
High Level Expression and Purification of Cecropin-like Antimicrobial Peptides in Escherichia coli

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Construction of the Expression Plasmid
2.3. Expression of His6-Intein-KR12AGPWR6
2.4. Purification of His6-Intein-KR12AGPWR6
2.5. Intein Self-Cleavage
2.6. Purification of rKR12AGPWR6 by RP-HPLC
2.7. SDS-PAGE Analysis
2.8. Antimicrobial Activity Assays
2.9. Circular Dichroism Spectroscopy
2.10. Nuclear Magnetic Resonance Spectroscopy (NMR)
3. Results and Discussion
3.1. Construction of the Recombinant Plasmid
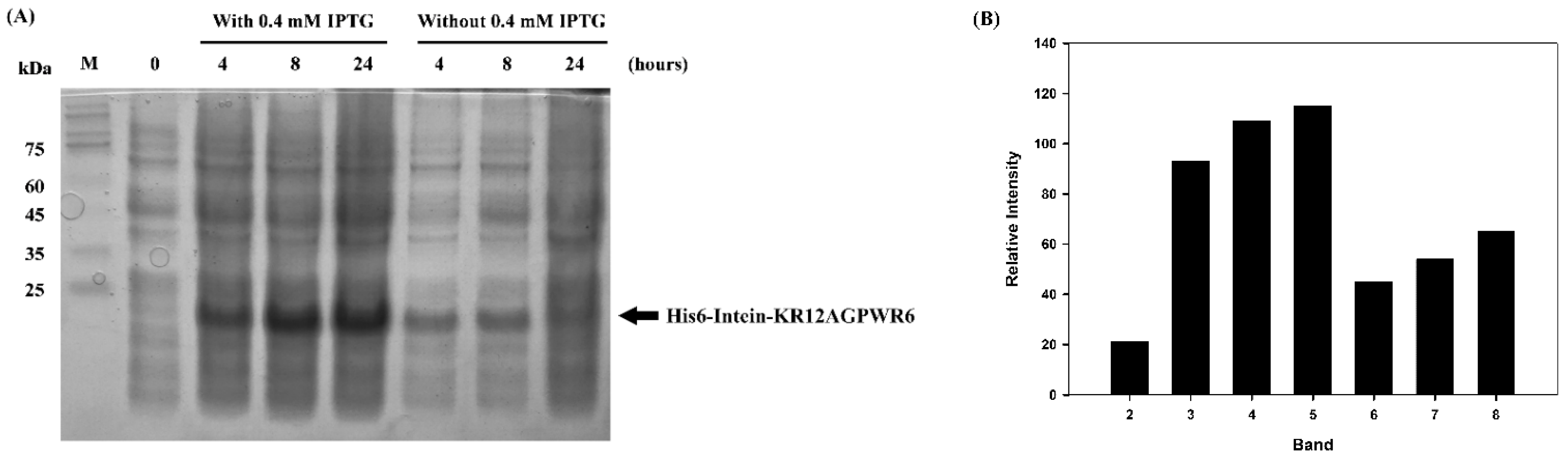
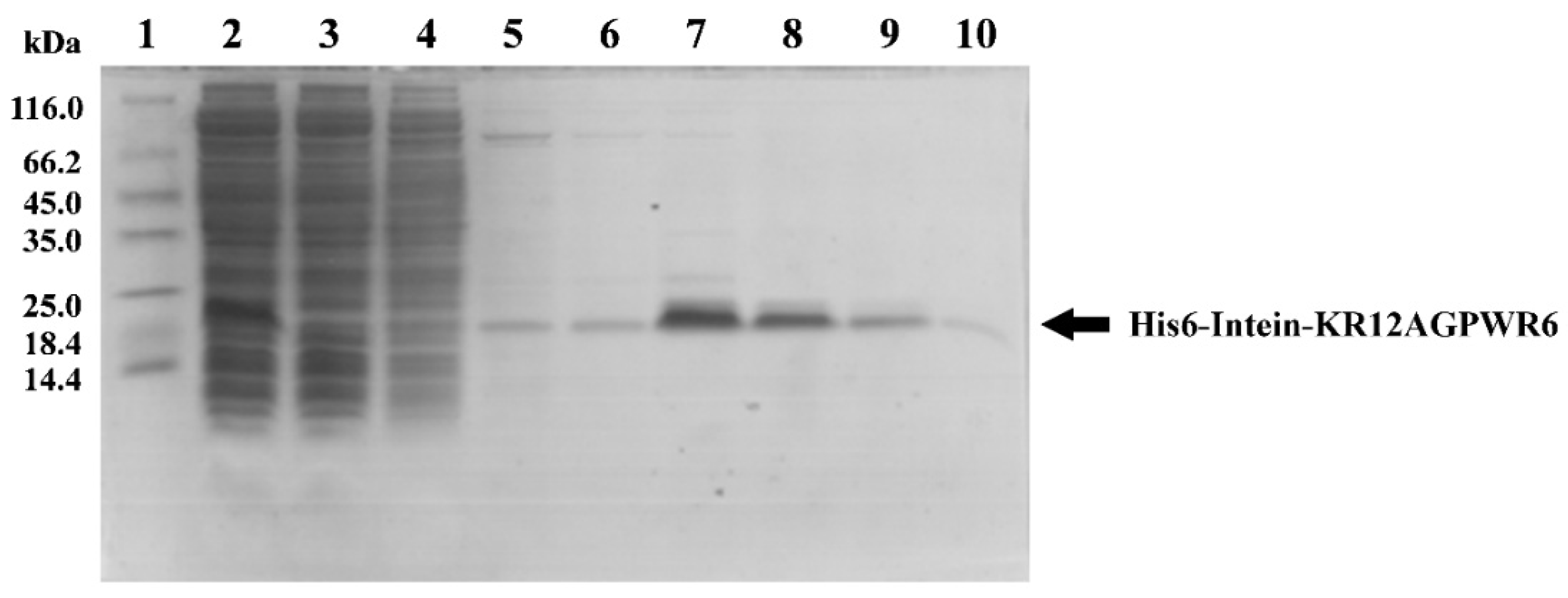
3.2. Expression, Extraction and Purification of Recombinant His6-Intein-KR12AGPWR6
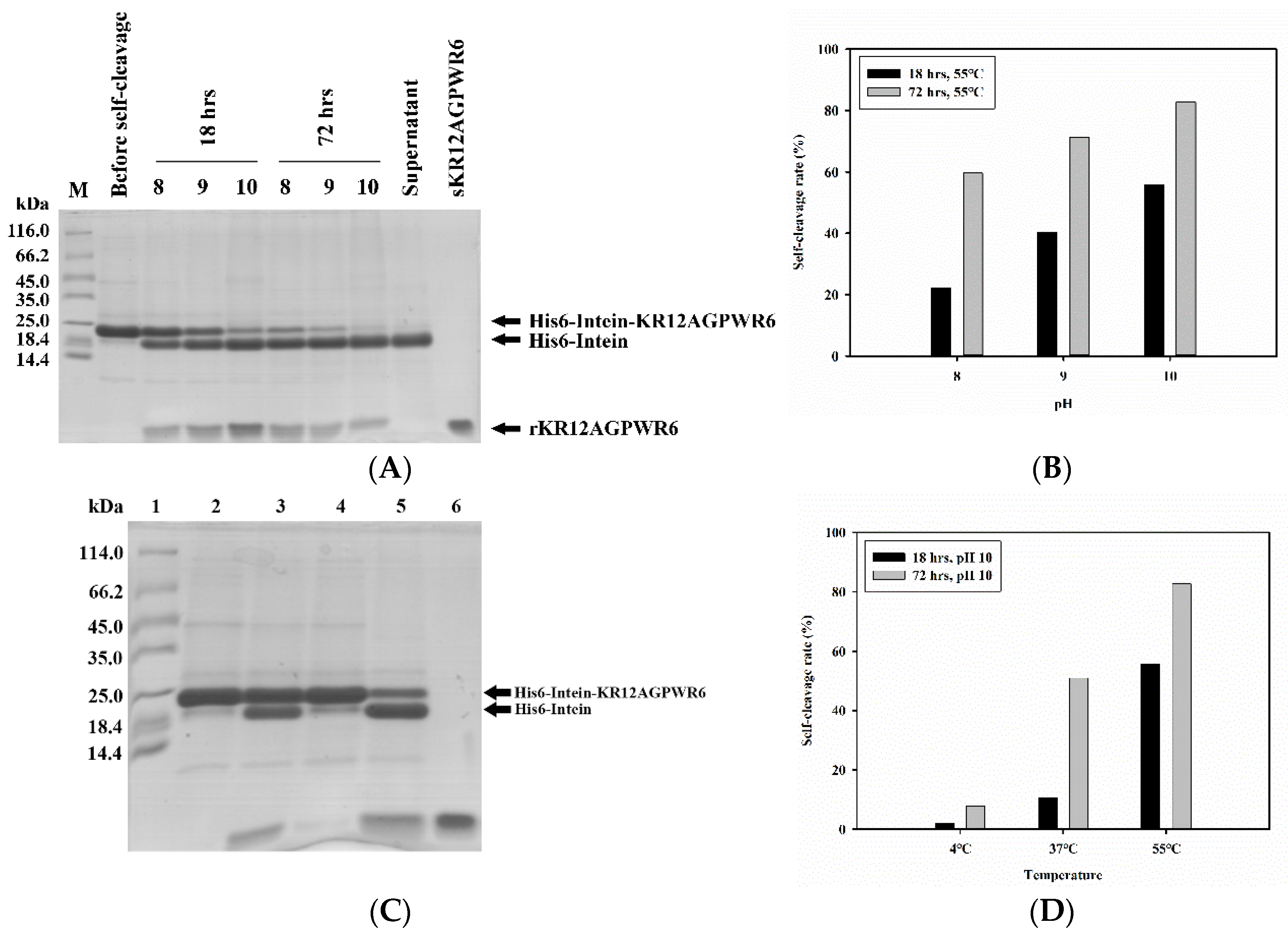
3.3. Optimization of Intein’s Self-Cleavage
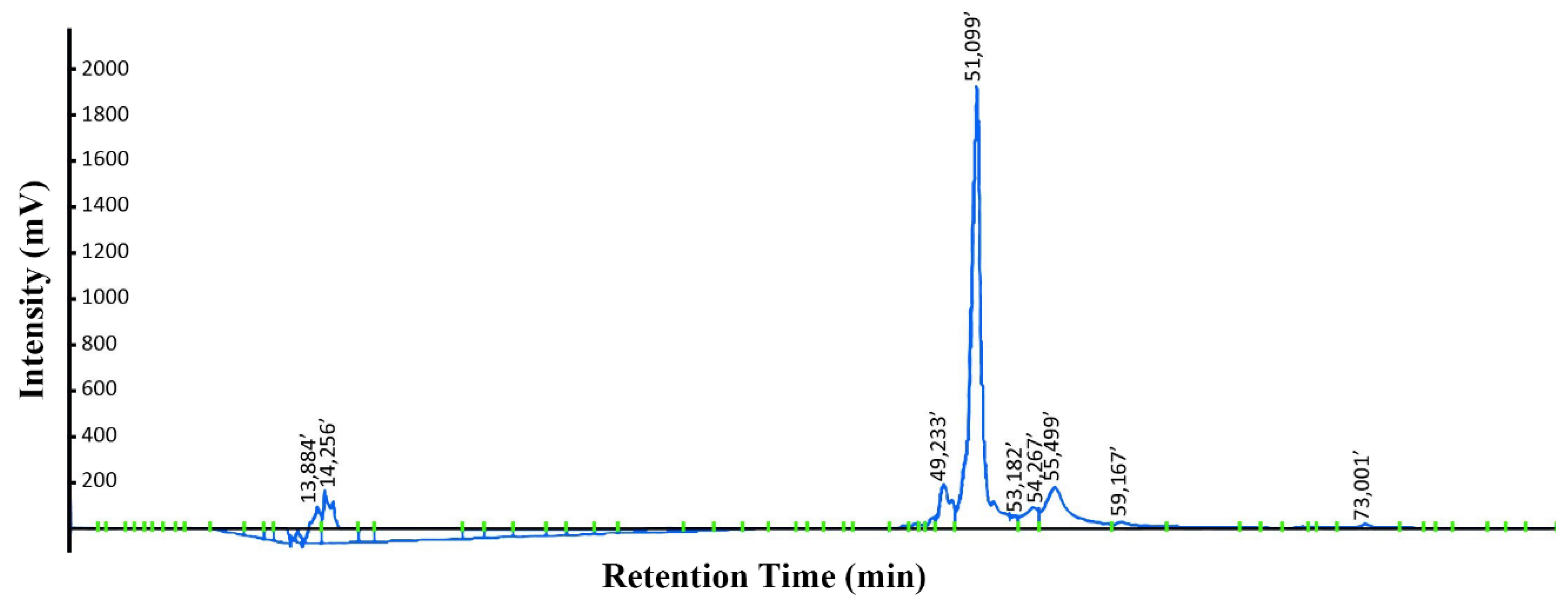
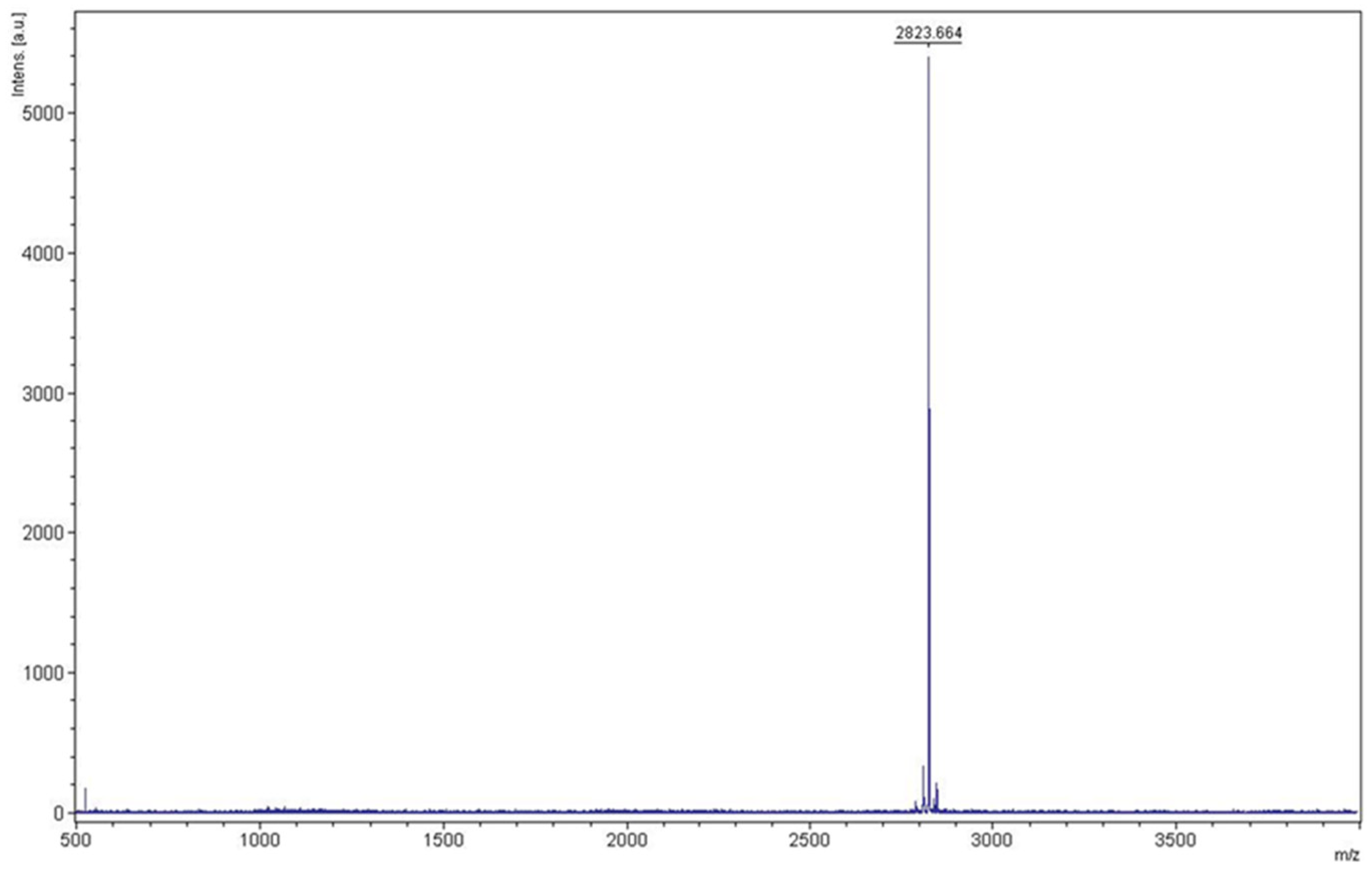
3.4. Purification of KR12AGPWR6
3.5. Antimicrobial Activity
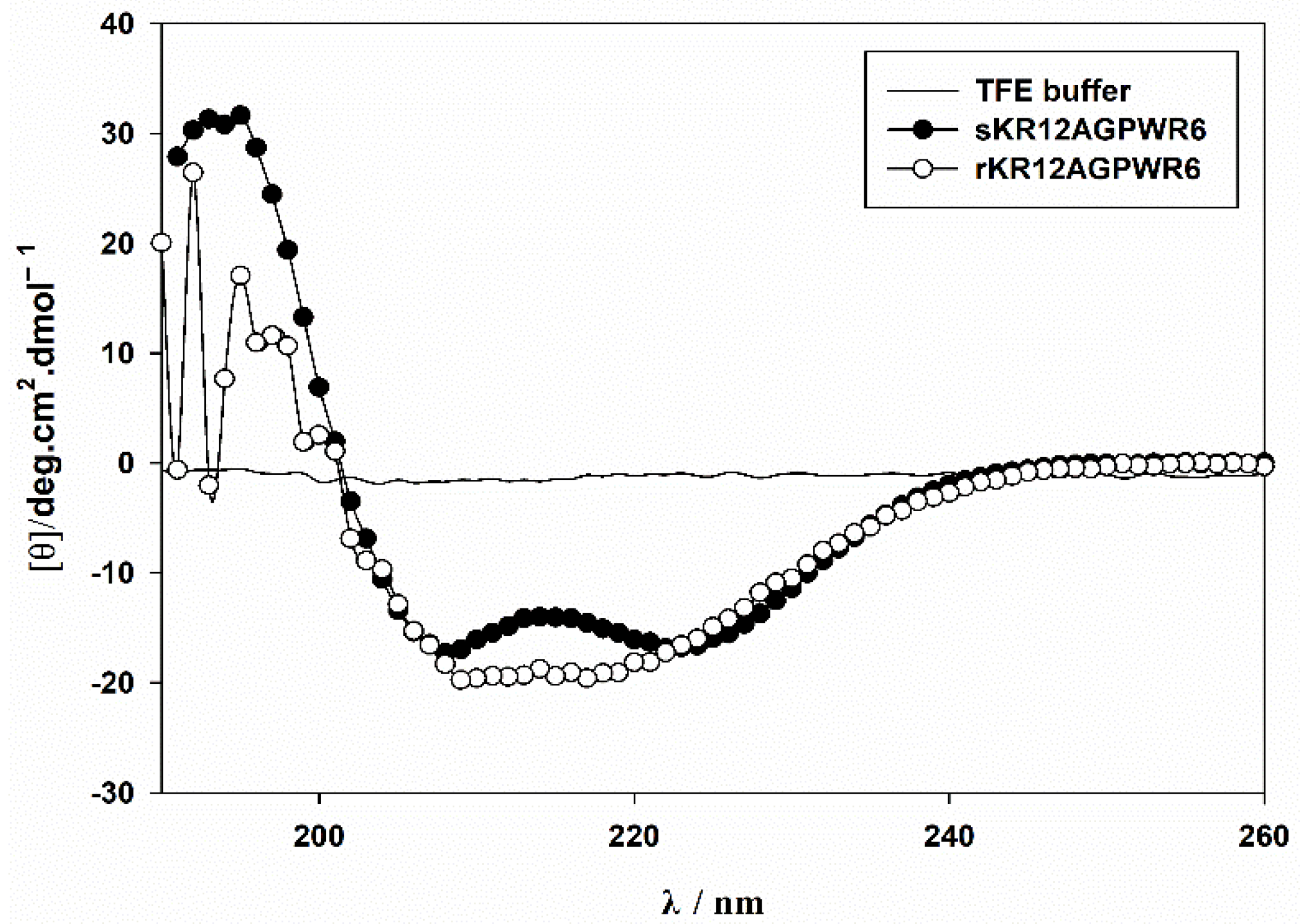
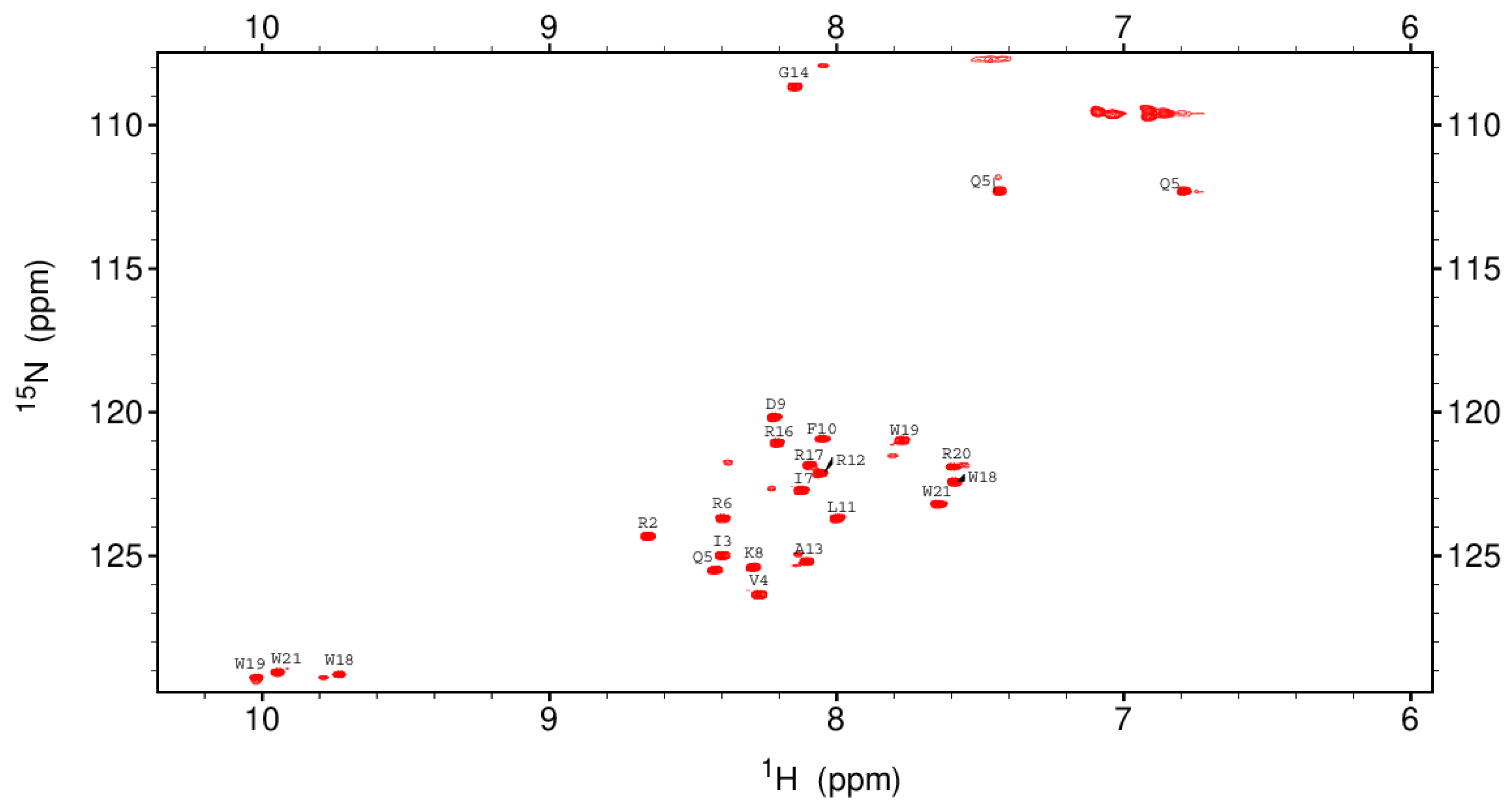
3.6. Characterization of the Recombinant rKR12AGPWR6 by CD and NMR
3.7. Design of New Cecropin-like Peptides
3.8. Expression and Purification of Cecropin-like AMPs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sinha, S.; Harioudh, M.K.; Dewangan, R.P.; Ng, W.J.; Ghosh, J.K.; Bhattacharjya, S. Cell-selective pore forming antimicrobial peptides of the prodomain of human furin: A conserved aromatic/cationic sequence mapping, membrane disruption, and atomic-resolution structure and dynamics. ACS Omega 2018, 3, 14650–14664. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, A.; Wu, X.; Yeow, E.K.L.; Bhattacharjya, S. Designed Heme-Cage beta-Sheet Miniproteins. Angew. Chem. Int. Ed. Engl. 2017, 56, 5904–5908. [Google Scholar] [CrossRef]
- Dwivedi, R.; Aggarwal, P.; Bhavesh, N.S.; Kaur, K.J. Design of therapeutically improved analogue of the antimicrobial peptide, indolicidin, using a glycosylation strategy. Amino Acids 2019, 51, 1443–1460. [Google Scholar] [CrossRef]
- Andreu, D.; Rivas, L. Animal antimicrobial peptides: An overview. Biopolymers 1998, 47, 415–433. [Google Scholar] [CrossRef]
- Bhattacharjya, S. NMR Structures and Interactions of Antimicrobial Peptides with Lipopolysaccharide: Connecting Structures to Functions. Curr. Top. Med. Chem. 2015, 16, 4–15. [Google Scholar] [CrossRef]
- Mohanram, H.; Bhattacharjya, S. ‘Lollipop’-shaped helical structure of a hybrid antimicrobial peptide of temporin B-lipopolysaccharide binding motif and mapping cationic residues in antibacterial activity. Biochim. Biophys. Acta 2016, 1860, 1362–1372. [Google Scholar] [CrossRef]
- Bechinger, B.; Gorr, S.U. Antimicrobial Peptides: Mechanisms of Action and Resistance. J. Dent. Res. 2016, 96, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.-K.; Kim, C.; Seo, C.H.; Park, Y. The therapeutic applications of antimicrobial peptides (AMPs): A patent review. J. Microbiol. 2016, 55, 1–12. [Google Scholar] [CrossRef]
- Draper, L.A.; Cotter, P.D.; Hill, C.; Ross, R.P. The two peptide lantibiotic lacticin 3147 acts synergistically with polymyxin to inhibit Gram negative bacteria. BMC Microbiol. 2013, 13, 212. [Google Scholar] [CrossRef] [Green Version]
- Naghmouchi, K.; Baah, J.; Hober, D.; Jouy, E.; Rubrecht, C.; Sané, F.; Drider, D. Synergistic Effect between Colistin and Bacteriocins in Controlling Gram-Negative Pathogens and Their Potential To Reduce Antibiotic Toxicity in Mammalian Epithelial Cells. Antimicrob. Agents Chemother. 2013, 57, 2719–2725. [Google Scholar] [CrossRef] [Green Version]
- Etayash, H.; Alford, M.; Akhoundsadegh, N.; Drayton, M.; Straus, S.K.; Hancock, R.E.W. Multifunctional Antibiotic-Host Defense Peptide Conjugate Kills Bacteria, Eradicates Biofilms, and Modulates the Innate Immune Response. J. Med. Chem. 2021, 64, 16854–16863. [Google Scholar] [CrossRef]
- Li, W.; O’Brien-Simpson, N.M.; Holden, J.A.; Otvos, L.; Reynolds, E.C.; Separovic, F.; Hossain, M.A.; Wade, J.D. Covalent conjugation of cationic antimicrobial peptides with a b-lactam antibiotic core. Pept. Sci. 2018, 110, e24059. [Google Scholar] [CrossRef]
- Bhattacharjya, S. De novo Designed Lipopolysaccharide Binding Peptides: Structure Based Development of Antiendotoxic and Antimicrobial Drugs. Curr. Med. Chem. 2010, 17, 3080–3093. [Google Scholar] [CrossRef]
- Lin, L.; Chi, J.; Yan, Y.; Luo, R.; Feng, X.; Zheng, Y.; Xian, D.; Li, X.; Quan, G.; Liu, D.; et al. Membrane-disruptive peptides/peptidomimetics-based therapeutics: Promising systems to combat bacteria and cancer in the drug-resistant era. Acta Pharm. Sin. B 2021, 11, 2609–2644. [Google Scholar] [CrossRef]
- Guryanova, S.V.; Ovchinnikova, T.V. Immunomodulatory and Allergenic Properties of Antimicrobial Peptides. Int. J. Mol. Sci. 2022, 23, 2499. [Google Scholar] [CrossRef]
- Wu, J.-M.; Jan, P.-S.; Yu, H.-C.; Haung, H.-Y.; Fang, H.-J.; Chang, Y.-I.; Cheng, J.-W.; Chen, H.M. Structure and function of a custom anticancer peptide, CB1a. Peptides 2009, 30, 839–848. [Google Scholar] [CrossRef]
- Moore, A.J.; Beazley, W.D.; Bibby, M.C.; Devine, D.A. Antimicrobial activity of cecropins. J. Antimicrob. Chemother. 1996, 37, 1077–1089. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Lee, E.; Shin, S.; Jeong, K.W.; Lee, J.Y.; Bae, S.Y.; Kim, S.H.; Lee, J.; Kim, S.R.; Lee, D.G.; et al. Structure and Function of Papiliocin with Antimicrobial and Anti-inflammatory Activities Isolated from the Swallowtail Butterfly, Papilio xuthus. J. Biol. Chem. 2011, 286, 41296–41311. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Shin, A.; Kim, Y. Anti-inflammatory activities of cecropin a and its mechanism of action. Arch. Insect Biochem. Physiol. 2015, 88, 31–44. [Google Scholar] [CrossRef]
- Chu, H.-L.; Chih, Y.-H.; Peng, K.-L.; Wu, C.-L.; Yu, H.-Y.; Cheng, D.; Chou, Y.-T.; Cheng, J.-W. Antimicrobial Peptides with Enhanced Salt Resistance and Antiendotoxin Properties. Int. J. Mol. Sci. 2020, 21, 6810. [Google Scholar] [CrossRef]
- Lee, Y.Z.; Sue, S.C. Salt-sensitive intein for large-scale polypeptide production. Methods Enzym. 2019, 621, 111–130. [Google Scholar]
- Gunasekaran, P.; Kim, E.Y.; Lee, J.; Ryu, E.K.; Shin, S.Y.; Bang, J.K. Synthesis of Fmoc-Triazine Amino Acids and Its Application in the Synthesis of Short Antibacterial Peptidomimetics. Int. J. Mol. Sci. 2020, 21, 3602. [Google Scholar] [CrossRef]
- Andersson, L.; Blomberg, L.; Flegel, M.; Lepsa, L.; Nilsson, B.; Verlander, M. Large-scale synthesis of peptides. Biopolymers 2000, 55, 227–250. [Google Scholar] [CrossRef]
- Li, Y. Recombinant production of antimicrobial peptides in Escherichia coli: A review. Protein Expr. Purif. 2011, 80, 260–267. [Google Scholar] [CrossRef]
- Cheng, K.-T.; Wu, C.-L.; Yip, B.-S.; Yu, H.-Y.; Cheng, H.-T.; Chih, Y.-H.; Cheng, J.-W. High Level Expression and Purification of the Clinically Active Antimicrobial Peptide P-113 in Escherichia coli. Molecules 2018, 23, 800. [Google Scholar] [CrossRef] [Green Version]
- Sang, M.; Wei, H.; Zhang, J.; Wei, Z.; Wu, X.; Chen, Y.; Zhuge, Q. Expression and characterization of the antimicrobial peptide ABP-dHC-cecropin A in the methylotrophic yeast Pichia pastoris. Protein Expr. Purif. 2017, 140, 44–51. [Google Scholar] [CrossRef]
- Dhakal, S.; Sapkota, K.; Huang, F.; Rangachari, V. Cloning, expression and purification of the low-complexity region of RanBP9 protein. Protein Expr. Purif. 2020, 172, 105630. [Google Scholar] [CrossRef]
- Wang, Q.; Yin, M.; Yuan, C.; Liu, X.; Hu, Z.; Zou, Z.; Wang, M. Identification of a Conserved Prophenoloxidase Activation Pathway in Cotton Bollworm Helicoverpa armigera. Front. Immunol. 2020, 11, 785. [Google Scholar] [CrossRef]
- Gibbs, G.M.; Davidson, B.E.; Hillier, A.J. Novel Expression System for Large-Scale Production and Purification of Recombinant Class IIa Bacteriocins and Its Application to Piscicolin 126. Appl. Environ. Microbiol. 2004, 70, 3292–3297. [Google Scholar] [CrossRef] [Green Version]
- Holbrook, S.R.; Cheong, C.; Tinoco, I., Jr.; Kim, S.-H. Crystal structure of an RNA double helix incorporating a track of non-Watson–Crick base pairs. Nature 1991, 353, 579–581. [Google Scholar] [CrossRef]
- Li, Q.; Chen, A.S.; Gayen, S.; Kang, C. Expression and purification of the p75 neurotrophin receptor transmembrane domain using a ketosteroid isomerase tag. Microb. Cell Fact. 2012, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Lai, Z.-T.; Lu, M.-K.; Gong, X.-G.; Xie, Y. Expression and Hydroxylamine Cleavage of Thymosin Alpha 1 Concatemer. J. Biomed. Biotechnol. 2008, 2008, 736060. [Google Scholar] [CrossRef] [Green Version]
- Perler, F.B.; Davis, E.O.; Dean, G.E.; Gimble, F.S.; Jack, W.E.; Neff, N.; Noren, C.J.; Thorner, J.; Belfort, M. Protein splicing elements: Inteins and exteins—A definition of terms and recommended nomenclature. Nucleic Acids Res. 1994, 22, 1125–1127. [Google Scholar] [CrossRef]
- Singleton, S.F.; A Simonette, R.; Sharma, N.C.; I Roca, A. Intein-mediated affinity-fusion purification of the Escherichia coli RecA protein. Protein Expr. Purif. 2002, 26, 476–488. [Google Scholar] [CrossRef]
- Guo, C.; Li, Z.; Shi, Y.; Xu, M.; Wise, J.; Trommer, W.E.; Yuan, J. Intein-mediated fusion expression, high efficient refolding, and one-step purification of gelonin toxin. Protein Expr. Purif. 2004, 37, 361–367. [Google Scholar] [CrossRef]
- Nudelman, H.; Lee, Y.Z.; Hung, Y.L.; Kolusheva, S.; Upcher, A.; Chen, Y.C.; Chen, J.Y.; Sue, S.C.; Zarivach, R. Understanding the Biomineralization Role of Magnetite-Interacting Components (MICs) From Magnetotactic Bacteria. Front. Microbiol. 2018, 9, 2480. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). In Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2015.
- Chen, H.M.; Wang, W.; Smith, D.; Chan, S.C. Effects of the anti-bacterial peptide cecropin B and its analogs, cecropins B-1 and B-2, on liposomes, bacteria, and cancer cells. Biochim. Biophys. Acta 1997, 1336, 171–179. [Google Scholar] [CrossRef]
- Wu, C.; Geng, X.; Wan, S.; Hou, H.; Yu, F.; Jia, B.; Wang, L. Cecropin-P17, an analog of Cecropin B, inhibits human hepatocellular carcinoma cell HepG-2 proliferation via regulation of ROS, Caspase, Bax, and Bcl-2. J. Pept. Sci. Off. Publ. Eur. Pept. Soc. 2015, 21, 661–668. [Google Scholar] [CrossRef]
- Fox, M.A.; Thwaite, J.E.; Ulaeto, D.O.; Atkins, T.P.; Atkins, H.S. Design and characterization of novel hybrid antimicrobial peptides based on cecropin A, LL-37 and magainin II. Peptides 2012, 33, 197–205. [Google Scholar] [CrossRef]
- Bhunia, A.; Ramamoorthy, A.; Bhattacharjya, S. Helical hairpin structure of a potent antimicrobial peptide MSI-594 in lipopolysaccharide micelles by NMR spectroscopy. Chemistry 2009, 15, 2036–2040. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Herrera, A.I.; Bommineni, Y.R.; Soulages, J.L.; Prakash, O.; Zhang, G. The Central Kink Region of Fowlicidin-2, an a-Helical Host Defense Peptide, Is Critically Involved in Bacterial Killing and Endotoxin Neutralizationon. J. Innate Immun. 2009, 1, 268–280. [Google Scholar] [CrossRef]
- Tack, B.F.; Sawai, M.V.; Kearney, W.R.; Robertson, A.D.; Sherman, M.A.; Wang, W.; Hong, T.; Boo, L.M.; Wu, H.; Waring, A.J.; et al. SMAP-29 has two LPS-binding sites and a central hinge. JBIC J. Biol. Inorg. Chem. 2002, 269, 1181–1189. [Google Scholar] [CrossRef]
- Bhunia, A.; Domadia, P.N.; Torres, J.; Hallock, K.J.; Ramamoorthy, A.; Bhattacharjya, S. NMR structure of pardaxin, a pore-forming antimicrobial peptide in lipopolysaccharide micelles: Mechanism of outer membrane permeabilization. J. Biol. Chem. 2010, 285, 3883–3895. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Kim, J.K.; Jeon, D.; Jeong, K.W.; Shin, A.; Kim, Y. Functional Roles of Aromatic Residues and Helices of Papilioncin in its Antimicrobial and Anti-inflammatory Activities. Sci. Rep. 2015, 5, 12048. [Google Scholar] [CrossRef] [Green Version]
- Pasupuleti, M.; Chalupka, A.; Mörgelin, M.; Schmidtchen, A.; Malmsten, M. Tryptophan end-tagging of antimicrobial peptides for increased potency against Pseudomonas aeruginosa. Biochim. Biophys. Acta 2009, 1790, 800–808. [Google Scholar] [CrossRef]
- Pasupuleti, M.; Schmidtchen, A.; Chalupka, A.; Ringstad, L.; Malmsten, M. End-Tagging of Ultra-Short Antimicrobial Peptides by W/F Stretches to Facilitate Bacterial Killing. PLoS ONE 2009, 4, e5285. [Google Scholar] [CrossRef] [Green Version]
- Schmidtchen, A.; Pasupuleti, M.; Mörgelin, M.; Davoudi, M.; Alenfall, J.; Chalupka, A.; Malmsten, M. Boosting Antimicrobial Peptides by Hydrophobic Oligopeptide End Tags. J. Biol. Chem. 2009, 284, 17584–17594. [Google Scholar] [CrossRef] [Green Version]
- Duong, D.T.; Singh, S.; Bagheri, M.; Verma, N.K.; Schmidtchen, A.; Malmsten, M. Pronounced peptide selectivity for melanoma through tryptophan end-tagging. Sci. Rep. 2016, 6, 24952. [Google Scholar] [CrossRef] [Green Version]
- Ember, J.A.; Johansen, N.L.; Hugli, T.E. Designing synthetic superagonists of C3a anaphylatoxin. Biochemistry 1991, 30, 3603–3612. [Google Scholar] [CrossRef]
- Chu, H.-L.; Yu, H.-Y.; Yip, B.-S.; Chih, Y.-H.; Liang, C.-W.; Cheng, H.-T.; Cheng, J.-W. Boosting Salt Resistance of Short Antimicrobial Peptides. Antimicrob. Agents Chemother. 2013, 57, 4050–4052. [Google Scholar] [CrossRef] [Green Version]
- Chih, Y.-H.; Lin, Y.-S.; Yip, B.-S.; Wei, H.-J.; Chu, H.-L.; Yu, H.-Y.; Cheng, H.-T.; Chou, Y.-T.; Cheng, J.-W. Ultrashort Antimicrobial Peptides with Antiendotoxin Properties. Antimicrob. Agents Chemother. 2015, 59, 5052–5056. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.Y.; Chen, Y.A.; Yip, B.S.; Wang, S.Y.; Wei, H.J.; Chih, Y.H.; Chen, K.H.; Cheng, J.W. Role of b-naphthylalanine end-tags in the enhancement of antiendotoxin activities: Solution structure of the antimicrobial peptide S1-Nal-Nal in complex with lipopolysaccharide. Biochim. Biophys. Acta 2017, 1859, 1114–1123. [Google Scholar] [CrossRef]
- Li, Y. Split-inteins and their bioapplications. Biotechnol. Lett. 2015, 37, 2121–2137. [Google Scholar] [CrossRef]
- Wood, D.W.; Camarero, J.A. Intein Applications: From Protein Purification and Labeling to Metabolic Control Methods. J. Biol. Chem. 2014, 289, 14512–14519. [Google Scholar] [CrossRef] [Green Version]
- Li, Y. Carrier proteins for fusion expression of antimicrobial peptides in Escherichia coli. Biotechnol. Appl. Biochem. 2009, 54, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ganjave, S.D.; Dodia, H.; Sunder, A.V.; Madhu, S.; Wangikar, P.P. High cell density cultivation of E. coli in shake flasks for the production of recombinant proteins. Biotechnol. Rep. 2021, 33, e00694. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | Minimum Inhibitory Concentration (μg/mL) | |
|---|---|---|
| sKR12AGPWR6 | rKR12AGPWR6 | |
| S. aureus ATCC 25923 | 2 | 2 |
| E. coli ATCC 25922 | 2 | 4 |
| P. aeruginosa ATCC 27853 | 2 | 4 |
| A. baumannii BCRC 14B0100 | 1 | 1 |
| Magainin-AGP-WR6 | GIGKFLHSAKKFGKAFVGEIMNS-AGP-RRWWRW |
| KR12-AGP-Cecropin P1 | KRIVQRIKDFLR-AGP-IAIAIQGGPR |
| Cecropin P1-AGP-WR6 | SWLSKTAKKLENSAKKRISE-AGP-RRWWRW |
| KR12-AGP-Cecropin A | KRIVQRIKDFLR-AGP-AVAVVGQATQIAK |
| CecropinA-AGP-WR6 | KWKLFKKIEKVGQNIRDGIIK-AGP-RRWWRW |
| Melittin-AGP-WR6 | GIGAVLKVLTTGLPALISWIKRKRQQ-AGP-RRWWRW |
| Melittin-AGP-Cecropin P1 | GIGAVLKVLTTGLPALISWIKRKRQQ-AGP-IAIAIQGGPR |
| Bacterial Strains | Minimum Inhibitory Concentration (μg/mL) |
|---|---|
| S. aureus ATCC 29213 | 16 |
| E. coli ATCC 25922 | 8 |
| P. aeruginosa ATCC 27853 | 8 |
| A. baumannii BCRC 14B0100 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.-L.; Chih, Y.-H.; Hsieh, H.-Y.; Peng, K.-L.; Lee, Y.-Z.; Yip, B.-S.; Sue, S.-C.; Cheng, J.-W. High Level Expression and Purification of Cecropin-like Antimicrobial Peptides in Escherichia coli. Biomedicines 2022, 10, 1351. https://doi.org/10.3390/biomedicines10061351
Wu C-L, Chih Y-H, Hsieh H-Y, Peng K-L, Lee Y-Z, Yip B-S, Sue S-C, Cheng J-W. High Level Expression and Purification of Cecropin-like Antimicrobial Peptides in Escherichia coli. Biomedicines. 2022; 10(6):1351. https://doi.org/10.3390/biomedicines10061351
Chicago/Turabian StyleWu, Chih-Lung, Ya-Han Chih, Hsin-Ying Hsieh, Kuang-Li Peng, Yi-Zong Lee, Bak-Sau Yip, Shih-Che Sue, and Jya-Wei Cheng. 2022. "High Level Expression and Purification of Cecropin-like Antimicrobial Peptides in Escherichia coli" Biomedicines 10, no. 6: 1351. https://doi.org/10.3390/biomedicines10061351
APA StyleWu, C. -L., Chih, Y. -H., Hsieh, H. -Y., Peng, K. -L., Lee, Y. -Z., Yip, B. -S., Sue, S. -C., & Cheng, J. -W. (2022). High Level Expression and Purification of Cecropin-like Antimicrobial Peptides in Escherichia coli. Biomedicines, 10(6), 1351. https://doi.org/10.3390/biomedicines10061351