
Inhibiting Transglutaminase 2 Mediates Kidney Fibrosis via Anti-Apoptosis

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Unilateral Ureteral Obstruction (UUO) Animal Model
2.2. 5/6 Nephrectomy (5/6 Nx.) Animal Model
2.3. Terminal Deoxynucleotidyl Transferase dUTP Nick-End Labeling (TUNEL) Assay
2.4. Human Samples
2.5. Quantitative Real-Time PCR
2.6. Histopathological Evaluations
2.7. Immunohistochemical Evaluation of TG2
2.8. Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Human Primary Tubular Epithelial Cell (hTEC) Preparation
2.10. Cell Apoptosis Assay
2.11. Western Blot Analysis
2.12. Statistical Analysis
3. Results
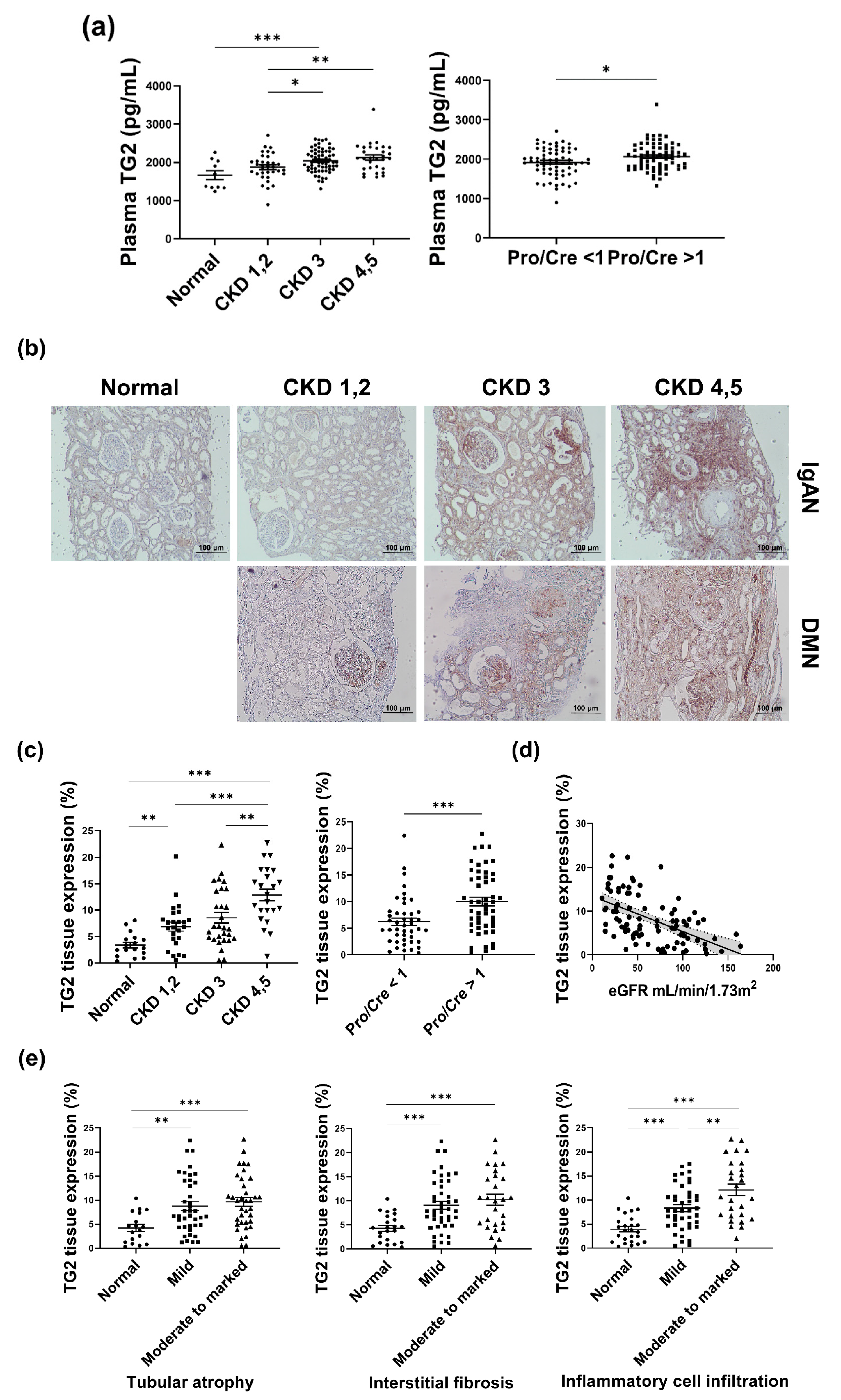
3.1. TG2 Levels Are Increased with Deterioration of the Kidney in Human Samples
3.2. TG2 Inhibition Using Cystamine Decreased rTGFβ-Induced Apoptosis and Suppressed Fibrotic Changes in Primary Cultures of Tubular Epithelial Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jha, V.; Garcia-Garcia, G.; Iseki, K.; Li, Z.; Naicker, S.; Plattner, B.; Saran, R.; Wang, A.Y.-M.; Yang, C.-W. Chronic kidney disease: Global dimension and perspectives. Lancet 2013, 382, 260–272. [Google Scholar] [CrossRef]
- Hill, N.R.; Fatoba, S.T.; Oke, J.L.; Hirst, J.A.; O’Callaghan, C.A.; Lasserson, D.S.; Hobbs, F.D. Global Prevalence of Chronic Kidney Disease-A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0158765. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Ortega, M.; Rayego-Mateos, S.; Lamas, S.; Ortiz, A.; Rodrigues-Diez, R.R. Targeting the progression of chronic kidney disease. Nat. Rev. Nephrol. 2020, 16, 269–288. [Google Scholar] [CrossRef]
- Min, B.; Chung, K.C. New insight into transglutaminase 2 and link to neurodegenerative diseases. BMB Rep. 2018, 51, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Di Sabatino, A.; Vanoli, A.; Giuffrida, P.; Luinetti, O.; Solcia, E.; Corazza, G.R. The function of tissue transglutaminase in celiac disease. Autoimmun. Rev. 2012, 11, 746–753. [Google Scholar] [CrossRef]
- Johnson, T.S.; El-Koraie, A.F.; Skill, N.J.; Baddour, N.M.; El Nahas, A.M.; Njloma, M.; Adam, A.G.; Griffin, M. Tissue transglutaminase and the progression of human renal scarring. J. Am. Soc. Nephrol. 2003, 14, 2052–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elli, L.; Bergamini, C.M.; Bardella, M.T.; Schuppan, D. Transglutaminases in inflammation and fibrosis of the gastrointestinal tract and the liver. Dig. Liver Dis. 2009, 41, 541–550. [Google Scholar] [CrossRef]
- Johnson, T.S.; Skill, N.J.; Nahas, A.M.E.; Oldroyd, S.D.; Thomas, G.L.; Douthwaite, J.A.; Haylor, J.L.; Griffin, M. Transglutaminase transcription and antigen translocation in experimental renal scarring. J. Am. Soc. Nephrol. 1999, 10, 2146–2157. [Google Scholar] [CrossRef]
- Scarpellini, A.; Huang, L.; Burhan, I.; Schroeder, N.; Funck, M.; Johnson, T.S.; Verderio, E.A. Syndecan-4 knockout leads to reduced extracellular transglutaminase-2 and protects against tubulointerstitial fibrosis. J. Am. Soc. Nephrol. 2014, 25, 1013–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furini, G.; Schroeder, N.; Huang, L.; Boocock, D.; Scarpellini, A.; Coveney, C.; Tonoli, E.; Ramaswamy, R.; Ball, G.; Verderio, C.; et al. Proteomic Profiling Reveals the Transglutaminase-2 Externalization Pathway in Kidneys after Unilateral Ureteric Obstruction. J. Am. Soc. Nephrol. 2018, 29, 880–905. [Google Scholar] [CrossRef] [Green Version]
- Priante, G.; Gianesello, L.; Ceol, M.; Del Prete, D.; Anglani, F. Cell Death in the Kidney. Int. J. Mol. Sci. 2019, 20, 3598. [Google Scholar] [CrossRef] [Green Version]
- Tatsukawa, H.; Furutani, Y.; Hitomi, K.; Kojima, S. Transglutaminase 2 has opposing roles in the regulation of cellular functions as well as cell growth and death. Cell Death Dis. 2016, 7, e2244. [Google Scholar] [CrossRef] [Green Version]
- Caccamo, D.; Curro, M.; Ferlazzo, N.; Condello, S.; Ientile, R. Monitoring of transglutaminase 2 under different oxidative stress conditions. Amino Acids 2012, 42, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Mirza, A.; Liu, S.L.; Frizell, E.; Zhu, J.; Maddukuri, S.; Martinez, J.; Davies, P.; Schwarting, R.; Norton, P.; Zern, M.A. A role for tissue transglutaminase in hepatic injury and fibrogenesis, and its regulation by NF-KB. Am. J. Physiol. 1997, 272 Pt 1, G281–G288. [Google Scholar] [PubMed]
- Kuncio, G.S.; Tsyganskaya, M.; Zhu, J.; Liu, S.L.; Nagy, L.; Thomazy, V.; Davies, P.J.; Zern, M.A. TNF-alpha modulates expression of the tissue transglutaminase gene in liver cells. Am. J. Physiol. 1998, 274, G240–G245. [Google Scholar]
- Singh, U.S.; Pan, J.; Kao, Y.L.; Joshi, S.; Young, K.L.; Baker, K.M. Tissue transglutaminase mediates activation of RhoA and MAP kinase pathways during retinoic acid-induced neuronal differentiation of SH-SY5Y cells. J. Biol. Chem. 2003, 278, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.; Zhang, Z.; Cohen, D.M. MAPK signaling and the kidney. Am. J. Physiol. Renal Physiol. 2000, 279, F593–F604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, N.; Thaiss, F.; Guo, L. NFkappaB and Kidney Injury. Front. Immunol. 2019, 10, 815. [Google Scholar] [CrossRef]
- Burhan, I.; Furini, G.; Lortat-Jacob, H.; Atobatele, A.G.; Scarpellini, A.; Schroeder, N.; Atkinson, J.; Maamra, M.; Nutter, F.H.; Watson, P.; et al. Interplay between transglutaminases and heparan sulphate in progressive renal scarring. Sci. Rep. 2016, 6, 31343. [Google Scholar] [CrossRef] [Green Version]
- Zakrzewicz, A.; Atanasova, S.; Padberg, W.; Grau, V. Monocytic Tissue Transglutaminase in a Rat Model for Reversible Acute Rejection and Chronic Renal Allograft Injury. Mediat. Inflamm. 2015, 2015, 429653. [Google Scholar] [CrossRef]
- Da Silva Lodge, M.; Pullen, N.; Pereira, M.; Johnson, T.S. Urinary levels of pro-fibrotic transglutaminase 2 (TG2) may help predict progression of chronic kidney disease. PLoS ONE 2022, 17, e0262104. [Google Scholar] [CrossRef]
- Jeitner, T.M.; Pinto, J.T.; Cooper, A.J. Cystamine and cysteamine as inhibitors of transglutamiunase activity in vivo. Biosci. Rep. 2018, 38, BSR20180691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsukawa, H.; Otsu, R.; Tani, Y.; Wakita, R.; Hitomi, K. Isozyme-specific comprehensive characterization of transglutaminase-crosslinked substrates in kidney fibrosis. Sci. Rep. 2018, 8, 7306. [Google Scholar] [CrossRef] [Green Version]
- Okamura, D.M.; Bahrami, N.M.; Ren, S.; Pasichnyk, K.; Williams, J.M.; Gangoiti, J.A.; Lopez-Guisa, J.M.; Yamaguchi, I.; Barshop, B.A.; Duffield, J.S.; et al. Cysteamine modulates oxidative stress and blocks myofibroblast activity in CKD. J. Am. Soc. Nephrol. 2014, 25, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, S.; Koide, R.; Shimohata, T.; Yamada, M.; Hayashi, Y.; Takano, H.; Date, H.; Oyake, M.; Sato, T.; Sato, A.; et al. Suppression of aggregate formation and apoptosis by transglutaminase inhibitors in cells expressing truncated DRPLA protein with an expanded polyglutamine stretch. Nat. Genet. 1998, 18, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.; Haendeler, J.; Aebly, M.R.; Kelly, L.A.; Cholewa, B.C.; Koike, G.; Kwitek-Black, A.; Jacob, H.J.; Berk, B.C.; Miano, J.M. Retinoic acid-induced tissue transglutaminase and apoptosis in vascular smooth muscle cells. Circ. Res. 2000, 87, 881–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Haylor, J.L.; Hau, Z.; Jones, R.A.; Vickers, M.E.; Wagner, B.; Griffin, M.; Saint, R.E.; Coutts, I.G.C.; El Nahas, A.M.; et al. Transglutaminase inhibition ameliorates experimental diabetic nephropathy. Kidney Int. 2009, 76, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.S.; Fisher, M.; Haylor, J.L.; Hau, Z.; Skill, N.J.; Jones, R.; Saint, R.; Coutts, I.; Vickers, M.E.; El Nahas, A.M.; et al. Transglutaminase inhibition reduces fibrosis and preserves function in experimental chronic kidney disease. J. Am. Soc. Nephrol. 2007, 18, 3078–3088. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, Y.; Kim, K.H.; Kim, D.K.; Joo, K.W.; Shin, S.J.; Kim, Y.S.; Yang, S.H. Chemokine (C-C Motif) Ligand 8 and Tubulo-Interstitial Injury in Chronic Kidney Disease. Cells 2022, 11, 658. [Google Scholar] [CrossRef]
- Lee, J.W.; Bae, E.; Kwon, S.H.; Yu, M.Y.; Cha, R.H.; Lee, H.; Kim, D.K.; Lee, J.P.; Ye, S.K.; Yoo, J.Y.; et al. Transcriptional modulation of the T helper 17/interleukin 17 axis ameliorates renal ischemia-reperfusion injury. Nephrol. Dial. Transplant. 2019, 34, 1481–1498. [Google Scholar] [CrossRef]
- Iwai, N.; Shimoike, H.; Kinoshita, M. Genes up-regulated in hypertrophied ventricle. Biochem. Biophys. Res. Commun. 1995, 209, 527–534. [Google Scholar] [CrossRef]
- Small, K.; Feng, J.F.; Lorenz, J.; Donnelly, E.T.; Yu, A.; Im, M.J.; Dorn, G.W., II; Liggett, S.B. Cardiac specific overexpression of transglutaminase II (G(h)) results in a unique hypertrophy phenotype independent of phospholipase C activation. J. Biol. Chem. 1999, 274, 21291–21296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Stuckey, D.J.; Murdoch, C.E.; Camelliti, P.; Lip, G.Y.H.; Griffin, M. Cardiac fibrosis can be attenuated by blocking the activity of transglutaminase 2 using a selective small-molecule inhibitor. Cell Death Dis. 2018, 9, 613. [Google Scholar] [CrossRef] [Green Version]
- Olsen, K.C.; Sapinoro, R.E.; Kottmann, R.M.; Kulkarni, A.A.; Iismaa, S.E.; Johnson, G.V.; Thatcher, T.H.; Phipps, R.P.; Sime, P.J. Transglutaminase 2 and its role in pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2011, 184, 699–707. [Google Scholar] [CrossRef] [Green Version]
- Nardacci, R.; Ciccosanti, F.; Falasca, L.; Lo Iacono, O.; Amendola, A.; Antonucci, G.; Piacentini, M. Tissue transglutaminase in HCV infection. Cell Death Differ. 2003, 10 (Suppl. 1), S79–S80. [Google Scholar] [CrossRef] [Green Version]
- Boor, P.; Ostendorf, T.; Floege, J. Renal fibrosis: Novel insights into mechanisms and therapeutic targets. Nat. Rev. Nephrol. 2010, 6, 643–656. [Google Scholar] [CrossRef]
- Greenberg, C.S.; Birckbichler, P.J.; Rice, R.H. Transglutaminases: Multifunctional cross-linking enzymes that stabilize tissues. FASEB J. 1991, 15, 3071–3077. [Google Scholar] [CrossRef]
- Nunes, I.; Gleizes, P.-E.; Metz, C.N.; Rifkin, D.B. Latent Transforming Growth Factor-β Binding Protein Domains Involved in Activation and Transglutaminase-dependent Cross-Linking of Latent Transforming Growth Factor-β. J. Cell Biol. 1997, 136, 1151–1163. [Google Scholar] [CrossRef] [PubMed]
- Okuda, S.; Languino, L.R.; Ruoslahti, E.; Border, W.A. Elevated expression of transforming growth factor-beta and proteoglycan production in experimental glomerulonephritis. Possible role in expansion of the mesangial extracellular matrix. J. Clin. Investig. 1990, 86, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Isaka, Y. Targeting TGF-beta Signaling in Kidney Fibrosis. Int. J. Mol. Sci. 2018, 19, 2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Perez, M.; Lee, E.S.; Kojima, S.; Griffin, M. The functional relationship between transglutaminase 2 and transforming growth factor beta1 in the regulation of angiogenesis and endothelial-mesenchymal transition. Cell Death Dis. 2017, 8, e3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katt, W.P.; Antonyak, M.A.; Cerione, R.A. Opening up about Tissue Transglutaminase: When Conformation Matters More than Enzymatic Activity. Med. One 2018, 3, 6. [Google Scholar]
- Philp, C.J.; Siebeke, I.; Clements, D.; Miller, S.; Habgood, A.; John, A.E.; Navaratnam, V.; Hubbard, R.B.; Jenkins, G.; Johnson, S.R. Extracellular Matrix Cross-Linking Enhances Fibroblast Growth and Protects against Matrix Proteolysis in Lung Fibrosis. Am. J. Respir. Cell Mol. Biol. 2018, 58, 594–603. [Google Scholar] [CrossRef]
- Park, K.S.; Han, B.G.; Lee, K.H.; Kim, D.S.; Kim, J.M.; Jeon, H.; Kim, H.S.; Suh, S.W.; Lee, E.H.; Kim, S.Y.; et al. Depletion of nucleophosmin via transglutaminase 2 cross-linking increases drug resistance in cancer cells. Cancer Lett. 2009, 274, 201–207. [Google Scholar] [CrossRef]
- Yuan, L.; Holmes, T.C.; Watts, R.E.; Khosla, C.; Broekelmann, T.J.; Mecham, R.; Zheng, H.; Izaguirre, E.W.; Rich, K.M. Novel chemo-sensitizing agent, ERW1227B, impairs cellular motility and enhances cell death in glioblastomas. J. Neurooncol. 2011, 103, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Tatsukawa, H.; Fukaya, Y.; Frampton, G.; Martinez-Fuentes, A.; Suzuki, K.; Kuo, T.F.; Nagatsuma, K.; Shimokado, K.; Okuno, M.; Wu, J.; et al. Role of transglutaminase 2 in liver injury via cross-linking and silencing of transcription factor Sp1. Gastroenterology 2009, 136, 1783–1795.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsukawa, H.; Sano, T.; Fukaya, Y.; Ishibashi, N.; Watanabe, M.; Okuno, M.; Moriwaki, H.; Kojima, S. Dual induction of caspase 3- and transglutaminase-dependent apoptosis by acyclic retinoid in hepatocellular carcinoma cells. Mol. Cancer 2011, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Oliverio, S.; Amendola, A.; Sano, F.D.; Farrace, M.G.; Fesus, L.; Nemes, Z.; Piredda, L.; Spinedi, A.; Piacentini, M. Tissue transglutaminase-dependent posttranslational modification of the retinoblastoma gene product in promonocytic cells undergoing apoptosis. Mol. Cell Biol. 1997, 17, 6040–6048. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.J.; Lee, K.B.; Son, Y.H.; Shin, J.; Lee, J.H.; Kim, H.J.; Hong, A.Y.; Bae, H.W.; Kwon, M.A.; Lee, W.J.; et al. Transglutaminase 2 mediates UV-induced skin inflammation by enhancing inflammatory cytokine production. Cell Death Dis. 2017, 8, e3148. [Google Scholar] [CrossRef] [PubMed]
- Bijli, K.M.; Kanter, B.G.; Minhajuddin, M.; Leonard, A.; Xu, L.; Fazal, F.; Rahman, A. Regulation of endothelial cell inflammation and lung polymorphonuclear lymphocyte infiltration by transglutaminase 2. Shock 2014, 42, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Su, T.; Qin, X.Y.; Furutani, Y. Transglutaminase 2 as a Marker for Inflammation and Therapeutic Target in Sepsis. Int. J. Mol. Sci. 2021, 22, 1897. [Google Scholar] [CrossRef] [PubMed]
- Shanmuganathan, R.; Ramanathan, K.; Padmanabhan, G.; Vijayaraghavan, B. Evaluation of Interleukin 8 gene polymorphism for predicting inflammation in Indian chronic kidney disease and peritoneal dialysis patients. Alex. J. Med. 2019, 53, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.K.; Estrella, M.M.; Appel, L.J.; Coresh, J.; Luo, S.; Obeid, W.; Parikh, C.R.; Grams, M.E. Serum levels of IL-6, IL-8 and IL-10 and risks of end-stage kidney disease and mortality. Nephrol. Dial. Transplant. 2021, 36, 561–563. [Google Scholar] [CrossRef]
- Bronze-da-Rocha, E.; Santos-Silva, A. Neutrophil Elastase Inhibitors and Chronic Kidney Disease. Int. J. Biol. Sci. 2018, 14, 1343–1360. [Google Scholar] [CrossRef] [Green Version]
- Du, S.H.; Zhang, W.; Yue, X.; Luo, X.Q.; Tan, X.H.; Liu, C.; Qiao, D.F.; Wang, H. Role of CXCR1 and Interleukin-8 in Methamphetamine-Induced Neuronal Apoptosis. Front. Cell Neurosci. 2018, 12, 230. [Google Scholar] [CrossRef]
- Kim, J.Y.; Wee, Y.M.; Choi, M.Y.; Jung, H.R.; Choi, J.Y.; Kwon, H.W.; Jung, J.H.; Cho, Y.M.; Go, H.; Han, M.; et al. Urinary transglutaminase 2 as a potent biomarker to predict interstitial fibrosis and tubular atrophy of kidney allograft during early posttransplant period in deceased donor kidney transplantation. Ann. Surg. Treat. Res. 2019, 97, 27–35. [Google Scholar] [CrossRef]
- Furini, G.; Burhan, I.; Huang, L.; Savoca, M.P.; Atobatele, A.; Johnson, T.; Verderio, E.A.M. Insights into the heparan sulphate-dependent externalisation of transglutaminase-2 (TG2) in glucose-stimulated proximal-like tubular epithelial cells. Anal. Biochem. 2020, 603, 113628. [Google Scholar] [CrossRef]
- Falasca, L.; Farrace, M.G.; Rinaldi, A.; Tuosto, L.; Melino, G.; Piacentini, M. Transglutaminase type II is involved in the pathogenesis of endotoxic shock. J. Immunol. 2008, 180, 2616–2624. [Google Scholar] [CrossRef]
- Tempest, R.; Guarnerio, S.; Maani, R.; Cooper, J.; Peake, N. The Biological and Biomechanical Role of Transglutaminase-2 in the Tumour Microenvironment. Cancers 2021, 13, 2788. [Google Scholar] [CrossRef]
- Schuppan, D.; Maki, M.; Lundin, K.E.A.; Isola, J.; Friesing-Sosnik, T.; Taavela, J.; Popp, A.; Koskenpato, J.; Langhorst, J.; Hovde, O.; et al. A Randomized Trial of a Transglutaminase 2 Inhibitor for Celiac Disease. N. Engl. J. Med. 2021, 385, 35–45. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Total (n = 137) | IgAN (n = 100) | DMN (n = 27) | CTL (n = 10) | p Value | |||
|---|---|---|---|---|---|---|---|---|
| Group | IgAN vs. CTL | DMN vs. CTL | IgAN vs. DMN | |||||
| Age (year) | 46.88 ± 15.73 | 46.09 ± 15.47 | 53.44 ± 14.06 | 42.00 ± 5.99 | 0.041 | 0.495 | 0.053 | 0.019 |
| Sex (male, %) | 81 (59.12) | 58 (58) | 18 (66.66) | 5 (50) | 0.597 | |||
| BMI (kg/m2) | 24.12 ± 3.56 | 24.11 ± 3.60 | 24.51 ± 3.232 | 23.16 ± 4.11 | 0.593 | 0.436 | 0.301 | 0.597 |
| CKD stage 1, 2 (%) | 34 (24.82) | 27 (27) | 7 (25.93) | |||||
| CKD stage 3 (%) | 63 (45.99) | 50 (50) | 13 (48.15) | |||||
| CKD stage 4, 5 (%) | 30 (21.90) | 23 (23) | 7 (25.93) | |||||
| Blood hemoglobin (g/dL) | 12.26 ± 2.05 | 12.41 ± 1.98 | 11.19 ± 1.95 | 13.69 ± 1.86 | 0.005 | 0.052 | 0.001 | 0.005 |
| Serum albumin (g/dL) | 3.84 ± 0.54 | 3.85 ± 0.56 | 3.69 ± 0.50 | 4.07 ± 0.39 | 0.145 | 0.236 | 0.039 | 0.176 |
| Creatinine (mg/dL) | 1.76 ± 1.53 | 1.79 ± 1.59 | 2.04 ± 1.50 | 0.78 ± 0.19 | 0.169 | 0.048 | 0.013 | 0.467 |
| eGFR (CKD-EPI, mL/min/1.73 m2) | 56.44 ± 33.79 | 54.08 ± 32.90 | 47.50 ± 26.42 | 104.2 ± 22.26 | <0.001 | <0.001 | <0.001 | 0.340 |
| Urine protein/creatinine ratio | 1.78 ± 2.06 | 1.55 ± 2.01 | 3.06 ± 2.08 | 0.60 ± 0.28 | 0.002 | 0.140 | 0.027 | 0.001 |
| Plasma TG2 level (pg/mL) | 1990 ± 357.8 | 2030 ± 347.8 | 1963 ± 334.0 | 1665 ± 381.3 | 0.021 | 0.002 | 0.001 | 0.371 |
| Variable | Total (n = 94) | IgAN (n = 56) | DMN (n = 21) | CTL (n = 17) | p-Value | |||
|---|---|---|---|---|---|---|---|---|
| Group | IgAN vs. CTL | DMN vs. CTL | IgAN vs. DMN | |||||
| Age (year) | 44.95 ± 17.90 | 45.84 ± 15.16 | 47.24 ± 11.26 | 39.18 ± 17.90 | 0.205 | 0.133 | 0.099 | 0.702 |
| Sex (male, %) | 56 (56.57) | 36 (64.29) | 14 (66.67) | 6 (35.29) | 0.077 | |||
| BMI (kg/m2) | 23.75 ± 3.72 | 24.28 ± 3.12 | 25.04 ± 3.97 | 25.22 ± 4.00 | 0.694 | 0.514 | 0.927 | 0.476 |
| CKD stage 1, 2 (%) | 20 (35.71) | 5 (23.81) | ||||||
| CKD stage 3 (%) | 20 (35.71) | 9 (42.86) | ||||||
| CKD stage 4, 5 (%) | 16 (28.57) | 7 (33.33) | ||||||
| Blood hemoglobin (g/dL) | 12.26 ± 2.10 | 12.24 ± 2.00 | 11.44 ± 2.37 | 13.34 ± 1.68 | 0.019 | 0.043 | 0.008 | 0.141 |
| Serum albumin (g/dL) | 3.81 ± 0.59 | 3.85 ± 0.49 | 3.42 ± 0.77 | 4.14 ± 0.34 | <0.001 | 0.031 | 0.001 | 0.004 |
| Creatinine (mg/dL) | 1.56 ± 1.00 | 1.66 ± 0.99 | 1.95 ± 1.10 | 0.75 ± 0.17 | <0.001 | <0.001 | <0.001 | 0.259 |
| eGFR (CKD-EPI, mL/min/1.73 m2) | 64.26 ± 37.75 | 56.49 ± 33.39 | 50.51 ± 36.81 | 106.8 ± 19.39 | <0.001 | <0.001 | <0.001 | 0.498 |
| Urine protein/creatinine ratio | 2.42 ± 3.42 | 1.75 ± 2.17 | 5.77 ± 5.01 | 0.47 ± 0.30 | <0.001 | 0.019 | <0.001 | <0.001 |
| TG2 positive area (%) | 8.24 ± 5.61 | 10.26 ± 5.25 | 6.76 ± 5.72 | 3.39 ± 2.24 | <0.001 | <0.001 | 0.028 | 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, J.-J.; Choi, Y.; Kim, K.-H.; Seo, A.; Kwon, S.; Kim, Y.-C.; Kim, D.-K.; Kim, Y.-S.; Yang, S.-H. Inhibiting Transglutaminase 2 Mediates Kidney Fibrosis via Anti-Apoptosis. Biomedicines 2022, 10, 1345. https://doi.org/10.3390/biomedicines10061345
Moon J-J, Choi Y, Kim K-H, Seo A, Kwon S, Kim Y-C, Kim D-K, Kim Y-S, Yang S-H. Inhibiting Transglutaminase 2 Mediates Kidney Fibrosis via Anti-Apoptosis. Biomedicines. 2022; 10(6):1345. https://doi.org/10.3390/biomedicines10061345
Chicago/Turabian StyleMoon, Jong-Joo, Yejin Choi, Kyu-Hyeon Kim, Areum Seo, Soie Kwon, Yong-Chul Kim, Dong-Ki Kim, Yon-Su Kim, and Seung-Hee Yang. 2022. "Inhibiting Transglutaminase 2 Mediates Kidney Fibrosis via Anti-Apoptosis" Biomedicines 10, no. 6: 1345. https://doi.org/10.3390/biomedicines10061345
APA StyleMoon, J. -J., Choi, Y., Kim, K. -H., Seo, A., Kwon, S., Kim, Y. -C., Kim, D. -K., Kim, Y. -S., & Yang, S. -H. (2022). Inhibiting Transglutaminase 2 Mediates Kidney Fibrosis via Anti-Apoptosis. Biomedicines, 10(6), 1345. https://doi.org/10.3390/biomedicines10061345