
Resilience of Spike-Specific Immunity Induced by COVID-19 Vaccines against SARS-CoV-2 Variants

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Sequences of SARS-CoV-2 Variants
2.2. SARS-CoV-2 Spike-Specific B Cell Epitopes Targeted by Neutralizing Antibodies
2.3. SARS-CoV-2 Spike-Specific T Cell Epitopes
2.4. Other Procedures
3. Results
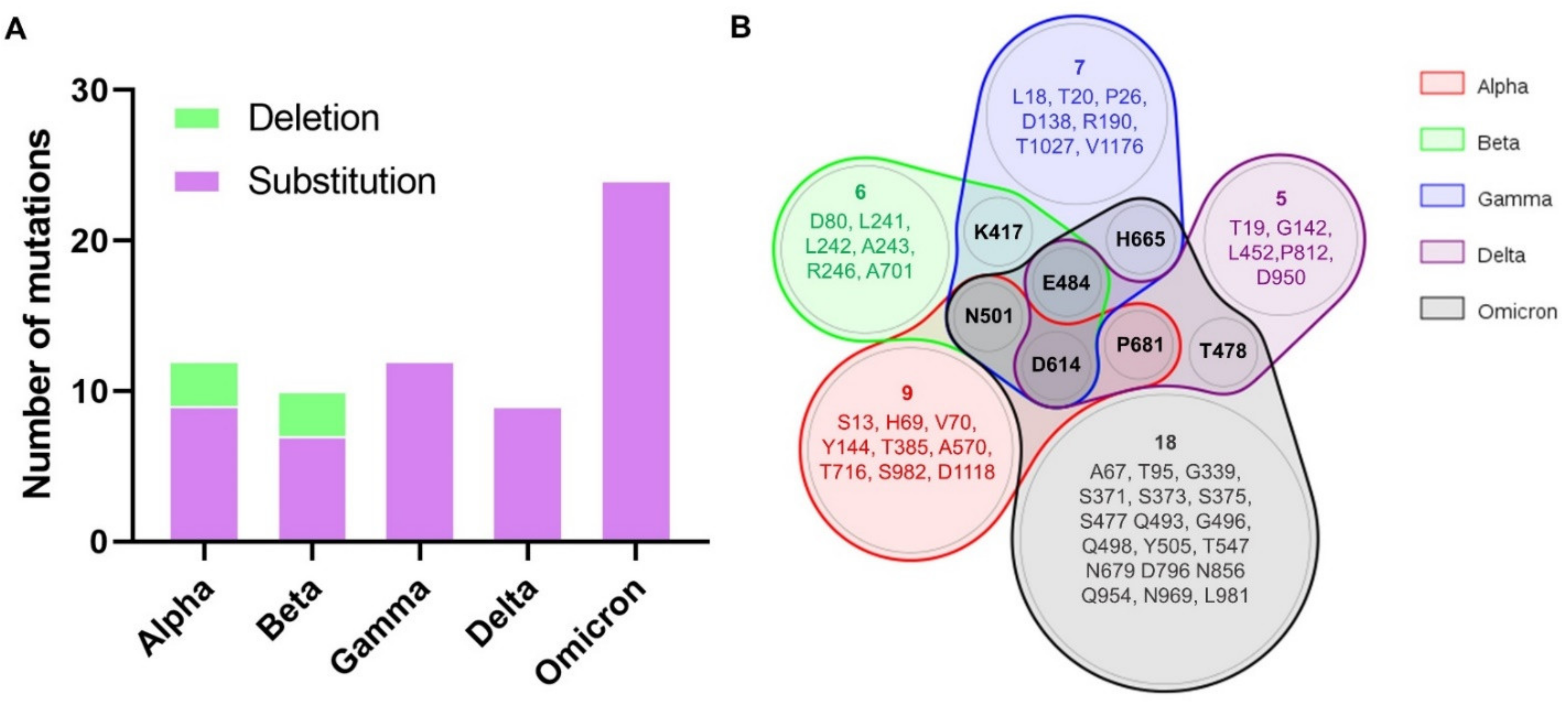
3.1. Identification of Mutations in SARS-CoV-2 Variants
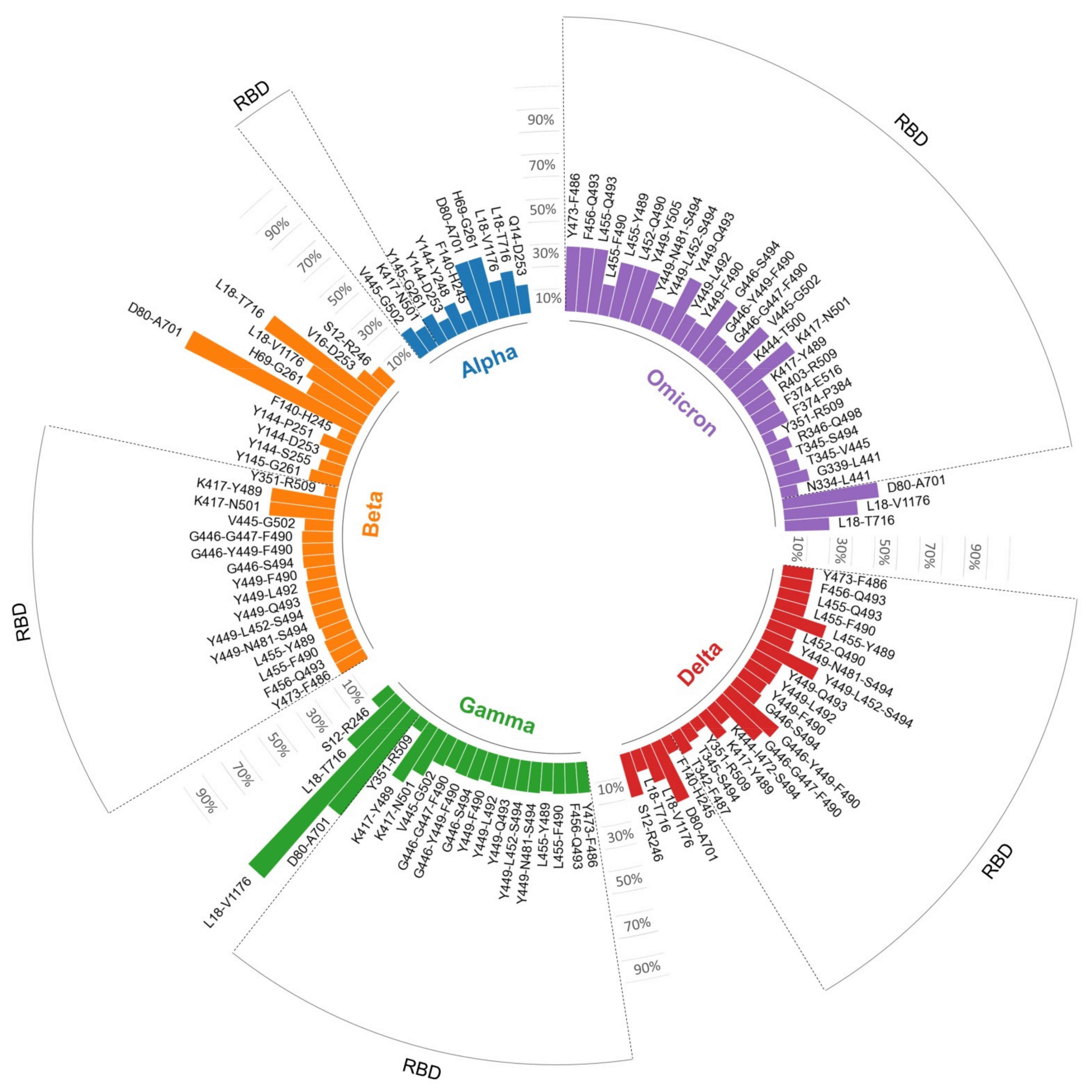
3.2. Mutations in B Cell Epitopes Targeted by Neutralizing Antibodies
3.3. Spike-Specific T Cell Epitopes Mutated in SARS-CoV-2 VOCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marian, A.J. Current state of vaccine development and targeted therapies for COVID-19: Impact of basic science discoveries. Cardiovasc. Pathol. 2021, 50, 107278. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef]
- Shamsi, A.; Mohammad, T.; Anwar, S.; AlAjmi, M.F.; Hussain, A.; Rehman, M.T.; Islam, A.; Hassan, M.I. Glecaprevir and Maraviroc are high-affinity inhibitors of SARS-CoV-2 main protease: Possible implication in COVID-19 therapy. Biosci. Rep. 2020, 26, 40. [Google Scholar] [CrossRef]
- Hwang, Y.C.; Lu, R.M.; Su, S.C.; Chiang, P.Y.; Ko, S.H.; Ke, F.Y.; Liang, K.H.; Hsieh, T.Y.; Wu, H.C. Monoclonal antibodies for COVID-19 therapy and SARS-CoV-2 detection. J. Biomed. Sci. 2022, 29, 1. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Weisblum, Y.; Schmidt, F.; Zhang, F.; DaSilva, J.; Poston, D.; Lorenzi, J.C.C.; Muecksch, F.; Rutkowska, M.; Hoffmann, H.H.; Michailidis, E.; et al. Escape from neutralizing antibodies 1 by SARS-CoV-2 spike protein variants. Elife 2020, 9, e61312. [Google Scholar] [CrossRef]
- Galloway, S.E.; Paul, P.; MacCannell, D.R.; Johansson, M.A.; Brooks, J.T.; MacNeil, A.; Slayton, R.B.; Tong, S.; Silk, B.J.; Armstrong, G.L.; et al. Emergence of SARS-CoV-2 B.1.1.7 Lineage—United States, December 29, 2020–January 12, 2021. MMWR Morb. Mortal Wkly. Rep. 2021, 70, 95–99. [Google Scholar] [CrossRef]
- Choi, J.Y.; Smith, D.M. SARS-CoV-2 variants of concern. Yonsei Med. J. 2021, 62, 961–968. [Google Scholar] [CrossRef]
- Vita, R.; Overton, J.A.; Greenbaum, J.A.; Ponomarenko, J.; Clark, J.D.; Cantrell, J.R.; Wheeler, D.K.; Gabbard, J.L.; Hix, D.; Sette, A.; et al. The immune epitope database (IEDB) 3.0. Nucleic Acids Res. 2015, 43, D405–D412. [Google Scholar] [CrossRef]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B. The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res. 2019, 47, D339–D343. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Silva, J.G.; Araujo-Voces, M.; Quesada, V. NVenn: Generalized, quasi-proportional Venn and Euler diagrams. Bioinform. Oxf. Univ. Press 2018, 34, 2322–2324. [Google Scholar] [CrossRef] [Green Version]
- Molero-Abraham, M.; Lafuente, E.M.; Flower, D.R.; Reche, P.A. Selection of conserved epitopes from hepatitis c virus for pan-populational stimulation of T-cell responses. Clin. Dev. Immunol. 2013, 2013, 601943. [Google Scholar] [CrossRef]
- Nguyen, T.H.O.; Cohen, C.A.; Rowntree, L.C.; Bull, M.B.; Hachim, A.; Kedzierska, K.; Valkenburg, S.A. T Cells Targeting SARS-CoV-2: By Infection, Vaccination, and Against Future Variants. Front. Med. 2021, 8, 793102. [Google Scholar] [CrossRef]
- Niessl, J.; Sekine, T.; Buggert, M. T cell immunity to SARS-CoV-2. Semin. Immunol. 2021, 55, 101505. [Google Scholar] [CrossRef] [PubMed]
- Lafuente, E.; Reche, P. Prediction of MHC-Peptide Binding: A Systematic and Comprehensive Overview. Curr. Pharm. Des. 2009, 15, 3209–3220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. npj Vaccines 2021, 6, 28. [Google Scholar] [CrossRef]
- Cheng, H.; Peng, Z.; Luo, W.; Si, S.; Mo, M.; Zhou, H.; Xin, X.; Liu, H.; Yu, Y. Efficacy and safety of covid-19 vaccines in phase iii trials: A meta-analysis. Vaccines 2021, 9, 582. [Google Scholar] [CrossRef] [PubMed]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; COVID-19 Genomics UK (COG-UK) Consortium; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Hacisuleyman, E.; Hale, C.; Saito, Y.; Blachere, N.E.; Bergh, M.; Conlon, E.G.; Schaefer-Babajew, D.J.; DaSilva, J.; Muecksch, F.; Gaebler, C.; et al. Vaccine Breakthrough Infections with SARS-CoV-2 Variants. N. Engl. J. Med. 2021, 384, 2212–2218. [Google Scholar] [CrossRef]
- Chaguza, C.; Coppi, A.; Earnest, R.; Ferguson, D.; Kerantzas, N.; Warner, F.; Young, H.P.; Breban, M.I.; Billig, K.; Koch, R.T.; et al. Rapid Emergence of SARS-CoV-2 Omicron Variant Is Associated with an Infection Advantage over Delta in Vaccinated Persons. medRxiv, 2022; online ahead of print. [Google Scholar] [CrossRef]
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning Immune Humoral Response to BNT162b2 Covid-19 Vaccine over 6 Months. N. Engl. J. Med. 2021, 385, e84. [Google Scholar] [CrossRef]
- Volz, E.; Hill, V.; McCrone, J.T.; Price, A.; Jorgensen, D.; O’Toole, Á.; Southgate, J.; Johnson, R.; Jackson, B.; Nascimento, F.; et al. Evaluating the Effects of SARS-CoV-2 Spike Mutation D614G on Transmissibility and Pathogenicity. Cell 2021, 184, 64–75.e11. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.J.; Chiba, S.; Halfmann, P.; Ehre, C.; Kuroda, M.; Dinnon, K.H.; Leist, S.R.; Schäfer, A.; Nakajima, N.; Takahashi, K.; et al. SARS-CoV-2 D614G variant exhibits efficient replication ex vivo and transmission in vivo. Science 2020, 370, 1464–1468. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827.e19. [Google Scholar] [CrossRef]
- Starr, T.N.; Greaney, A.J.; Hilton, S.K.; Ellis, D.; Crawford, K.H.D.; Dingens, A.S.; Navarro, M.; Bowen, J.; Tortorici, M.; Walls, A.; et al. Deep Mutational Scanning of SARS-CoV-2 Receptor Binding Domain Reveals Constraints on Folding and ACE2 Binding. Cell 2020, 182, 1295–1310.e20. [Google Scholar] [CrossRef] [PubMed]
- Teruel, N.; Mailhot, O.; Najmanovich, R.J. Modelling conformational state dynamics and its role on infection for SARS-CoV-2 Spike protein variants. PLoS Comput. Biol. 2021, 17, e1009286. [Google Scholar] [CrossRef]
- Nelson, G.; Buzko, O.; Patricia, S.; Niazi, K.; Rabizadeh, S.; Soon-Shiong, P. Molecular dynamic simulation reveals E484K mutation enhances spike RBD-ACE2 affinity and the 1 combination of E484K, K417N and N501Y mutations (501Y.V2 variant) induces conformational 2 change greater than N501Y mutant alone, potentially resulting in an escape mutant. bioRxiv 2021. [Google Scholar] [CrossRef]
- Cherian, S.; Potdar, V.; Jadhav, S.; Yadav, P.; Gupta, N.; Das, M.; Rakshit, P.; Singh, S.; Abraham, P.; Panda, S.; et al. SARS-CoV-2 Spike Mutations, L452R, T478K, E484Q and P681R, in the Second Wave of COVID-19 in Maharashtra, India. Microorganisms 2021, 9, 1542. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.G.; Tang, Y.; Akter, S.; Zheng, C. Roles of the polybasic furin cleavage site of spike protein in SARS-CoV-2 replication, pathogenesis, and host immune responses and vaccination. J. Med. Virol. 2021, 94, 1815–1820. [Google Scholar] [CrossRef]
- Gong, S.Y.; Chatterjee, D.; Richard, J.; Prévost, J.; Tauzin, A.; Gasser, R.; Bo, Y.; Vézina, D.; Goyette, G.; Gendron-Lepage, G.; et al. Contribution of single mutations to selected SARS-CoV-2 emerging variants spike antigenicity. Virology 2021, 563, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Zahradník, J.; Marciano, S.; Shemesh, M.; Zoler, E.; Harari, D.; Chiaravalli, J.; Meyer, B.; Rudich, Y.; Li, C.; Marton, I.; et al. SARS-CoV-2 variant prediction and antiviral drug design are enabled by RBD in vitro evolution. Nat. Microbiol. 2021, 6, 1188–1198. [Google Scholar] [CrossRef]
- Barnes, C.O.; Jette, C.A.; Abernathy, M.E.; Dam, K.M.A.; Esswein, S.R.; Gristick, H.B.; Malyutin, A.; Sharaf, N.; Huey-Tubman, K.; Lee, Y.; et al. SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies. Nature 2020, 588, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Mathew, N.R.; Angeletti, D. Recombinant Influenza Vaccines: Saviors to Overcome Immunodominance. Front. Immunol. 2020, 10, 2997. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.K.; Crotty, S. Factors in B cell competition and immunodominance. Immunol. Rev. 2020, 296, 120–131. [Google Scholar] [CrossRef]
- Angeletti, D.; Yewdell, J.W. Understanding and Manipulating Viral Immunity: Antibody Immunodominance Enters Center Stage. Trends Immunol. 2018, 39, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, A.A.; Kirchenbaum, G.A.; Zhang, T.; Reche, P.A.; Lehmann, P.V. Deconvoluting the T Cell Response to SARS-CoV-2: Specificity Versus Chance and Cognate Cross-Reactivity. Front. Immunol. 2021, 12, 635942. [Google Scholar] [CrossRef]
- Kim, A.R.; Sadegh-Nasseri, S. Determinants of immunodominance for CD4 T cells. Curr. Opin. Immunol. 2015, 34, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yewdell, J.W. Confronting Complexity: Real-World Immunodominance in Antiviral CD8+ T Cell Responses. Immunity 2006, 25, 533–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, C.K.V.; Franco, M.M.; Gräf, T.; De Lorenzo Barcia, C.A.; De Ávila Mendonça, R.N.; De Sousa, K.A.F.; Neiva, L.; Fosenca, V.; Mendes, A.; de Aguiar, R.; et al. Genomic evidence of SARS-CoV-2 reinfection involving E484K spike mutation, Brazil. Emerg. Infect. Dis. 2021, 27, 1522–1524. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Nair, M.S.; Liu, L.; Iketani, S.; Luo, Y.; Guo, Y.; Wang, M.; Yu, J.; Zhang, B.; Kwong, P.D.; et al. Antibody resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7. Nature 2021, 593, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Arora, P.; Groß, R.; Seidel, A.; Hörnich, B.F.; Hahn, A.S.; Krüger, N.; Graichen, L.; Hofmann-Winkler, H.; Kempf, A.; et al. SARS-CoV-2 variants B.1.351 and P.1 escape from neutralizing antibodies. Cell 2021, 184, 2384–2393.e12. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Madhi, S.A.; Izu, A.; Pollard, A.J. ChAdOx1 nCoV-19 Vaccine Efficacy against the B.1.351 Variant. Reply. N. Engl. J. Med. 2021, 385, 571–572. [Google Scholar]
- Madhi, S.A.; Baillie, V.; Cutland, C.L.; Voysey, M.; Koen, A.L.; Fairlie, L.; Padayachee, S.D.; Dheda, K.; Barnabas, S.L.; Bhorat, Q.E.; et al. Efficacy of the ChAdOx1 nCoV-19 Covid-19 Vaccine against the B.1.351 Variant. N. Engl. J. Med. 2021, 384, 1885–1898. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Tang, H.; McDanal, C.; Wagh, K.; Fischer, W.; Theiler, J.; Yoon, H.; Li, D.; Haynes, B.; Saunders, K.; et al. SARS-CoV-2 variant B.1.1.7 is susceptible to neutralizing antibodies elicited by ancestral spike vaccines. Cell Host Microbe 2021, 29, 529–539.e3. [Google Scholar] [CrossRef]
- Wadman, M. Novavax vaccine delivers 89% efficacy against COVID-19 in U.K.—But is less potent in South Africa. Science 2021, 12, 277. [Google Scholar] [CrossRef]
- Rayati Damavandi, A.; Dowran, R.; Al Sharif, S.; Kashanchi, F.; Jafari, R. Molecular variants of SARS-CoV-2: Antigenic properties and current vaccine efficacy. Med. Microbiol. Immunol. 2022. [Google Scholar] [CrossRef]
- Mistry, P.; Barmania, F.; Mellet, J.; Peta, K.; Strydom, A.; Viljoen, I.M.; James, W.; Gordon, S.; Pepper, M. SARS-CoV-2 Variants, Vaccines, and Host Immunity. Front. Immunol. 2022, 12, 809244. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Enhancing response to Omicron SARS-CoV-2 Variant. Available online: https://www.who.int/publications/m/item/enhancing-readiness-for-omicron-(b.1.1.529)-technical-brief-and-priority-actions-for-member-states (accessed on 22 April 2022).
- Carreño, J.M.; Alshammary, H.; Tcheou, J.; Singh, G.; Raskin, A.; Kawabata, H.; Sominsky, L.A.; Clark, J.J.; Adelsberg, D.C.; Bielak, D.A.; et al. Activity of convalescent and vaccine serum against SARS-CoV-2 Omicron. Nature 2021, 602, 682–688. [Google Scholar] [CrossRef]
- Cele, S.; Jackson, L.; Khoury, D.S.; Khan, K.; Moyo-Gwete, T.; Tegally, H.; San, J.; Cromer, D.; Scheepers, C.; Amoako, D.; et al. Omicron extensively but incompletely escapes Pfizer BNT162b2 neutralization. Nature 2021, 602, 654–656. [Google Scholar] [CrossRef]
- Fiolet, T.; Kherabi, Y.; Macdonald, C.; Ghosn, J.; Peiffer-smadja, N. Comparing COVID-19 vaccines for their characteristics, efficacy and effectiveness against SARS-CoV-2 and variants of concern: A narrative review. Clin. Microbiol. Infect. 2022, 28, 202–221. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Huo, J.; Zhou, D.; Zahradník, J.; Supasa, P.; Liu, C.; Duyvesteyn, H.; Ginn, H.M.; Mentzer, A.J.; Tuekprakhon, A.; et al. SARS-CoV-2 Omicron-B.1.1.529 leads to widespread escape from neutralizing antibody responses. Cell 2022, 185, 467–484.e15. [Google Scholar] [CrossRef] [PubMed]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.M.; et al. Covid-19 Vaccine Effectiveness against the Omicron (B.1.1.529) Variant. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef] [PubMed]
- Ewer, K.J.; Barrett, J.R.; Belij-Rammerstorfer, S.; Sharpe, H.; Makinson, R.; Morter, R.; Flaxman, A.; Wright, D.; Bellamy, D.; Bittaye, M.; et al. T cell and antibody responses induced by a single dose of ChAdOx1 nCoV-19 (AZD1222) vaccine in a phase 1/2 clinical trial. Nat. Med. 2021, 27, 270–278. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- Moss, P. The T cell immune response against SARS-CoV-2. Vol. 23, Nature Immunology. Nat. Res. 2022, 23, 186–193. [Google Scholar]
- Reche, P.A.; Reinherz, E.L. Sequence variability analysis of human class I and class II MHC molecules: Functional and structural correlates of amino acid polymorphisms. J. Mol. Biol. 2003, 331, 623–641. [Google Scholar] [CrossRef] [Green Version]
- Tarke, A.; Coelho, C.H.; Zhang, Z.; Dan, J.M.; Yu, E.D.; Methot, N.; Bloom, N.I.; Goodwin, B.; Phillips, E.; Mallal, S.; et al. SARS-CoV-2 vaccination induces immunological T cell memory able to cross-recognize variants from Alpha to Omicron. Cell 2022, 185, 847–859.e11. [Google Scholar] [CrossRef] [PubMed]
- Barandalla, I.; Alvarez, C.; Barreiro, P.; de Mendoza, C.; González-Crespo, R.; Soriano, V. Impact of scaling up SARS-CoV-2 vaccination on COVID-19 hospitalizations in Spain. Int. J. Infect. Dis. 2021, 112, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Monereo-Sánchez, J.; Luykx, J.J.; Pinzón-Espinosa, J.; Richard, G.; Motazedi, E.; Westlye, L.T.; Andreassen, O.A.; van der Meer, D. Diphtheria and Tetanus Vaccination History Is Associated With Lower Odds of COVID-19 Hospitalization. Front. Immunol. 2021, 12, 749264. [Google Scholar] [CrossRef]
- Mysore, V. Clinical and Translational Article Protective heterologous T cell immunity in COVID-19 induced by the trivalent MMR and Tdap vaccine antigens. Med 2021, 2, 1050–1071.e7. [Google Scholar] [CrossRef]
- Reche, P.A. Potential Cross-Reactive Immunity to SARS-CoV-2 From Common Human Pathogens and Vaccines. Front. Immunol. 2020, 11, 586984. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, J.S.; Bilich, T.; Tandler, C.; Nelde, A.; Maringer, Y.; Marconato, M.; Reusch, J.; Jäger, S.; Denk, M.; Richter, M.; et al. A COVID-19 peptide vaccine for the induction of SARS-CoV-2 T cell immunity. Nature 2022, 601, 617–622. [Google Scholar] [CrossRef]
- Guo, L.; Wang, G.; Wang, Y.; Zhang, Q.; Ren, L.; Gu, X.; Huang, T.; Zhong, J.; Wang, Y.; Wang, X.; et al. SARS-CoV-2-specific antibody and T-cell responses 1 year after infection in people recovered from COVID-19: A longitudinal cohort study. Lancet Microbe, 2022; online ahead of print. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Name | GISAID Accession ID | Lineage | Location | Collection Date |
|---|---|---|---|---|
| hCoV-19/England/ QEUH-119AACD/2021 | EPI_ISL_944882 | B.1.1.7 (Alpha) | United Kingdom/England | 28 January 2021 |
| hCoV-19/South Africa/N00390/2020 | EPI_ISL_712081 | B.1.315 (Beta) | South Africa/Eastern Cape | 8 October 2020 |
| hCoV-19/Brazil/AM-1031/2021 | EPI_ISL_906075 | P.1 (Gamma) | Brazil/Manaus | 19 January 2021 |
| hCoV-19/India/TN-SEQ_5119_S219_R1_001/2021 | EPI_ISL_6033571 | B.1.617.2 (Delta) | India/Tamil Nadu | 26 July 2021 |
| hCoV-19/Nepal/NPHL-S-263/2021 | EPI_ISL_7196120 | B.1.1.529 (Omicron) | Nepal/Bagmati | 26 November 2021 |
| Variant | Amino Acid Substitutions | Amino Acid Deletions |
|---|---|---|
| B.1.1.7 (Alpha) | S13T, T385N, N501Y, A570D, D614G, P681H, T716I, S982A, D1118H | H69-, V70-, Y144- |
| B.1.315 (Beta) | D80A, R246I, K417N, E484K, N501Y, D614G, A701V | L241-, L242-, A243- |
| P.1 (Gamma) | L18F, T20N, P26S, D138Y, R190S, K417T, E484K, N501Y, D614G, H655Y, T1027I, V1176F | - |
| B.1.617.2 (Delta) | T19R, G142D, L452R, T478K, E484Q, D614G, P681R, P812T, D950N | - |
| B. 1.1.529 (Omicron) | A67V, T95I, G339D, S371L, S373P, S375F, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H, T547K, D614G, H655Y, N679K, P681H, D796Y, N856K, Q954H, N969K, L981F | - |
| Cluster 1 | Epitope Core | N° Mutations | ||||
|---|---|---|---|---|---|---|
| α | β | γ | δ | ο | ||
| S12-R246 | S12,C15,L18,T19,C136,G142,H146,K147,N149,R246 | 0 | 1 | 1 | 2 | 0 |
| Q14-D253 | Q14,Y144,Y145,H146,K147,F157,G252,D253 | 1 | 0 | 0 | 0 | 0 |
| V16-D253 | V16,Y144,K147,R246,P251,G252,D253 | 0 | 1 | 0 | 0 | 0 |
| L18-T716 | L18,Y144,L242,A243,L244,R246,E484,N501,A701,T716 | 2 | 6 | 3 | 1 | 2 |
| L18-V1176 | L18,T20,P26,D138,R190,K417,E484,N501,D614,H655,T1027,V1176 | 2 | 4 | 12 | 2 | 4 |
| H69-G261 | H69,V70,Y144,L242,A243,L244,G261 | 2 | 2 | 0 | 0 | 0 |
| D80-A701 | D80,D215,K417,E484,N501,D614,A701 | 2 | 6 | 4 | 2 | 3 |
| F140-H245 | F140,G142,V143,Y145,H146,N148,N149,W152,E154,F157,A243,L244,H245 | 1 | 1 | 0 | 1 | 0 |
| Y144-Y248 | Y144,Y145,H146,K147,K150,W152,Y248 | 1 | 0 | 0 | 0 | 0 |
| Y144-P251 | Y144,H146,K147,R246,Y248,L249,P251 | 0 | 1 | 0 | 0 | 0 |
| Y144-D253 | Y144,Y145,H146,K147,R158,R246,L249,T250,P251,G252,D253 | 1 | 1 | 0 | 0 | 0 |
| Y144-S255 | Y144,R246,S247,Y248,T250,P251,D253,S254,S255 | 0 | 1 | 0 | 0 | 0 |
| Y145-G261 | Y145,H146,K147,R246,Y248,P251,G261 | 1 | 1 | 0 | 0 | 0 |
| N334-L441 | N334,L335,P337,G339,E340,N343,A344,T345,K356,R357,S359,C361,L441 | 0 | 0 | 0 | 0 | 1 |
| G339-L441 | G339,E340,N343,A344,T345,R346,L441 | 0 | 0 | 0 | 0 | 1 |
| T342-F487 | T342,R343,Y348,Y446,N447,L449,I465,T467,F487 | 0 | 0 | 0 | 1 | 0 |
| R343-T497 | R343,N436,N437,L438,K441,V442,G443,G444,P496,T497 | 0 | 0 | 0 | 0 | 0 |
| T345-V445 | T345,R346,S373,W436,N437,N440,L441,K444,V445 | 0 | 0 | 0 | 0 | 1 |
| T345-V455-N450 | T345,R346,N440,L441,S443,K444,V445,N450 | 0 | 0 | 0 | 0 | 0 |
| T345-Y449-N450 | T345,R346,K444,G446,G447,Y449,N450 | 0 | 0 | 0 | 0 | 0 |
| T345-S494 | T345,R346,A348,S349,N354,K356,Y449,N450,L452,T470,Q493,S494 | 0 | 0 | 0 | 1 | 1 |
| R346-Q498 | R346,N440,L441,K444,V445,G446,N448,Y449,Q498 | 0 | 0 | 0 | 0 | 1 |
| Y351-R509 | Y351,A352,W353,N360,L368,A419,V433,Y449,N450,D467,C480,E484,C488,F490,S494,R509 | 0 | 1 | 1 | 1 | 1 |
| W353-R466 | W353,R355,R357,Y396,K462,F464,R466 | 0 | 0 | 0 | 0 | 0 |
| R355-R466 | R355,R357,D428,K462,P463,F464,R466 | 0 | 0 | 0 | 0 | 0 |
| R355-I498 | R355,R457,K462,F464,E465,R466,D467,I468 | 0 | 0 | 0 | 0 | 0 |
| F374-P384 | F374,S375,T376,F377,K378,C379,P384 | 0 | 0 | 0 | 0 | 1 |
| F374-E516 | F374,S375,T376,F377,C379,F392,D427,E516 | 0 | 0 | 0 | 0 | 1 |
| R403-R509 | R403,D405,A419,G502,G504,Y505,R509 | 0 | 0 | 0 | 0 | 1 |
| T415-G502 | T415,Y421,A475,G476,N487,S494,G502 | 0 | 0 | 0 | 0 | 0 |
| K417-Y489 | K417,L455,F456,A475,E484,F486,Y489 | 0 | 2 | 2 | 1 | 1 |
| K417-N501 | K417,Y449,L455,F456,Y489,G496,Q498,T500,N501 | 1 | 2 | 2 | 0 | 3 |
| S443-S494 | S443,V445,G446,G447,Y449,P499,T500 | 0 | 0 | 0 | 0 | 0 |
| K444-N450-S494 | K444,V445,G446,G447,Y449,N450,S494 | 0 | 0 | 0 | 0 | 0 |
| K444-I472-S494 | K444,V445,G446,L452,L455,F456,T470,E471,I472,S494 | 0 | 0 | 0 | 1 | 0 |
| K444-T500 | K444,V445,G446,N450,Q498,P499,T500 | 0 | 0 | 0 | 0 | 1 |
| V445-G502 | V445,G446,Q498,P499,T500,N501,G502 | 1 | 1 | 1 | 0 | 2 |
| G446-G447-F490 | G446,G447,N448,Y449,N450,L452,E484,F490 | 0 | 1 | 1 | 2 | 1 |
| G446-Y449-F490 | G446,Y449,L452,V483,E484,G485,F490 | 0 | 1 | 1 | 2 | 1 |
| G446-S494 | G446,Y449,E484,F490,L492,Q493,S494 | 0 | 1 | 1 | 1 | 2 |
| Y449-F490 | Y449,I472,N481,V483,E484,G485,F486,F490 | 0 | 1 | 1 | 1 | 1 |
| Y449-L492 | Y449,N481,G482,V483,E484,F490,L492 | 0 | 1 | 1 | 1 | 1 |
| Y449-Q493 | Y449,E484,G485,F486,Y489,F490,Q493 | 0 | 1 | 1 | 1 | 2 |
| Y449-L452-S494 | Y449,L452,T470,E484,F490,L492,S494 | 0 | 1 | 1 | 2 | 1 |
| Y449-N481-S494 | Y449,N481,G482,V483,E484,F490,S494 | 0 | 1 | 1 | 1 | 1 |
| Y449-Y505 | Y449,Y453,L455,F456,Y489,Q493,Y495,Y505 | 0 | 0 | 0 | 0 | 2 |
| L452-Q490 | L452,F453,T475,G482,F483,N484,Y486,Q490 | 0 | 0 | 0 | 1 | 2 |
| L455-Y489 | L455,F456,A475,T478,E484,F486,N487,Y489 | 0 | 1 | 1 | 2 | 2 |
| L455-F490 | L455,V483,E484,G485,F486,Y489,F490 | 0 | 1 | 1 | 1 | 1 |
| L455-Q493 | L455,A475,T478,G485,F486,Y489,Q493 | 0 | 0 | 0 | 1 | 2 |
| F456-Q493 | F456,E484,G485,F486,C488,Y489,Q493 | 0 | 1 | 1 | 1 | 2 |
| Y473-F486 | Y473,A475,G476,S477,E484,G485,F486 | 0 | 1 | 1 | 1 | 2 |
| N556-D568 | N556,K557,K558,L560,P561,F562,Q563,D568 | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballesteros-Sanabria, L.; Pelaez-Prestel, H.F.; Ras-Carmona, A.; Reche, P.A. Resilience of Spike-Specific Immunity Induced by COVID-19 Vaccines against SARS-CoV-2 Variants. Biomedicines 2022, 10, 996. https://doi.org/10.3390/biomedicines10050996
Ballesteros-Sanabria L, Pelaez-Prestel HF, Ras-Carmona A, Reche PA. Resilience of Spike-Specific Immunity Induced by COVID-19 Vaccines against SARS-CoV-2 Variants. Biomedicines. 2022; 10(5):996. https://doi.org/10.3390/biomedicines10050996
Chicago/Turabian StyleBallesteros-Sanabria, Laura, Hector F. Pelaez-Prestel, Alvaro Ras-Carmona, and Pedro A. Reche. 2022. "Resilience of Spike-Specific Immunity Induced by COVID-19 Vaccines against SARS-CoV-2 Variants" Biomedicines 10, no. 5: 996. https://doi.org/10.3390/biomedicines10050996
APA StyleBallesteros-Sanabria, L., Pelaez-Prestel, H. F., Ras-Carmona, A., & Reche, P. A. (2022). Resilience of Spike-Specific Immunity Induced by COVID-19 Vaccines against SARS-CoV-2 Variants. Biomedicines, 10(5), 996. https://doi.org/10.3390/biomedicines10050996