USA, 2010; pp. 2000–2013. [Google Scholar]
- Yamamoto, N.; Tanigaki, K.; Han, H.; Hiai, H.; Honjo, T. Notch/RBP-J signaling regulates epidermis/hair fate determination of hair follicular stem cells. Curr. Biol. 2003, 13, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Zhang, X.; Chen, Z.; Li, Y.; Li, B.; Li, L. ERK1/2 Pathway Is involved in the enhancement of fatty acids from Phaeodactylum tricornutum Extract (PTE) on hair follicle cell proliferation. BioMed Res. Int. 2020, 11, 2916104. [Google Scholar] [CrossRef]
- Huang, H.C.; Lin, H.; Huang, M.C. Lactoferrin promotes hair growth in mice and increases dermal papilla cell proliferation through Erk/Akt and Wnt signaling pathways. Arch. Dermatol. Res. 2019, 311, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, W.Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence, and apoptosis. J. Recept. Signal Transduct. 2015, 35, 600–604. [Google Scholar] [CrossRef]
- Lin, W.H.; Xiang, L.J.; Shi, H.X.; Zhang, J.; Jiang, L.P.; Cai, P.T.; Lin, Z.L.; Lin, B.B.; Huang, Y.; Zhang, H.L.; et al. Fibroblast growth factors stimulate hair growth through β-catenin and Shh expression in C57BL/6 mice. BioMed Res. Int. 2015, 2015, 730139. [Google Scholar]
- Imamura, T. Physiological functions and underlying mechanisms of fibroblast growth factor (FGF) family members: Recent findings and implications for their pharmacological application. Biol. Pharm. Bull. 2014, 37, 1081–1089. [Google Scholar] [CrossRef] [Green Version]
- Lv, X.; Chen, W.; Sun, W.; Hussain, Z.; Wang, S.; Wang, J. Analysis of lncRNAs expression profiles in hair follicle of hu sheep lambskin. Animals 2020, 10, 1035. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Yoon, J.; Shin, S.H.; Zahoor, M.; Kim, H.J.; Park, P.J.; Park, W.S.; Min, D.; Kim, H.Y.; Choi, K.Y. Valproic acid induces hair regeneration in murine model and activates alkaline phosphatase activity in human dermal papilla cells. PLoS ONE 2012, 7, e34152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iida, M.; Ihara, S.; Matsuzaki, T. Hair cycle-dependent changes of alkaline phosphatase activity in the mesenchyme and epithelium in mouse vibrissa follicles. Dev. Growth Differ. 2007, 49, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fan, Z.; Wang, X.; Mo, M.; Zeng, S.B.; Xu, R.H.; Wang, X.; Wu, Y. PI3K/Akt signaling pathway is essential for de novo hair follicle regeneration. Stem Cell Res. Ther. 2020, 11, 144. [Google Scholar] [CrossRef] [Green Version]
- Laurikkala, J.; Pispa, J.; Jung, H.S.; Nieminen, P.; Mikkola, M.; Wang, X.; Saarialho-Kere, U.; Galceran, J.; Grosschedl, R.; Thesleff, I. Regulation of hair follicle development by the TNF signal ectodysplasin and its receptor Edar. Development 2002, 129, 2541–2553. [Google Scholar] [CrossRef]
- Drosten, M.; Lechuga, C.; Barbacid, M. Ras signaling is essential for skin development. Oncogene 2014, 33, 2857–2865. [Google Scholar] [CrossRef]
- Geyfman, M.; Andersen, B. Clock genes, hair growth and aging. Aging 2010, 2, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Trüeb, R.M. Molecular mechanisms of androgenetic alopecia. Exp. Gerontol. 2002, 37, 981–990. [Google Scholar] [CrossRef]
- Qiu, W.; Chuong, C.M.; Lei, M. Regulation of melanocyte stem cells in the pigmentation of skin and its appendages: Biological patterning and therapeutic potentials. Exp. Dermatol. 2019, 28, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Roh, C.; Tao, Q.; Lyle, S. Dermal papilla-induced hair differentiation of adult epithelial stem cells from human skin. Physiol. Genom. 2004, 19, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Young, T.H.; Tu, H.R.; Chan, C.C.; Huang, Y.C.; Yen, M.H.; Cheng, N.C.; Chiu, H.C.; Lin, S.J. The enhancement of dermal papilla cell aggregation by extracellular matrix proteins through effects on cell-substratum adhesivity and cell motility. Biomaterials 2009, 30, 5031–5040. [Google Scholar] [CrossRef] [PubMed]
- Suchonwanit, P.; Thammarucha, S.; Leerunyakul, K. Minoxidil and its use in hair disorders: A review. Drug Des. Dev. Ther. 2019, 13, 2777–2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, D.J., Jr.; Hodges, S. Medical therapy for benign prostatic hyperplasia: Sexual dysfunction and impact on quality of life. Int. J. Impot. Res. 2003, 15, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondaini, N.; Gontero, P.; Giubilei, G.; Lombardi, G.; Cai, T.; Gavazzi, A.; Bartoletti, R. Finasteride 5mg and sexual side effects: How many of these are related to a nocebo phenomenon? J. Sex. Med. 2007, 4, 1708–1712. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Biological Function | Fold-Change | * p-Value |
|---|---|---|---|---|
| CORIN | Corin, Serine Peptidase | Morphogenesis of HF | 33.29 | 4.37 × 10−20 |
| SOX2 | SRY (sex determining region Y) box 2 | Controls mesenchymal–epithelial crosstalk | 5.31 | 1.72 × 10−17 |
| STAT5A | Signal transducer and activator of transcription 5 | Anagen inducer | 4.2 | 1.25 × 10−14 |
| WLS | Wntless | Wnt secretion and pathway | 3.83 | 1.19 × 10−13 |
| FGF7 | Fibroblast growth factor 7 | HF morphogenesis; hair growth cycle regulation | 3.81 | 1.09 × 10−16 |
| CTNNB1 | Catenin (cadherin associated protein) beta 1 | Regulation of anagen phase regulation, fibroblast growth, cell proliferation, and HF morphogenesis | 3.59 | 1.09 × 10−14 |
| MITF | Microphthalmia-associated transcription factor | Controls pigmentation enzymes | 2.89 | 5.36 × 10−12 |
| ALPL | Alkaline phosphatase | Wnt/β-catenin pathway regulator | 2.39 | 6.93 × 10−13 |
| WNT5A | Wingless-type MMTV, member 11 | Wnt pathway | 2.36 | 5.19 × 10−14 |
| COL3A1 | Collagen, type III, alpha 1 | Fibrillogenesis in the skin and the HF | 2.36 | 1.85 × 10−14 |
| IGFBP2 | Insulin-like growth factor Binding Protein 2 | IGF1 receptor | 2.17 | 1.31 × 10−13 |
| VCAN | Versican | Cell aggregation, adhesion, sand proliferation | 2.1 | 3.03 × 10−13 |
| SOX4 | SRY (sex determining region Y)-box 4 | Hair regeneration | 2.08 | 4.08 × 10−12 |
| BMP8A | Bone morphogenetic protein 8a | Control HF growth and development | 1.98 | 6.30 × 10−12 |
| STAT3 | Signal transducer and activator of transcription 3 | Wound healing and HF development | 1.9 | 4.92 × 10−12 |
| FGF22 | Fibroblast growth factor 22 | Cutaneous development and repair | 1.82 | 4.59 × 10−11 |
| RBPJ | Recombinant signal binding protein J | Transcriptional effector of Notch pathway | 1.74 | 3.51 × 10−09 |
| IGF1 | Insulin-like growth factor 1 | Maintains anagen phase and cell proliferation | 1.7 | 1.09 × 10−08 |
| LEF1 | Lymphoid enhancer factor 1 | Regulation of DPCs proliferation by Wnt signaling | 1.58 | 1.44 × 10−09 |
| HGF | Hepatocyte growth factor | Promotes follicular growth | 1.57 | 1.38 × 10−08 |
| FGF1 | Fibroblast growth factor 1 | Anagen inducer | 1.53 | 7.67 × 10−07 |
| FGFR1 | Fibroblast growth factor receptor 1 | Primary transducers of FGF signaling in DP | 1.53 | 1.82 × 10−08 |
| Gene Symbol | Gene Name | Biological Function | Fold-Change | * p-Value |
|---|---|---|---|---|
| VCAN | Versican | Regulates Cell aggregation, adhesion, and proliferation | 5.46 | 5.66 × 10−18 |
| CORIN | Corin, Serine Peptidase | Morphogenesis of HF | 22.08 | 1.84 × 10−19 |
| CTNNB1 | Catenin (cadherin associated protein), beta 1 | Regulation of anagen phase regulation, fibroblast growth, cell proliferation, and HF morphogenesis | 4.41 | 2.04 × 10−15 |
| FGF7 | Fibroblast growth factor 7 | HF morphogenesis; hair growth cycle regulation | 4.09 | 6.16 × 10−17 |
| WLS | Wntless | Wnt secretion and pathway | 3.89 | 1.06 × 10−15 |
| COL3A1 | Collagen, type III, alpha 1 | Fibrillogenesis in the skin and the HF | 3.75 | 1.43 × 10−16 |
| RBPJ | Recombinant signal binding protein J | Transcriptional effector of Notch pathway | 3.69 | 1.20 × 10−13 |
| WNT5A | Wingless-type MMTV, member 11 | Wnt pathway | 3.14 | 1.39 × 10−16 |
| SOX2 | SRY (sex determining region Y) box 2 | Controls mesenchymal-epithelial crosstalk | 2.95 | 2.74 × 10−15 |
| STAT5A | Signal transducer and activator of transcription 5 | Anagen inducer | 2.81 | 4.48 × 10−13 |
| IGF1R | Insulin-like growth factor 1 | Maintains anagen phase and cell proliferation | 2.51 | 1.41 × 10−13 |
| IGFBP2 | Insulin-like growth factor Binding Protein 2 | IGF1 receptor | 2.44 | 2.73 × 10−14 |
| ALPL | Alkaline phosphatase | Wnt/β-catenin pathway regulator | 2.37 | 7.61 × 10−13 |
| MITF | Microphthalmia-associated transcription factor | Controls pigmentation enzymes | 2.31 | 5.36 × 10−11 |
| STAT3 | Signal transducer and activator of transcription 3 | Wound healing and HF development | 2.09 | 1.01 × 10−12 |
| LEF1 | Lymphoid enhancer factor 1 | Regulation of DPCs proliferation by Wnt signaling | 2.06 | 8.88 × 10−12 |
| SOX4 | SRY (sex determining region Y)-box 4 | Hair regeneration | 1.97 | 1.86 × 10−13 |
| CDH3 | Cadherin 3, Type 1, P-cadherin (placenta) | Cell–cell communication and adhesion | 1.95 | 2.73 × 10−10 |
| FGF1 | Fibroblast growth factor 1 | Anagen inducer | 1.94 | 6.17 × 10−11 |
| BMP8A | Bone morphogenetic protein 8a | Control HF growth and development | 1.76 | 4.94 × 10−11 |
| FGF22 | Fibroblast growth factor 22 | Cutaneous development and repair | 1.63 | 5.04 × 10−10 |
| HGF | Hepatocyte growth factor | Promotes follicular growth | 1.58 | 2.98 × 10−08 |
| FGFR1 | Fibroblast growth factor receptor 1 | Primary transducers of FGF signaling in DP | 1.52 | 2.16 × 10−08 |
| Gene Symbol | Gene Name | Biological Function | Fold-Change | * p-Value |
|---|---|---|---|---|
| IGFBP5 | Insulin like growth factor binding protein 5 | Negative regulator of cell proliferation in the HF | −13.19 | 1.43 × 10−18 |
| BDNF | Brain-derived Neurotrophic Factor | Catagen induction and inhibition of hair shaft elongation | −3.1 | 2.59 × 10−15 |
| TGFB1 | Transforming growth factor beta 1 | Catagen induction marker | −2.61 | 2.11 × 10−13 |
| ZYX | Zyxin | Inhibits HF growth by regulating fibroblast HF cycle and promotes cell apoptosis, thus reduced expression enhances hair shaft, delays catagen entry, and HFDPCs proliferation and inductivity | −2.42 | 1.78 × 10−12 |
| DKK1 | Dickkopf Wnt signaling pathway inhibitor 3 | Wnt signaling negative regulator | −2.35 | 4.81 × 10−12 |
| BMP4 | Bone morphogenetic protein 4 | Suppress proliferation in the hair matrix | −2.31 | 3.11 × 10−10 |
| GSK3B | Glycogen synthase kinase 3 beta | Phosphorylation of β-catenin | −1.64 | 1.74 × 10−08 |
| AEBP1 | AE binding protein 1 | Regulator of telogen HF | −1.59 | 4.84 × 10−11 |
| H2AFV | H2A histone family | Stem cell quiescence, Anagen delay | −1.53 | 2.06 × 10−09 |
| Gene Symbol | Gene Name | Biological Function | Fold-Change | * p-Value |
|---|---|---|---|---|
| Igfbp5 | Insulin-like growth factor binding protein 5 | Negative regulator of cell proliferation in the hair follicle | −5.29 | 2.63 × 10−13 |
| DKK1 | Dickkopf WNT signaling pathway inhibitor 3 | Wnt signaling negative regulator | −3.67 | 2.33 × 10−14 |
| TGFB1 | Transforming growth factor beta 1 | Catagen induction marker | −2.94 | 5.27 × 10−14 |
| ZYX | Zyxin | Inhibits HF growth by regulating fibroblast HF cycle and promotes cell apoptosis | −2.37 | 5.64 × 10−12 |
| BDNF | Brain-Derived Neurotrophic Factor | Catagen induction and inhibition of hair shaft elongation | −1.94 | 1.26 × 10−13 |
| GSK3B | Glycogen synthase kinase 3 beta | Phosphorylation of β-catenin | −1.71 | 3.19 × 10−09 |
| LDB3 | LIM Domain Binding 3 | Wnt-responsive gene; Regulation of hair follicle during telogen | −1.53 | 5.35 × 10−06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliva, A.K.; Bejaoui, M.; Hirano, A.; Arimura, T.; Linh, T.N.; Uchiage, E.; Nukaga, S.; Tominaga, K.; Nozaki, H.; Isoda, H. Elucidation of the Potential Hair Growth-Promoting Effect of Botryococcus terribilis, Its Novel Compound Methylated-Meijicoccene, and C32 Botryococcene on Cultured Hair Follicle Dermal Papilla Cells Using DNA Microarray Gene Expression Analysis. Biomedicines 2022, 10, 1186. https://doi.org/10.3390/biomedicines10051186
Oliva AK, Bejaoui M, Hirano A, Arimura T, Linh TN, Uchiage E, Nukaga S, Tominaga K, Nozaki H, Isoda H. Elucidation of the Potential Hair Growth-Promoting Effect of Botryococcus terribilis, Its Novel Compound Methylated-Meijicoccene, and C32 Botryococcene on Cultured Hair Follicle Dermal Papilla Cells Using DNA Microarray Gene Expression Analysis. Biomedicines. 2022; 10(5):1186. https://doi.org/10.3390/biomedicines10051186
Chicago/Turabian StyleOliva, Aprill Kee, Meriem Bejaoui, Atsushi Hirano, Takashi Arimura, Tran Ngoc Linh, Eriko Uchiage, Sachiko Nukaga, Kenichi Tominaga, Hiroyuki Nozaki, and Hiroko Isoda. 2022. "Elucidation of the Potential Hair Growth-Promoting Effect of Botryococcus terribilis, Its Novel Compound Methylated-Meijicoccene, and C32 Botryococcene on Cultured Hair Follicle Dermal Papilla Cells Using DNA Microarray Gene Expression Analysis" Biomedicines 10, no. 5: 1186. https://doi.org/10.3390/biomedicines10051186
APA StyleOliva, A. K., Bejaoui, M., Hirano, A., Arimura, T., Linh, T. N., Uchiage, E., Nukaga, S., Tominaga, K., Nozaki, H., & Isoda, H. (2022). Elucidation of the Potential Hair Growth-Promoting Effect of Botryococcus terribilis, Its Novel Compound Methylated-Meijicoccene, and C32 Botryococcene on Cultured Hair Follicle Dermal Papilla Cells Using DNA Microarray Gene Expression Analysis. Biomedicines, 10(5), 1186. https://doi.org/10.3390/biomedicines10051186