
Bromelain Protects Critically Perfused Musculocutaneous Flap Tissue from Necrosis

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Bromelain Treatment
2.3. Anesthesia
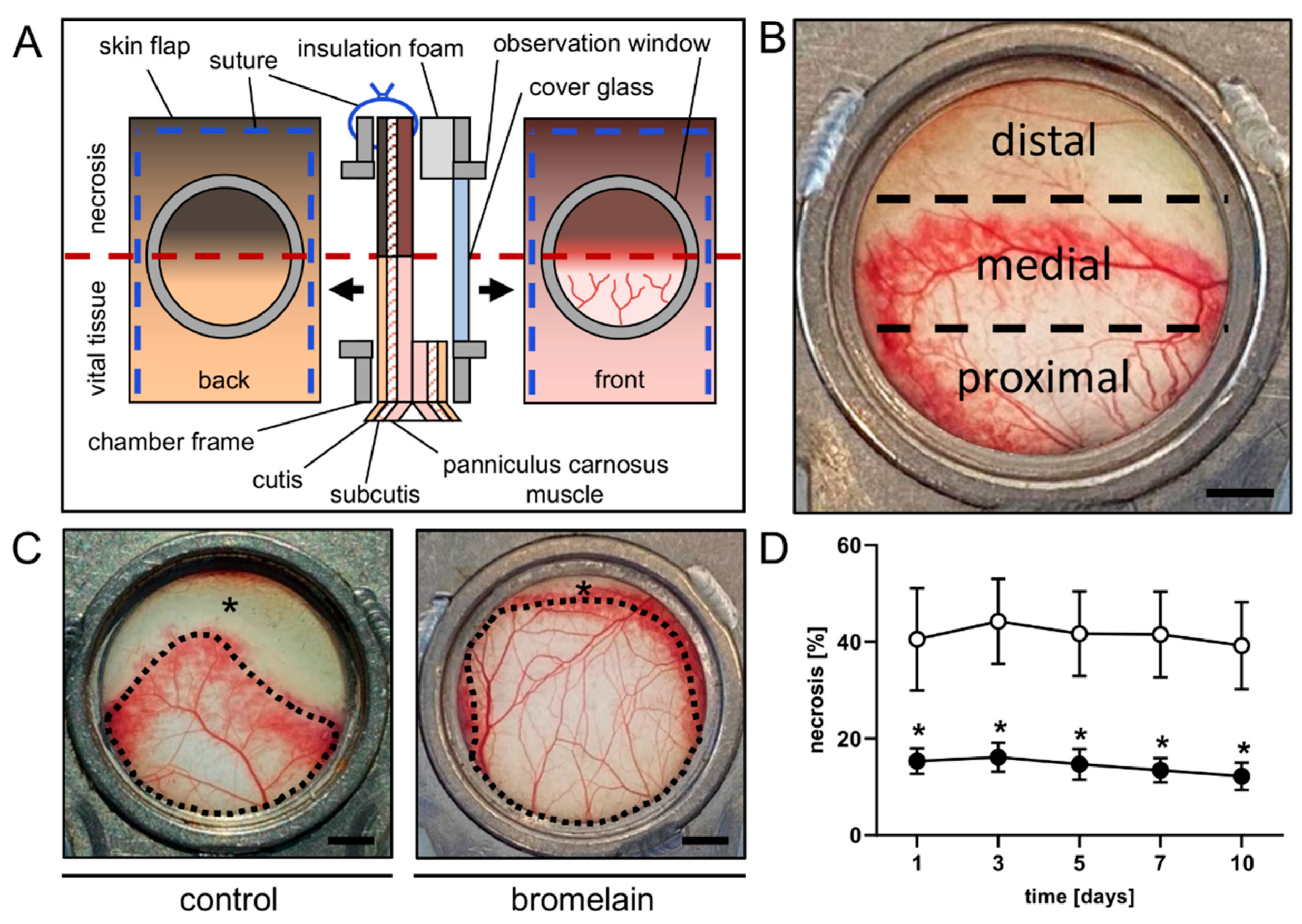
2.4. Dorsal Skinfold Chamber-Flap Model
2.5. Intravital Fluorescence Microscopy
2.6. Histology and Immunohistochemistry
2.7. Statistical Analysis
3. Results
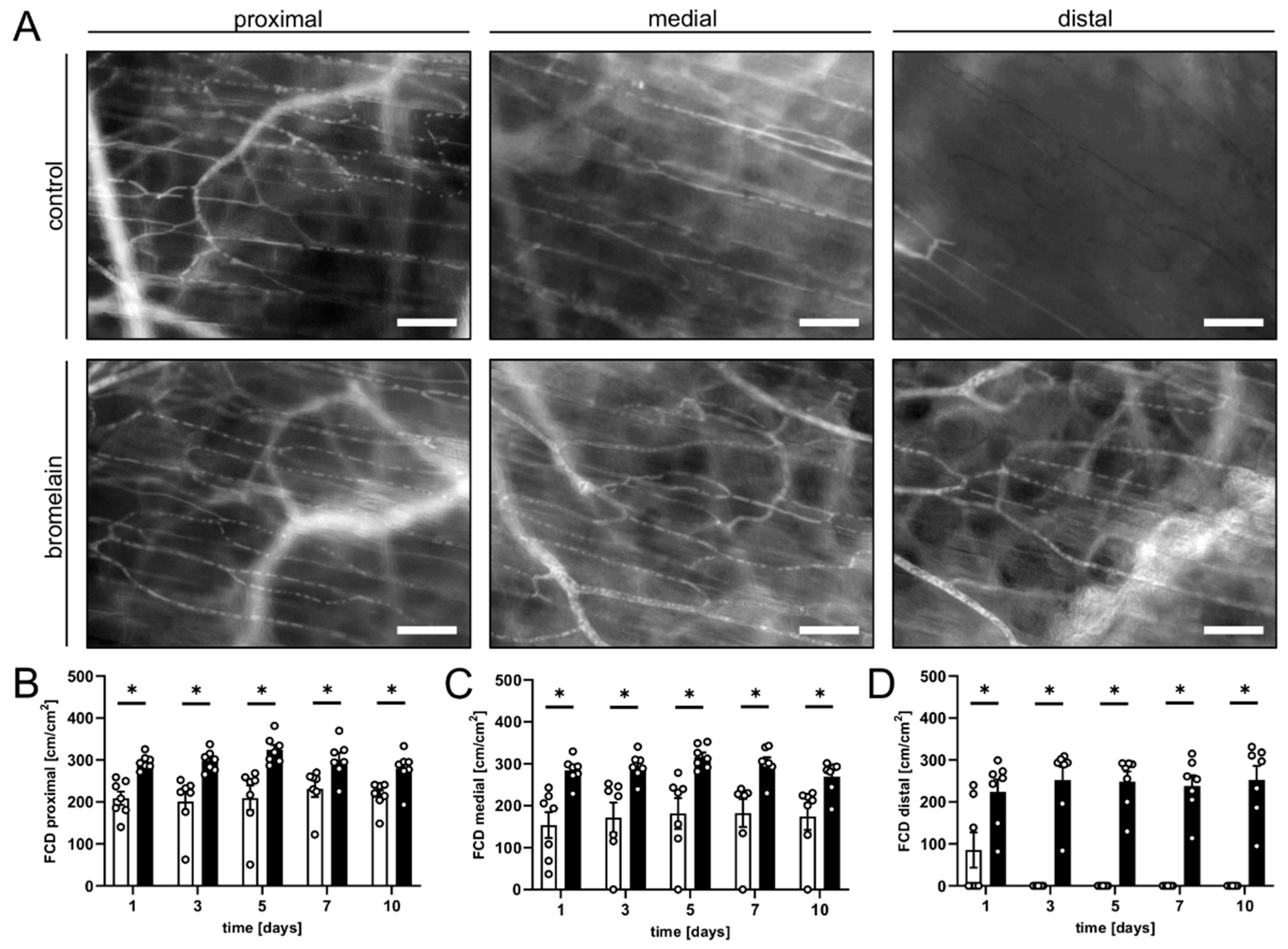
3.1. Intravital Fluorescence Microscopy
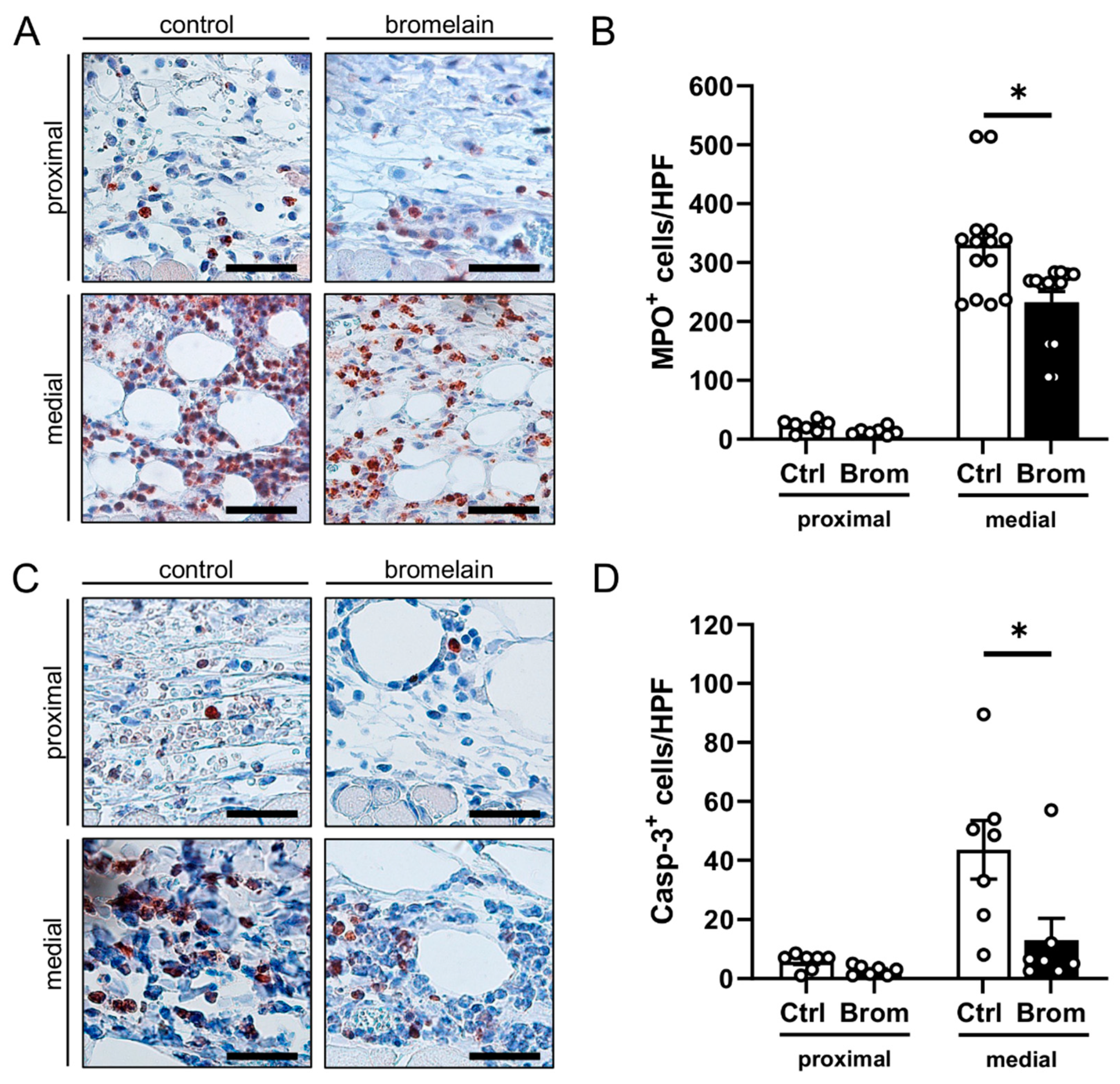
3.2. Histological and Immunohistochemical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, A.M. Natural Products as a Resource for New Drugs. Pharm. Res. 1996, 13, 1133–1141. [Google Scholar] [CrossRef]
- Cheuka, P.M.; Mayoka, G.; Mutai, P.; Chibale, K. The Role of Natural Products in Drug Discovery and Development against Neglected Tropical Diseases. Molecules 2017, 22, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, J.M.; Schneider, B.; Beuth, H.J. Therapeutic Use, Efficiency and Safety of the Proteolytic Pineapple Enzyme Bromelain-POS® in Children with Acute Sinusitis in Germany. In Vivo 2005, 19, 417–421. [Google Scholar] [PubMed]
- Taub, S.J. The Use of Ananase in Sinusitis. A Study of 60 Patients. Eye Ear Nose Throat Mon. 1966, 45, 96. [Google Scholar]
- Chakraborty, A.J.; Mitra, S.; Tallei, T.E.; Tareq, A.M.; Nainu, F.; Cicia, D.; Dhama, K.; Emran, T.B.; Simal-Gandara, J.; Capasso, R. Bromelain a Potential Bioactive Compound: A Comprehensive Overview from a Pharmacological Perspective. Life 2021, 11, 317. [Google Scholar] [CrossRef] [PubMed]
- Pavan, R.; Jain, S.; Shraddha; Kumar, A. Properties and Therapeutic Application of Bromelain: A Review. Biotechnol. Res. Int. 2012, 2012, 976203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, H.R. Bromelain: Biochemistry, Pharmacology and Medical Use. Cell. Mol. Life Sci. 2001, 58, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Taussig, S.J.; Batkin, S. Bromelain, the Enzyme Complex of Pineapple (Ananas Comosus) and Its Clinical Application. An Update. J. Ethnopharmacol. 1988, 22, 191–203. [Google Scholar] [CrossRef]
- Kumakura, S.; Yamashita, M.; Tsurufuji, S. Effect of Bromelain on Kaolin-Induced Inflammation in Rats. Eur. J. Pharmacol. 1988, 150, 295–301. [Google Scholar] [CrossRef]
- Azarkan, M.; González, M.M.; Esposito, R.C.; Errasti, M.E. Stem Bromelain Proteolytic Machinery: Study of the Effects of Its Components on Fibrin (Ogen) and Blood Coagulation. Protein Pept. Lett. 2020, 27, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Onken, J.E.; Greer, P.K.; Calingaert, B.; Hale, L.P. Bromelain Treatment Decreases Secretion of Pro-Inflammatory Cytokines and Chemokines by Colon Biopsies in Vitro. Clin. Immunol. 2008, 126, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzhugh, D.J.; Shan, S.; Dewhirst, M.W.; Hale, L.P. Bromelain Treatment Decreases Neutrophil Migration to Sites of Inflammation. Clin. Immunol. 2008, 128, 66–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahde, R.; Palmes, D.; Minin, E.; Stratmann, U.; Diller, R.; Haier, J.; Spiegel, H.U. Bromelain Ameliorates Hepatic Microcirculation After Warm Ischemia. J. Surg. Res. 2007, 139, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Deramo, P.; Rose, J. Flaps, Muscle and Musculocutaneous; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Saint-Cyr, M.; Wong, C.; Schaverien, M.; Mojallal, A.; Rohrich, R.J. The Perforasome Theory: Vascular Anatomy and Clinical Implications. Plast. Reconstr. Surg. 2009, 124, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Memarzadeh, K.; Sheikh, R.; Blohmé, J.; Torbrand, C.; Malmsjö, M. Perfusion and Oxygenation of Random Advancement Skin Flaps Depend More on the Length and Thickness of the Flap Than on the Width to Length Ratio. Eplasty 2016, 16, e12. [Google Scholar]
- Pestana, I.A.; Jones, V.M.; Velazquez, C. Breast Reconstruction and Nipple-Sparing Mastectomy: Technical Modifications and Their Outcomes Over Time at an Academic Breast Center. Ann. Plast. Surg. 2021, 86, e81. [Google Scholar] [CrossRef]
- Juhasz, B.; Thirunavukkarasu, M.; Pant, R.; Zhan, L.; Penumathsa, S.V.; Secor, E.R.; Srivastava, S.; Raychaudhuri, U.; Menon, V.P.; Otani, H.; et al. Bromelain Induces Cardioprotection against Ischemia-Reperfusion Injury through Akt/FOXO Pathway in Rat Myocardium. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1365–H1370. [Google Scholar] [CrossRef] [Green Version]
- Harder, Y.; Amon, M.; Erni, D.; Menger, M.D. Evolution of Ischemic Tissue Injury in a Random Pattern Flap: A New Mouse Model Using Intravital Microscopy. J. Surg. Res. 2004, 121, 197–205. [Google Scholar] [CrossRef]
- De Vriese, A.S.; Verbeuren, T.J.; Vallez, M.O.; Lameire, N.H.; De Buyzere, M.; Vanhoutte, P.M. Off-Line Analysis of Red Blood Cell Velocity in Renal Arterioles. J. Vasc. Res. 2000, 37, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.; Wayland, H. On-Line Volume Flow Rate and Velocity Profile Measurement for Blood in Microvessels. Microvasc. Res. 1974, 7, 131–143. [Google Scholar] [CrossRef]
- Menger, M.D.; Lehr, H.A. Scope and Perspectives of Intravital Microscopy-Bridge over from In Vitro to In Vivo. Immunol. Today 1993, 14, 519–522. [Google Scholar] [CrossRef]
- Hikisz, P.; Bernasinska-Slomczewska, J. Beneficial Properties of Bromelain. Nutrients 2021, 13, 4313. [Google Scholar] [CrossRef] [PubMed]
- Che, C.T.; Zhang, H. Plant Natural Products for Human Health. Int. J. Mol. Sci. 2019, 20, 830. [Google Scholar] [CrossRef] [Green Version]
- Neumayer, C.; Fügl, A.; Nanobashvili, J.; Blumer, R.; Punz, A.; Gruber, H.; Polterauer, P.; Huk, I. Combined Enzymatic and Antioxidative Treatment Reduces Ischemia-Reperfusion Injury in Rabbit Skeletal Muscle. J. Surg. Res. 2006, 133, 150–158. [Google Scholar] [CrossRef]
- Shoba, E.; Lakra, R.; Kiran, M.S.; Korrapati, P.S. Fabrication of Core-Shell Nanofibers for Controlled Delivery of Bromelain and Salvianolic Acid B for Skin Regeneration in Wound Therapeutics. Biomed. Mater. 2017, 12, 035005. [Google Scholar] [CrossRef]
- Shoba, E.; Lakra, R.; Kiran, M.S.; Korrapati, P.S. 3 D Nano Bilayered Spatially and Functionally Graded Scaffold Impregnated Bromelain Conjugated Magnesium Doped Hydroxyapatite Nanoparticle for Periodontal Regeneration. J. Mech. Behav. Biomed. Mater. 2020, 109, 103822. [Google Scholar] [CrossRef]
- Rathnavelu, V.; Alitheen, N.B.; Sohila, S.; Kanagesan, S.; Ramesh, R. Potential Role of Bromelain in Clinical and Therapeutic Applications (Review). Biomed. Rep. 2016, 5, 283–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsen, M.; Hovden, A.O.; Vogelsang, P.; Tysnes, B.B.; Appel, S. Bromelain Treatment Leads to Maturation of Monocyte-Derived Dendritic Cells but Cannot Replace PGE 2 in a Cocktail of IL-1β, IL-6, TNF-α and PGE 2. Scand. J. Immunol. 2011, 74, 135–143. [Google Scholar] [CrossRef]
- Metzig, C.; Grabowska, E.; Eckert, K.; Rehse, K.; Maurer, H.R. Bromelain Proteases Reduce Human Platelet Aggregation in Vitro, Adhesion to Bovine Endothelial Cells and Thrombus Formation in Rat Vessels in Vivo. In Vivo 1999, 13, 7–12. [Google Scholar]
- Gläser, D.; Hilberg, T. The Influence of Bromelain on Platelet Count and Platelet Activity in Vitro. Platelets 2006, 17, 37–41. [Google Scholar] [CrossRef]
- Fitzal, F.; DeLano, F.A.; Young, C.; Schmid-Schönbein, G.W. Early Capillary No-Reflow during Low-Flow Reperfusion after Hind Limb Ischemia in the Rat. Ann. Plast. Surg. 2002, 49, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Menger, M.D.; Laschke, M.W.; Amon, M.; Schramm, R.; Thorlacius, H.; Rücker, M.; Vollmar, B. Experimental Models to Study Microcirculatory Dysfunction in Muscle Ischemia-Reperfusion and Osteomyocutaneous Flap Transfer. Langenbecks Arch Surg. 2003, 388, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Errasti, M.E.; Prospitti, A.; Viana, C.A.; Gonzalez, M.M.; Ramos, M.v.; Rotelli, A.E.; Caffini, N.O. Effects on Fibrinogen, Fibrin, and Blood Coagulation of Proteolytic Extracts from Fruits of Pseudananas Macrodontes, Bromelia Balansae, and B. Hieronymi (Bromeliaceae) in Comparison with Bromelain. Blood Coagul. Fibrinolysis 2016, 27, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Romano, B.; Fasolino, I.; Pagano, E.; Capasso, R.; Pace, S.; de Rosa, G.; Milic, N.; Orlando, P.; Izzo, A.A.; Borrelli, F. The Chemopreventive Action of Bromelain, from Pineapple Stem (Ananas Comosus L.), on Colon Carcinogenesis Is Related to Antiproliferative and Proapoptotic Effects. Mol. Nutr. Food Res. 2014, 58, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Insuan, O.; Janchai, P.; Thongchuai, B.; Chaiwongsa, R.; Khamchun, S.; Saoin, S.; Insuan, W.; Pothacharoen, P.; Apiwatanapiwat, W.; Boondaeng, A.; et al. Anti-Inflammatory Effect of Pineapple Rhizome Bromelain through Downregulation of the NF-ΚB-and MAPKs-Signaling Pathways in Lipopolysaccharide (LPS)-Stimulated RAW264.7 Cells. Curr. Issues Mol. Biol. 2021, 43, 93–106. [Google Scholar] [CrossRef]
- Kasemsuk, T.; Vivithanaporn, P.; Unchern, S. Anti-Inflammatory Effects of Bromelain in Lps-Induced Human U937 Macrophages. Chiang Mai J. Sci. 2018, 45, 299–307. [Google Scholar]
- Hale, L.P.; Greer, P.K.; Sempowski, G.D. Bromelain Treatment Alters Leukocyte Expression of Cell Surface Molecules Involved in Cellular Adhesion and Activation. Clin. Immunol. 2002, 104, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Harder, Y.; Amon, M.; Wettstein, R.; Rücker, M.; Schramm, R.; Menger, M.D. Gender-Specific Ischemic Tissue Tolerance in Critically Perfused Skin. Langenbeck’s Arch. Surg. 2010, 395, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harder, Y.; Amon, M.; Georgi, M.; Scheuer, C.; Schramm, R.; Rücker, M.; Pittet, B.; Erni, D.; Menger, M.D. Aging Is Associated with an Increased Susceptibility to Ischaemic Necrosis Due to Microvascular Perfusion Failure but Not a Reduction in Ischaemic Tolerance. Clin. Sci. 2007, 112, 429–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Lippo, L.; Labella, R.; Tan, S.L.; Marsden, B.D.; Dustin, M.L.; Ramasamy, S.K.; Kusumbe, A.P. Decreased Blood Vessel Density and Endothelial Cell Subset Dynamics during Ageing of the Endocrine System. EMBO J. 2021, 40, e105242. [Google Scholar] [CrossRef]
- Chen, J.; Sivan, U.; Tan, S.L.; Lippo, L.; de Angelis, J.; Labella, R.; Singh, A.; Chatzis, A.; Cheuk, S.; Medhghalchi, M.; et al. High-Resolution 3D Imaging Uncovers Organ-Specific Vascular Control of Tissue Aging. Sci. Adv. 2021, 7, eabd7819. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volumetric Blood Flow [pL/s] | d1 | d3 | d5 | d7 | d10 | |
|---|---|---|---|---|---|---|
| Arterioles | ||||||
| proximal | Ctrl | 458 ± 93 | 598 ± 158 | 814 ± 152 | 1145 ± 230 | 1272 ± 160 |
| Brom | 790 ± 86 * | 807 ± 127 | 960 ± 146 | 1459 ± 196 | 1634 ± 160 | |
| medial | Ctrl | 437 ± 96 | 603 ± 117 | 733 ± 90 | 1013 ± 152 | 1085 ± 159 |
| Brom | 687 ± 107 | 1036 ± 91 * | 1302 ± 165 * | 1574 ± 120 * | 1550 ± 188 | |
| distal | Ctrl | 485 ± 149 | - | - | - | - |
| Brom | 538 ± 116 | 962 ± 237 | 1141 ± 276 | 1203 ± 261 | 1540 ± 248 | |
| Capillaries | ||||||
| proximal | Ctrl | 3 ± 0 | 3 ± 0 | 4 ± 1 | 5 ± 1 | 6 ± 1 |
| Brom | 4 ± 0 | 4 ± 0 * | 4 ± 1 | 6 ± 1 | 7 ± 1 | |
| medial | Ctrl | 3 ± 0 | 3 ± 0 | 4 ± 1 | 5 ± 1 | 6 ± 1 |
| Brom | 4 ± 0 | 4 ± 0 * | 4 ± 1 | 6 ± 1 | 7 ± 1 | |
| distal | Ctrl | 2 ± 1 | - | - | - | - |
| Brom | 2 ± 0 | 4 ± 0 | 5 ± 1 | 6 ± 1 | 6 ± 1 | |
| Venules | ||||||
| proximal | Ctrl | 383 ± 47 | 502 ± 89 | 667 ± 100 | 776 ± 137 | 1233 ± 517 |
| Brom | 773 ± 87 * | 1167 ± 131 * | 1570 ± 410 * | 1894 ± 536 * | 1876 ± 397 | |
| medial | Ctrl | 333 ± 47 | 452 ± 74 | 691 ± 140 | 897 ± 200 | 779 ± 266 |
| Brom | 833 ± 158 * | 1265 ± 146 * | 1766 ± 389 * | 2009 ± 362 * | 1650 ± 433 | |
| distal | Ctrl | 146 ± 41 | - | - | - | - |
| Brom | 472 ± 171 | 1012 ± 308 | 1387 ± 326 | 1268 ± 301 | 1859 ± 337 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weinzierl, A.; Harder, Y.; Schmauss, D.; Menger, M.D.; Laschke, M.W. Bromelain Protects Critically Perfused Musculocutaneous Flap Tissue from Necrosis. Biomedicines 2022, 10, 1449. https://doi.org/10.3390/biomedicines10061449
Weinzierl A, Harder Y, Schmauss D, Menger MD, Laschke MW. Bromelain Protects Critically Perfused Musculocutaneous Flap Tissue from Necrosis. Biomedicines. 2022; 10(6):1449. https://doi.org/10.3390/biomedicines10061449
Chicago/Turabian StyleWeinzierl, Andrea, Yves Harder, Daniel Schmauss, Michael D. Menger, and Matthias W. Laschke. 2022. "Bromelain Protects Critically Perfused Musculocutaneous Flap Tissue from Necrosis" Biomedicines 10, no. 6: 1449. https://doi.org/10.3390/biomedicines10061449
APA StyleWeinzierl, A., Harder, Y., Schmauss, D., Menger, M. D., & Laschke, M. W. (2022). Bromelain Protects Critically Perfused Musculocutaneous Flap Tissue from Necrosis. Biomedicines, 10(6), 1449. https://doi.org/10.3390/biomedicines10061449