

Cardioprotective Role for Paraoxonase-1 in Chronic Kidney Disease

 , , ,
, , ,  , , and
, , and 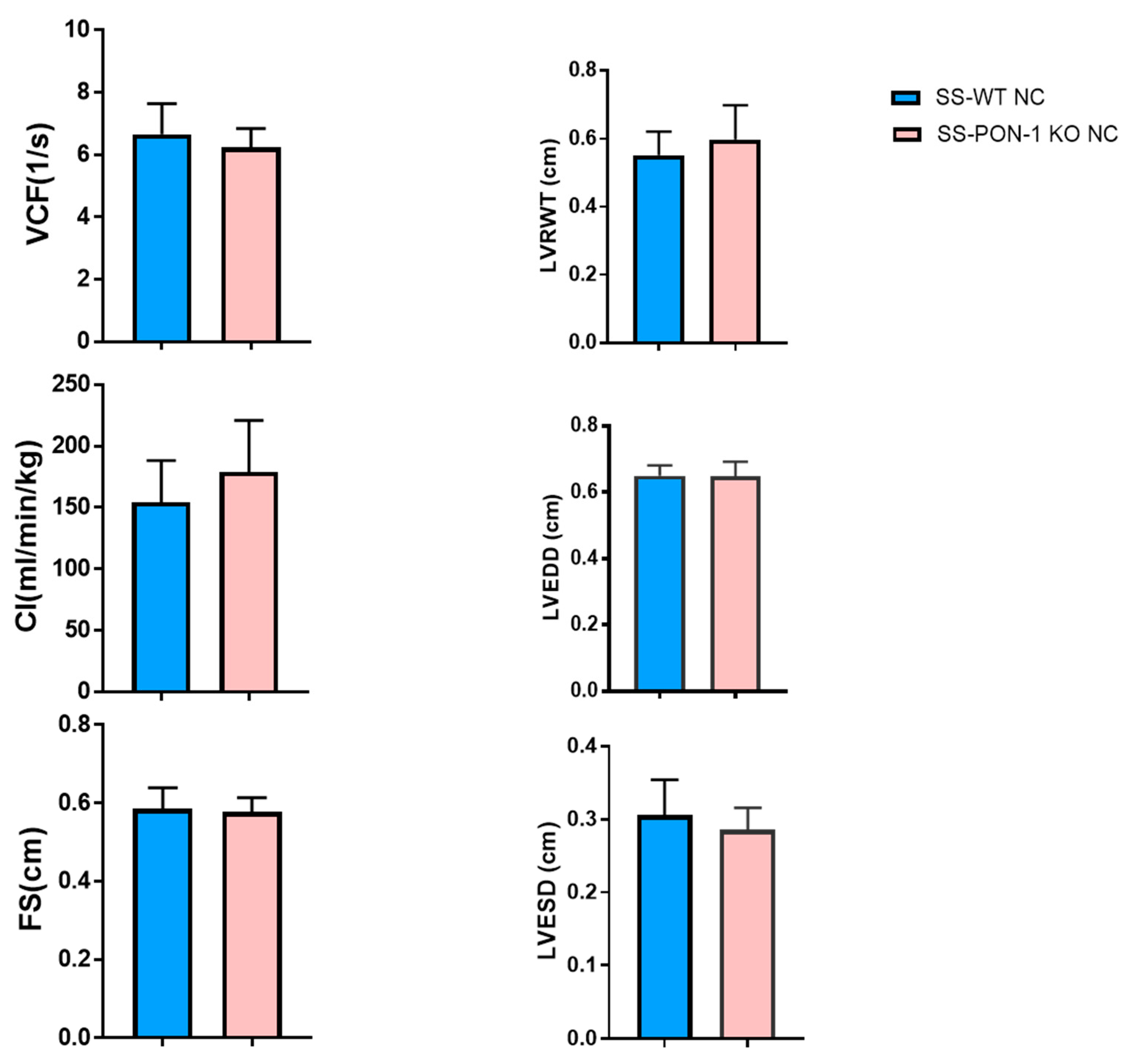{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Echocardiography
2.3. Histology
2.4. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) and RNA Isolation
2.5. Statistical Analysis
3. Results
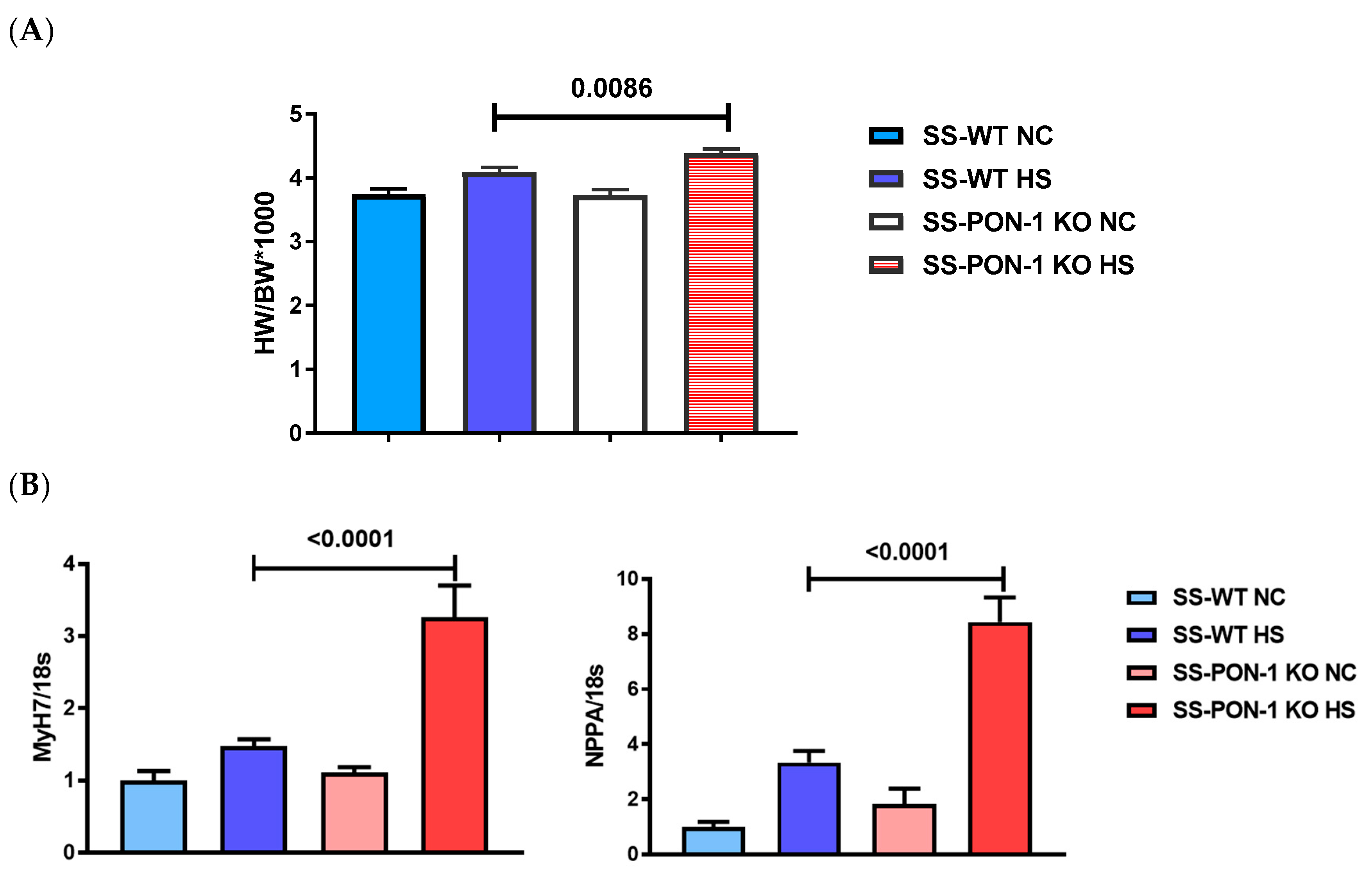
3.1. PON-1 KO Rats on High-Salt Diet Demonstrate LV Cardiac Hypertrophy
3.2. SS-PON-1 KO Rats on High-Salt Diet Demonstrate Significantly Increase in Cardiac Fibrosis Compared to Age-Matched SS-WT Rats
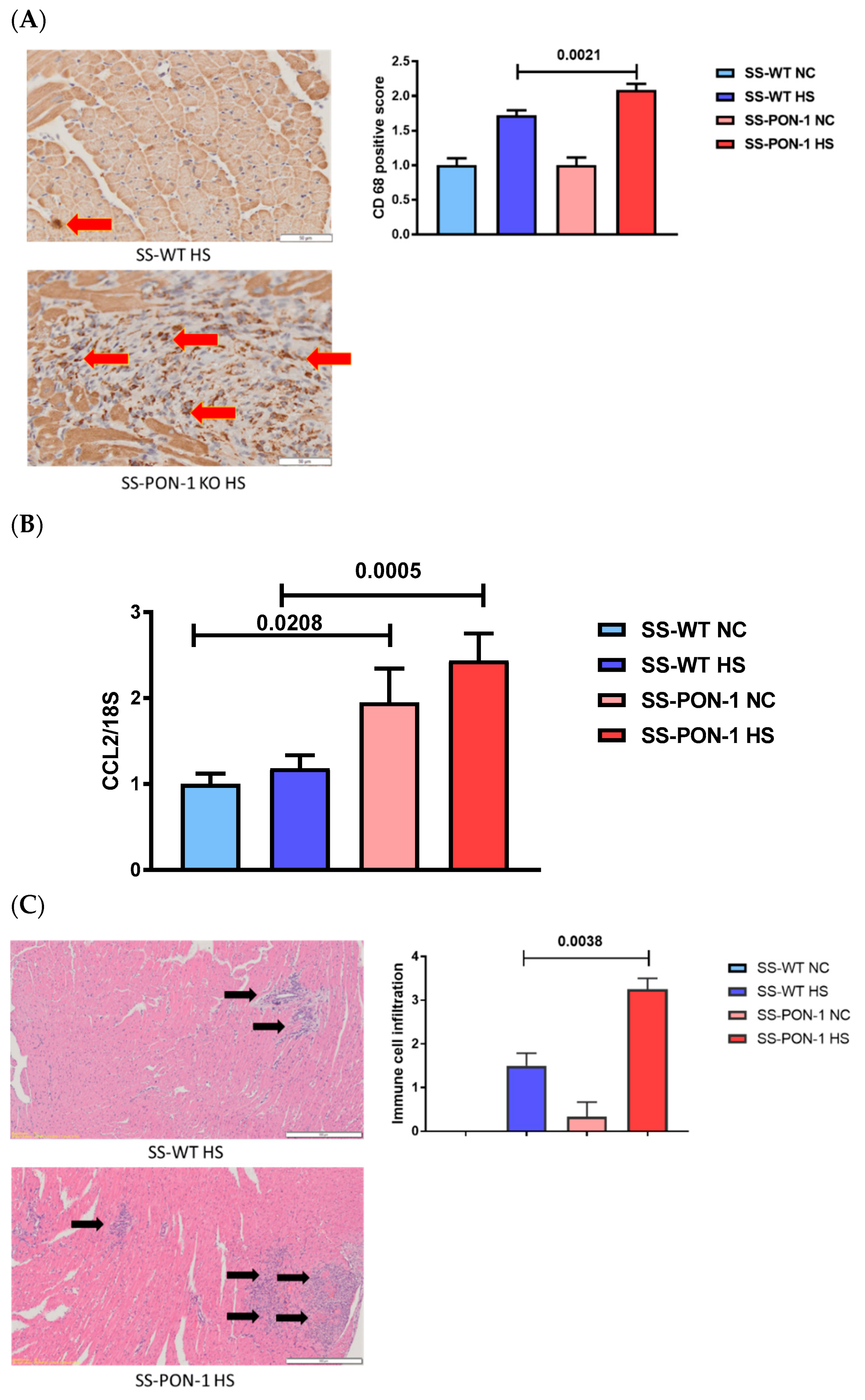
3.3. SS-PON-1 KO Rats on High-Salt Diet Demonstrate Significantly Increase in Cardiac Inflammation Compared to SS-WT Rats
3.4. SS-PON-1 KO Rats on High-Salt Diet Demonstrate Significantly Decreased Calcium Handling Genes Compared to SS-WT Rats
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gansevoort, R.T.; Correa-Rotter, R.; Hemmelgarn, B.R.; Jafar, T.H.; Heerspink, H.J.L.; Mann, J.F.; Matsushita, K.; Wen, C.P. Chronic kidney disease and cardiovascular risk: Epidemiology, mechanisms, and prevention. Lancet 2013, 382, 339–352. [Google Scholar] [CrossRef]
- Sarnak, M.J. Cardiovascular complications in chronic kidney disease. Am. J. Kidney Dis. 2003, 41, 11–17. [Google Scholar] [CrossRef]
- Herzog, C.A.; Asinger, R.W.; Berger, A.K.; Charytan, D.M.; Díez, J.; Hart, R.G.; Eckardt, K.-U.; Kasiske, B.L.; McCullough, P.A.; Passman, R.S.; et al. Cardiovascular disease in chronic kidney disease. A clinical update from Kidney Disease: Improving Global Outcomes (KDIGO). Kidney Int. 2011, 80, 572–586. [Google Scholar] [CrossRef]
- Saritas, T.; Floege, J. Cardiovascular disease in patients with chronic kidney disease. Herz 2020, 45, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Harlos, J.; Heidland, A. Hypertension as Cause and Consequence of Renal Disease in the 19th Century. Am. J. Nephrol. 1994, 14, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Foley, R.N.; Murray, A.M.; Li, S.; Herzog, C.A.; McBean, A.M.; Eggers, P.W.; Collins, A.J. Chronic Kidney Disease and the Risk for Cardiovascular Disease, Renal Replacement, and Death in the United States Medicare Population, 1998 to 1999. J. Am. Soc. Nephrol. 2005, 16, 489–495. [Google Scholar] [CrossRef]
- Kennedy, D.J.; Tang, W.H.W.; Fan, Y.; Wu, Y.; Mann, S.; Pepoy, M.; Hazen, S.L. Diminished Antioxidant Activity of High-Density Lipoprotein–Associated Proteins in Chronic Kidney Disease. J. Am. Hear. Assoc. 2013, 2, e000104. [Google Scholar] [CrossRef]
- Mohammed, C.J.; Xie, Y.; Brewster, P.S.; Ghosh, S.; Dube, P.; Sarsour, T.; Kleinhenz, A.L.; Crawford, E.L.; Malhotra, D.; James, R.W.; et al. Circulating Lactonase Activity but Not Protein Level of PON-1 Predicts Adverse Outcomes in Subjects with Chronic Kidney Disease. J. Clin. Med. 2019, 8, 1034. [Google Scholar] [CrossRef]
- Mohammed, C.J.; Lamichhane, S.; Connolly, J.A.; Soehnlen, S.M.; Khalaf, F.K.; Malhotra, D.; Haller, S.T.; Isailovic, D.; Kennedy, D.J. A PON for All Seasons: Comparing Paraoxonase Enzyme Substrates, Activity and Action including the Role of PON3 in Health and Disease. Antioxidants 2022, 11, 590. [Google Scholar] [CrossRef]
- Sentí, M.; Tomás, M.; Vila, J.; Marrugat, J.; Elosua, R.; Sala, J.; Masiá, R. Relationship of age-related myocardial infarction risk and Gln/Arg 192 variants of the human paraoxonase1 gene: The REGICOR study. Atherosclerosis 2001, 156, 443–449. [Google Scholar] [CrossRef]
- Tward, A.; Xia, Y.-R.; Wang, X.-P.; Shi, Y.-S.; Park, C.; Castellani, L.W.; Lusis, A.J.; Shih, D.M. Decreased Atherosclerotic Lesion Formation in Human Serum Paraoxonase Transgenic Mice. Circulation 2002, 106, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Mackness, I.M.; Abbott, C.; Arrol, S.; Durrington, P.N. The role of high-density lipoprotein and lipid-soluble antioxidant vitamins in inhibiting low-density lipoprotein oxidation. Biochem. J. 1993, 294, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Arrol, S.; Mackness, M.; Durrington, P. High density lipoprotein-associated enzymes and the prevention of low density lipoprotein oxidation. Eur. J. Lab. Med. 1996, 4, 33–38. [Google Scholar]
- Jayakumari, N.; Thejaseebai, G. High Prevalence of Low Serum Paraoxonase-1 in Subjects with Coronary Artery Disease. J. Clin. Biochem. Nutr. 2009, 45, 278–284. [Google Scholar] [CrossRef]
- Robertson, K.S.; Hawe, E.; Miller, G.J.; Talmud, P.J.; Humphries, S.E. Human paraoxonase gene cluster polymorphisms as predictors of coronary heart disease risk in the prospective Northwick Park Heart Study II. Biochim. et Biophys. Acta (BBA)-Mol. Basis Dis. 2003, 1639, 203–212. [Google Scholar] [CrossRef]
- Mackness, M.I.; Durrington, P.N.; Mackness, B. The Role of Paraoxonase 1 Activity in Cardiovascular Disease. Am. J. Cardiovasc. Drugs 2004, 4, 211–217. [Google Scholar] [CrossRef]
- Watson, A.D.; Berliner, J.A.; Hama, S.Y.; La Du, B.N.; Faull, K.F.; Fogelman, A.M.; Navab, M. Protective effect of high density lipoprotein associated paraoxonase. Inhibition of the biological activity of minimally oxidized low density lipoprotein. J. Clin. Investig. 1995, 96, 2882–2891. [Google Scholar] [CrossRef]
- Aviram, M.; Billecke, S.; Sorenson, R.; Bisgaier, C.; Newton, R.; Rosenblat, M.; Erogul, J.; Hsu, C.; Dunlop, C.; La Du, B. Paraoxonase active site required for protection against LDL oxidation involves its free sulfhydryl group and is different from that required for its arylesterase/paraoxonase activities: Selective action of human paraoxonase allozymes Q and R. Arter. Thromb. Vasc. Biol. 1998, 18, 1617–1624. [Google Scholar] [CrossRef]
- Mackness, M.; Arrol, S.; Abbott, C.; Durrington, P. Protection of low-density lipoprotein against oxidative modification by high-density lipoprotein associated paraoxonase. Atherosclerosis 1993, 104, 129–135. [Google Scholar] [CrossRef]
- Miljkovic, M.; Stefanovic, A.; Vekic, J.; Zeljkovic, A.; Gojkovic, T.; Simic-Ogrizovic, S.; Bogavac-Stanojevic, N.; Cerne, D.; Ilic, J.; Stefanovic, I.; et al. Activity of paraoxonase 1 (PON1) on HDL2 and HDL3 subclasses in renal disease. Clin. Biochem. 2018, 60, 52–58. [Google Scholar] [CrossRef]
- Khalaf, F.K.; Mohammed, C.J.; Dube, P.; Connolly, J.A.; Lad, A.; Ashraf, U.M.; Breidenbach, J.D.; Su, R.C.; Kleinhenz, A.L.; Malhotra, D.; et al. Paraoxonase-1 Regulation of Renal Inflammation and Fibrosis in Chronic Kidney Disease. Antioxidants 2022, 11, 900. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.E.; Faulx, M.D.; McElfresh, T.A.; Kung, T.A.; Zawaneh, M.S.; Stanley, W.C.; Chandler, M.P.; Hoit, B.D. Validation of echocardiographic methods for assessing left ventricular dysfunction in rats with myocardial infarction. Am. J. Physiol. Circ. Physiol. 2004, 287, H2049–H2053. [Google Scholar] [CrossRef] [PubMed]
- Cutler, M.J.; Wan, X.; Plummer, B.N.; Liu, H.; Deschenes, I.; Laurita, K.R.; Hajjar, R.J.; Rosenbaum, D.S. Targeted Sarcoplasmic Reticulum Ca 2+ ATPase 2a Gene Delivery to Restore Electrical Stability in the Failing Heart. Circulation 2012, 126, 2095–2104. [Google Scholar] [CrossRef] [PubMed]
- Stenvinkel, P. Chronic kidney disease: A public health priority and harbinger of premature cardiovascular disease. J. Intern. Med. 2010, 268, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Thompson, S.; James, M.; Wiebe, N.; Hemmelgarn, B.; Manns, B.; Klarenbach, S.; Tonelli, M. Cause of Death in Patients with Reduced Kidney Function. J. Am. Soc. Nephrol. 2015, 26, 2504–2511. [Google Scholar] [CrossRef]
- Mihai, S.; Codrici, E.; Popescu, I.D.; Enciu, A.-M.; Albulescu, L.; Necula, L.G.; Mambet, C.; Anton, G.; Tanase, C. Inflammation-Related Mechanisms in Chronic Kidney Disease Prediction, Progression, and Outcome. J. Immunol. Res. 2018, 2018, 2180373. [Google Scholar] [CrossRef]
- Kanbay, M.; Solak, Y.; Unal, H.U.; Kurt, Y.G.; Gok, M.; Cetinkaya, H.; Karaman, M.; Oguz, Y.; Eyileten, T.; Vural, A.; et al. Monocyte count/HDL cholesterol ratio and cardiovascular events in patients with chronic kidney disease. Int. Urol. Nephrol. 2014, 46, 1619–1625. [Google Scholar] [CrossRef]
- Patra, S.K.; Singh, K.; Singh, R. Paraoxonase 1: A better atherosclerotic risk predictor than HDL in type 2 diabetes mellitus. Diabetes Metab. Syndr. Clin. Res. Rev. 2013, 7, 108–111. [Google Scholar] [CrossRef]
- Otocka-Kmiecik, A.; Mikhailidis, D.P.; Nicholls, S.J.; Davidson, M.; Rysz, J.; Banach, M. Dysfunctional HDL: A novel important diagnostic and therapeutic target in cardiovascular disease? Prog. Lipid Res. 2012, 51, 314–324. [Google Scholar] [CrossRef]
- Tavori, H.; Aviram, M.; Khatib, S.; Musa, R.; Nitecki, S.; Hoffman, A.; Vaya, J. Human carotid atherosclerotic plaque increases oxidative state of macrophages and low-density lipoproteins, whereas paraoxonase 1 (PON1) decreases such atherogenic effects. Free Radic. Biol. Med. 2009, 46, 607–615. [Google Scholar] [CrossRef]
- Mackness, B.; Turkie, W.; Mackness, M. Paraoxonase-1 (PON1) promoter region polymorphisms, serum PON1 status and coronary heart disease. Arch. Med Sci. 2013, 1, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Melnichenko, A.A.; Orekhov, A.N.; Bobryshev, Y.V. Paraoxonase and atherosclerosis-related cardiovascular diseases. Biochimie 2017, 132, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Mackness, B.; Durrington, P.; McElduff, P.; Yarnell, J.; Azam, N.; Watt, M.; Mackness, M. Low Paraoxonase Activity Predicts Coronary Events in the Caerphilly Prospective Study. Circulation 2003, 107, 2775–2779. [Google Scholar] [CrossRef] [PubMed]
- Shih, D.M.; Gu, L.; Xia, Y.-R.; Navab, M.; Li, W.-F.; Hama, S.; Castellani, L.W.; Furlong, C.E.; Costa, L.G.; Fogelman, A.M.; et al. Mice lacking serum paraoxonase are susceptible to organophosphate toxicity and atherosclerosis. Nature 1998, 394, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.W.; Wu, Y.; Mann, S.; Pepoy, M.; Shrestha, K.; Borowski, A.G.; Hazen, S.L. Diminished Antioxidant Activity of High-Density Lipoprotein–Associated Proteins in Systolic Heart Failure. Circ. Heart Fail. 2011, 4, 59–64. [Google Scholar] [CrossRef]
- Hammadah, M.; Kalogeropoulos, A.P.; Georgiopoulou, V.V.; Weber, M.; Wu, Y.; Hazen, S.L.; Butler, J.; Tang, W.W. High-density lipoprotein-associated paraoxonase-1 activity for prediction of adverse outcomes in outpatients with chronic heart failure. Eur. J. Heart Fail. 2017, 19, 748–755. [Google Scholar] [CrossRef]
- Cicila, G.T.; Morgan, E.E.; Lee, S.J.; Farms, P.; Yerga-Woolwine, S.; Toland, E.J.; Ramdath, R.S.; Gopalakrishnan, K.; Bohman, K.; Nestor-Kalinoski, A.L.; et al. Epistatic Genetic Determinants of Blood Pressure and Mortality in a Salt-Sensitive Hypertension Model. Hypertension 2009, 53, 725–732. [Google Scholar] [CrossRef]
- Gugliucci, A.; Kotani, K.; Kimura, S. Paraoxonase 1 in Chronic Kidney Failure. J. Lipids 2012, 2012, 726048. [Google Scholar] [CrossRef]
- Causes of death. USRDS. United States Renal Data System. Am. J. Kidney Dis. 1997, 30, S107–S117.
- Efe, T.H.; Ertem, A.G.; Altunoglu, A.; Koseoglu, C.; Erayman, A.; Bilgin, M.; Kurmuş, Ö.; Aslan, T.; Bilge, M. Serum Paraoxonase Levels are Correlated with Impaired Aortic Functions in Patients with Chronic Kidney Disease. Acta Cardiol. Sin. 2016, 32, 75–80. [Google Scholar] [CrossRef]
- Paragh, G.; Seres, I.; Balogh, Z.; Varga, Z.; Kárpáti, I.; Mátyus, J.; Újhelyi, L.; Kakuk, G. The Serum Paraoxonase Activity in Patients with Chronic Renal Failure and Hyperlipidemia. Nephron Exp. Nephrol. 1998, 80, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Dantoine, T.F.; Debord, J.; Charmes, J.P.; Merle, L.; Marquet, P.; Lachatre, G.; Leroux-Robert, C. Decrease of serum paraoxonase activity in chronic renal failure. J. Am. Soc. Nephrol. 1998, 9, 2082–2088. [Google Scholar] [CrossRef] [PubMed]
- Henning, B.F.; Holzhausen, H.; Tepel, M. Continuous Reduction of Plasma Paraoxonase Activity with Increasing Dialysis Vintage in Hemodialysis Patients. Ther. Apher. Dial. 2010, 14, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Kannampuzha, J.; Darling, P.B.; Maguire, G.F.; Donnelly, S.; McFarlane, P.; Chan, C.T.; Connelly, P. Paraoxonase 1 arylesterase activity and mass are reduced and inversely related to C-reactive protein in patients on either standard or home nocturnal hemodialysis. Clin. Nephrol. 2010, 73, 131–138. [Google Scholar]
- Dirican, M.; Akca, R.; Sarandol, E.; Dilek, K. Serum paraoxonase activity in uremic predialysis and hemodialysis patients. J. Nephrol. 2004, 17, 813–818. [Google Scholar]
- Rebrova, T.Y.; Muslimova, E.F.; Kondratieva, D.S.; Budnikova, O.V.; Ahmedov, S.D.; Afanasiev, S.A.; Popov, S.V. The Role of Ca2+-ATPase 2a (ATP2A2), Ryanodine Receptors (RYR2), and Calsequestrin (CASQ2) Gene Polymorphisms in the Development of Heart Failure. Russ. J. Genet. 2018, 54, 604–608. [Google Scholar] [CrossRef]
- Angrisano, T.; Schiattarella, G.; Keller, S.; Pironti, G.; Florio, E.; Magliulo, F.; Bottino, R.; Pero, R.; Lembo, F.; Avvedimento, E.V.; et al. Epigenetic Switch at Atp2a2 and Myh7 Gene Promoters in Pressure Overload-Induced Heart Failure. PLoS ONE 2014, 9, e106024. [Google Scholar] [CrossRef]
- Kennedy, D.; Omran, E.; Periyasamy, S.M.; Nadoor, J.; Priyadarshi, A.; Willey, J.C.; Malhotra, D.; Xie, Z.; Shapiro, J.I. Effect of Chronic Renal Failure on Cardiac Contractile Function, Calcium Cycling, and Gene Expression of Proteins Important for Calcium Homeostasis in the Rat. J. Am. Soc. Nephrol. 2003, 14, 90–97. [Google Scholar] [CrossRef]
- Kennedy, D.J.; Vetteth, S.; Periyasamy, S.M.; Kanj, M.; Fedorova, L.; Khouri, S.; Kahaleh, M.B.; Xie, Z.; Malhotra, D.; Kolodkin, N.I.; et al. Central Role for the Cardiotonic Steroid Marinobufagenin in the Pathogenesis of Experimental Uremic Cardiomyopathy. Hypertension 2006, 47, 488–495. [Google Scholar] [CrossRef]
- Elkareh, J.; Kennedy, D.J.; Yashaswi, B.; Vetteth, S.; Shidyak, A.; Kim, E.G.R.; Smaili, S.; Periyasamy, S.M.; Hariri, I.M.; Fedorova, L.; et al. Marinobufagenin Stimulates Fibroblast Collagen Production and Causes Fibrosis in Experimental Uremic Cardiomyopathy. Hypertension 2007, 49, 215–224. [Google Scholar] [CrossRef]
- Sztanek, F.; Seres, I.; Harangi, M.; Lőcsey, L.; Padra, J.; Paragh, G.J.; Asztalos, L. Decreased paraoxonase 1 (PON1) lactonase activity in hemodialyzed and renal transplanted patients. A novel cardiovascular biomarker in end-stage renal disease. Nephrol. Dial. Transplant. 2012, 27, 2866–2872. [Google Scholar] [CrossRef] [PubMed]
- Marsillach, J.; Martínez-Vea, A.; Marcas, L.; Mackness, B.; Mackness, M.; Ferré, N.; Joven, J.; Camps, J. Administration of exogenous erythropoietin β affects lipid peroxidation and serum paraoxonase-1 activity and concentration in predialysis patients with chronic renal disease and anaemia. Clin. Exp. Pharmacol. Physiol. 2007, 34, 347–349. [Google Scholar] [CrossRef]
- Lou-Bonafonte, J.M.; Gabás-Rivera, C.; Navarro, M.A.; Osada, J. The Search for Dietary Supplements to Elevate or Activate Circulating Paraoxonases. Int. J. Mol. Sci. 2017, 18, 416. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, S.; Kisa, F.; Karadeniz, A.; Yildrim, A.; Karakoc, A.; Can, I.; Kara, A.; Simsek, N. Effects of pomegranate seed extract on liver paraoxonase and bcl-xL activities in rats treated with cisplatin. J. Med. Plants Res. 2012, 6, 2317–2323. [Google Scholar]
- Martini, D.; Del Bo’, C.; Porrini, M.; Ciappellano, S.; Riso, P. Role of polyphenols and polyphenol-rich foods in the modulation of PON1 activity and expression. J. Nutr. Biochem. 2017, 48, 1–8. [Google Scholar] [CrossRef]
- Lou-Bonafonte, J.M.; Gabás-Rivera, C.; Navarro, M.A.; Osada, J. PON1 and Mediterranean Diet. Nutrients 2015, 7, 4068–4092. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dube, P.; Khalaf, F.K.; DeRiso, A.; Mohammed, C.J.; Connolly, J.A.; Battepati, D.; Lad, A.; Breidenbach, J.D.; Kleinhenz, A.L.; Khatib-Shahidi, B.; et al. Cardioprotective Role for Paraoxonase-1 in Chronic Kidney Disease. Biomedicines 2022, 10, 2301. https://doi.org/10.3390/biomedicines10092301
Dube P, Khalaf FK, DeRiso A, Mohammed CJ, Connolly JA, Battepati D, Lad A, Breidenbach JD, Kleinhenz AL, Khatib-Shahidi B, et al. Cardioprotective Role for Paraoxonase-1 in Chronic Kidney Disease. Biomedicines. 2022; 10(9):2301. https://doi.org/10.3390/biomedicines10092301
Chicago/Turabian StyleDube, Prabhatchandra, Fatimah K. Khalaf, Armelle DeRiso, Chrysan J. Mohammed, Jacob A. Connolly, Dhanushya Battepati, Apurva Lad, Joshua D. Breidenbach, Andrew L. Kleinhenz, Bella Khatib-Shahidi, and et al. 2022. "Cardioprotective Role for Paraoxonase-1 in Chronic Kidney Disease" Biomedicines 10, no. 9: 2301. https://doi.org/10.3390/biomedicines10092301
APA StyleDube, P., Khalaf, F. K., DeRiso, A., Mohammed, C. J., Connolly, J. A., Battepati, D., Lad, A., Breidenbach, J. D., Kleinhenz, A. L., Khatib-Shahidi, B., Patel, M., Tassavvor, I., Gohara, A. F., Malhotra, D., Morgan, E. E., Haller, S. T., & Kennedy, D. J. (2022). Cardioprotective Role for Paraoxonase-1 in Chronic Kidney Disease. Biomedicines, 10(9), 2301. https://doi.org/10.3390/biomedicines10092301