
Dipeptidyl Peptidase 4 (DPP4) as A Novel Adipokine: Role in Metabolism and Fat Homeostasis



{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. DPP4: Forms and Functions
1.2. DPP4 Expression and Secretion
2. DPP4 and Metabolism: Role of DPP4 in the Regulation of Glucose Metabolism
2.1. DPP4 and Its Substrates: Incretin System
2.1.1. Glucagon-like Peptide 1 (GLP-1)
2.1.2. Glucagon-like Peptide 2 (GLP-2)
2.1.3. Glucose-Dependent Insulinotropic Peptide (GIP)
2.1.4. Neuropeptide Y (NPY), Peptide YY, and Pancreatic Polypeptide
2.1.5. Insulin-like Growth Factor 1 (IGF-1)
2.1.6. Stromal-Derived Factor 1 (SDF-1)
2.1.7. Substance P
2.2. DPP4 and Insulin Resistance
2.3. DPP4, Emotional Behavior, and Physical Activity
2.4. DPP4 and Pancreatic β Cell Survival
3. DPP4 and Lipid Metabolism
4. DPP4 in Adipose Tissue and Fat Distribution
4.1. DPP4 and the Adipose Tissue
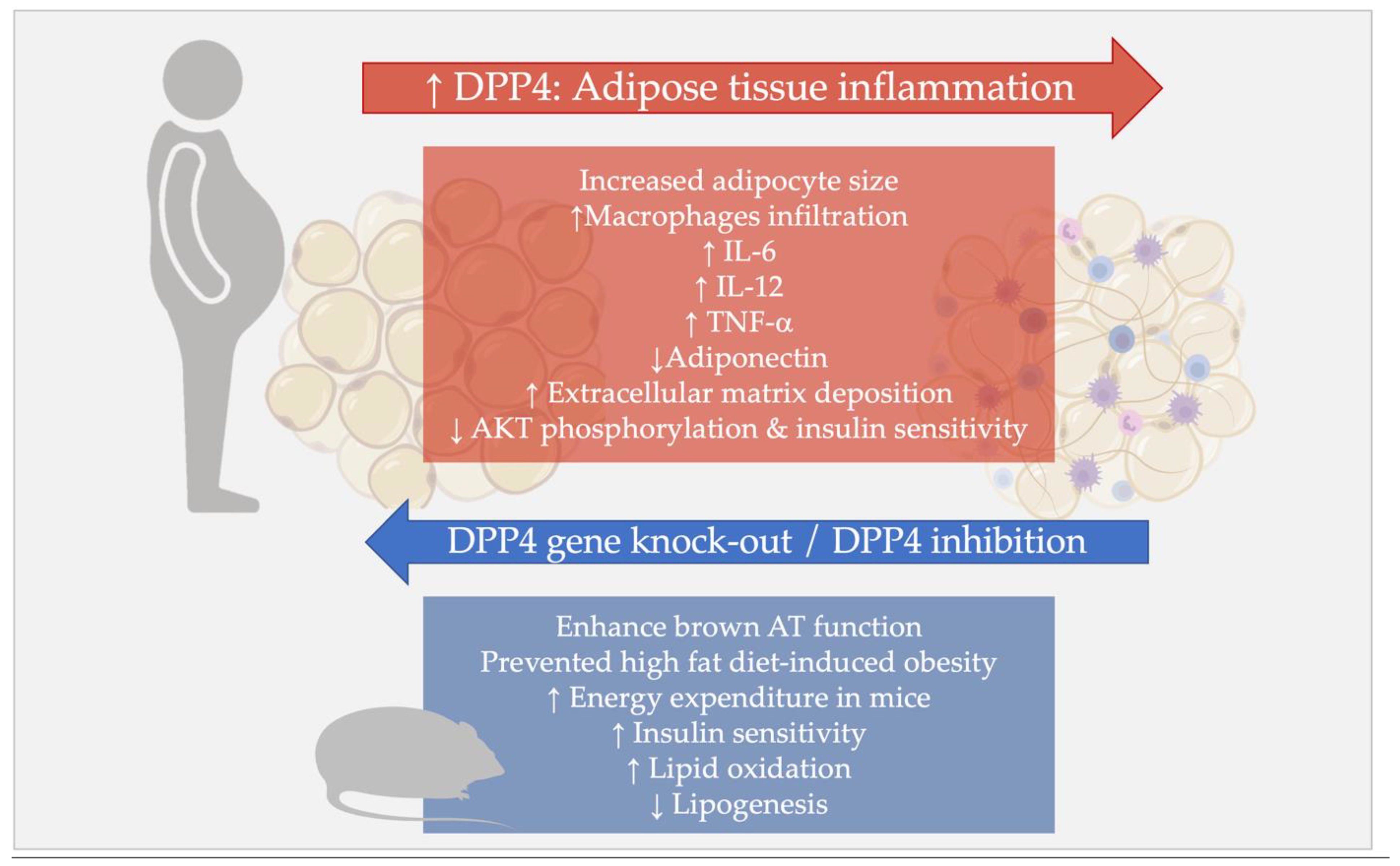
4.2. DPP4 and Sick Fat: Role of DPP4 in the Adipose Tissue Inflammation
4.3. DPP4 and Ectopic Fat Deposition
5. DPP4 in Metabolic Diseases
5.1. Obesity
5.2. Type 2 Diabetes Mellitus (T2DM)
5.3. Nonalcoholic Fatty Liver Disease (NAFLD) and Steatohepatitis (NASH)
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hopsu-Havu, V.K.; Glenner, G.G. A new dipeptide naphthylamidase hydrolyzing glycyl-prolyl-beta-naphthylamide. Histochemie 1966, 7, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, A.J.; Mattern, T.; Feller, A.C.; Heymann, E.; Flad, H.D. CD26 antigen is a surface dipeptidyl peptidase IV (DPPIV) as characterized by monoclonal antibodies clone TII-19-4-7 and 4EL1C7. Scand. J. Immunol. 1990, 31, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Abbott, C.; McCaughan, G.; Baker, E.; Sutherland, G. Genomic organization, exact localization, and tissue expression of the human CD26 (dipeptidyl peptidase IV) gene. Immunogenetics 1994, 40, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.M.; Zhang, Y.Y.; Hung, J.S.; Chung, J.Y.; Huang, X.R.; To, K.F.; Lan, H.Y. DPP4/CD32b/NF-κB Circuit: A Novel Druggable Target for Inhibiting CRP-Driven Diabetic Nephropathy. Mol. Ther. 2021, 29, 365–375. [Google Scholar] [CrossRef]
- Senkel, S.; Lucas, B.; Klein-Hitpass, L.; Ryffel, G.U. Identification of target genes of the transcription factor HNF1beta and HNF1alpha in a human embryonic kidney cell line. Biochim. Biophys. Acta 2005, 1731, 179–190. [Google Scholar] [CrossRef]
- Du, J.; Fu, L.; Ji, F.; Wang, C.; Liu, S.; Qiu, X. FosB recruits KAT5 to potentiate the growth and metastasis of papillary thyroid cancer in a DPP4-dependent manner. Life Sci. 2020, 259, 118374. [Google Scholar] [CrossRef]
- Bauvois, B.; Djavaheri-Mergny, M.; Rouillard, D.; Dumont, J.; Wietzerbin, J. Regulation of CD26/DPPIV gene expression by interferons and retinoic acid in tumor B cells. Oncogene 2000, 19, 265–272. [Google Scholar] [CrossRef]
- Dunaevsky, Y.E.; Tereshchenkova, V.F.; Oppert, B.; Belozersky, M.A.; Filippova, I.Y.; Elpidina, E.N. Human proline specific peptidases: A comprehensive analysis. BiochimBiophys Acta Gen. Subj. 2020, 1864, 129636. [Google Scholar] [CrossRef]
- Wagner, L.; Klemann, C.; Stephan, M.; von Hörsten, S. Unravelling the immunological roles of dipeptidyl peptidase 4 (DPP4) activity and/or structure homologue (DASH) proteins. Clin. Exp. Immunol. 2016, 184, 265–283. [Google Scholar] [CrossRef]
- Engel, M.; Hoffmann, T.; Wagner, L.; Wermann, M.; Heiser, U.; Kiefersauer, R.; Huber, R.; Bode, W.; Demuth, H.U.; Brandstetter, H. The crystal structure of dipeptidyl peptidase IV (CD26) reveals its functional regulation and enzymatic mechanism. Proc. Natl. Acad. Sci. USA 2003, 100, 5063–5068. [Google Scholar] [CrossRef] [Green Version]
- Nagatsu, I.; Nagatsu, T.; Yamamoto, T. Hydrolysis of amino acid beta-naphthylamides by aminopeptidases in human parotid salva and human serum. Experientia 1968, 24, 347–348. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.M.; Cheng, J.H.; Suen, C.S.; Huang, C.H.; Tsai, C.H.; Huang, L.H.; Chen, Y.R.; Wang, A.H.; Jiaang, W.T.; Hwang, M.J.; et al. The dimeric transmembrane domain of prolyl dipeptidase DPP-IV contributes to its quaternary structure and enzymatic activities. Protein Sci. 2010, 19, 1627–1638. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Meng, W.; Kilian, C.; Grams, S.; Reutter, W. Domain-specific N-glycosylation of the membrane glycoprotein dipeptidylpeptidase IV (CD26) influences its subcellular trafficking, biological stability, enzyme activity and protein folding. Eur. J. Biochem. 1997, 246, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Slimane, T.A.; Lenoir, C.; Sapin, C.; Maurice, M.; Trugnan, G. Apical secretion and sialylation of soluble dipeptidyl peptidase IV are two related events. Exp. Cell Res. 2000, 258, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.E.; Talhouk, J.W.; Brown, E.E.; Edgar, S.E. The significance of hypersialylation of dipeptidyl peptidase IV (CD26) in the inhibition of its activity by Tat and other cationic peptides. CD26: A subverted adhesion molecule for HIV peptide binding. AIDS Res. Hum. Retrovir. 1998, 14, 851–868. [Google Scholar] [CrossRef]
- Lone, A.M.; Nolte, W.M.; Tinoco, A.D.; Saghatelian, A. Peptidomics of the prolyl peptidases. AAPS J. 2010, 12, 483–491. [Google Scholar] [CrossRef]
- Zhong, J.; Rao, X.; Rajagopalan, S. An emerging role of dipeptidyl peptidase 4 (DPP4) beyond glucose control: Potential implications in cardiovascular disease. Atherosclerosis 2013, 226, 305–314. [Google Scholar] [CrossRef]
- Zhulai, G.; Oleinik, E.; Shibaev, M.; Ignatev, K. Adenosine-Metabolizing Enzymes, Adenosine Kinase and Adenosine Deaminase, in Cancer. Biomolecules 2022, 12, 418. [Google Scholar] [CrossRef]
- Gao, Z.W.; Wang, X.; Zhang, H.Z.; Lin, F.; Liu, C.; Dong, K. The roles of adenosine deaminase in autoimmune diseases. Autoimmun. Rev. 2021, 20, 102709. [Google Scholar] [CrossRef]
- Hu, X.; Chen, S.; Xie, C.; Li, Z.; Wu, Z.; You, Z. DPP4 gene silencing inhibits proliferation and epithelial-mesenchymal transition of papillary thyroid carcinoma cells through suppression of the MAPK pathway. J. Endocrinol. Investig. 2021, 44, 1609–1623. [Google Scholar] [CrossRef]
- Huang, J.; Liu, X.; Wei, Y.; Li, X.; Gao, S.; Dong, L.; Rao, X.; Zhong, J. Emerging Role of Dipeptidyl Peptidase-4 in Autoimmune Disease. Front. Immunol. 2022, 13, 830863. [Google Scholar] [CrossRef] [PubMed]
- Piazza, G.A.; Callanan, H.M.; Mowery, J.; Hixson, D.C. Evidence for a role of dipeptidyl peptidase IV in fibronectin-mediated interactions of hepatocytes with extracellular matrix. Biochem. J. 1989, 262, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Löster, K.; Zeilinger, K.; Schuppan, D.; Reutter, W. The cysteine-rich region of dipeptidyl peptidase IV (CD 26) is the collagen-binding site. Biochem. Biophys. Res. Commun. 1995, 217, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Anoop, S.; Misra, A.; Bhatt, S.P.; Gulati, S.; Pandey, R.M.; Mahajan, H. High circulating plasma dipeptidyl peptidase- 4 levels in non-obese Asian Indians with type 2 diabetes correlate with fasting insulin and LDL-C levels, triceps skinfolds, total intra-abdominal adipose tissue volume and presence of diabetes: A case-control study. BMJ Open Diabetes Res. Care 2017, 5, e000393. [Google Scholar] [CrossRef] [PubMed]
- Barchetta, I.; Ceccarelli, V.; Cimini, F.A.; Barone, E.; Sentinelli, F.; Coluzzi, M.; Chiappetta, C.; Bertoccini, L.; Tramutola, A.; Labbadia, G.; et al. Circulating dipeptidyl peptidase-4 is independently associated with the presence and severity of NAFLD/NASH in individuals with and without obesity and metabolic disease. J. Endocrinol. Investig. 2021, 44, 979–988. [Google Scholar] [CrossRef]
- Blauschmidt, S.; Greither, T.; Lampe, K.; Köller, S.; Kaltwaßer, P.; Behre, H.M. Dipeptidyl peptidase 4 serum activity and concentration are increased in women with polycystic ovary syndrome. Clin. Endocrinol. 2017, 87, 741–747. [Google Scholar] [CrossRef]
- Sedighi, M.; Baluchnejadmojarad, T.; Fallah, S.; Moradi, N.; Afshin-Majd, S.; Roghani, M. The Association Between Circulating Klotho and Dipeptidyl Peptidase-4 Activity and Inflammatory Cytokines in Elderly Patients With Alzheimer Disease. Basic Clin. Neurosci. 2020, 11, 349–357. [Google Scholar] [CrossRef]
- Lamers, D.; Famulla, S.; Wronkowitz, N.; Hartwig, S.; Lehr, S.; Ouwens, D.M.; Eckardt, K.; Kaufman, J.M.; Ryden, M.; Müller, S.; et al. Dipeptidyl peptidase 4 is a novel adipokine potentially linking obesity to the metabolic syndrome. Diabetes 2011, 60, 1917–1925. [Google Scholar] [CrossRef]
- Röhrborn, D.; Eckel, J.; Sell, H. Shedding of dipeptidyl peptidase 4 is mediated by metalloproteases and up-regulated by hypoxia in human adipocytes and smooth muscle cells. FEBS Lett. 2014, 588, 3870–3877. [Google Scholar] [CrossRef]
- McKillop, A.M.; Stevenson, C.L.; Moran, B.M.; Abdel-Wahab, Y.H.A.; Flatt, P.R. Tissue expression of DPP-IV in obesity-diabetes and modulatory effects on peptide regulation of insulin secretion. Peptides 2018, 100, 165–172. [Google Scholar] [CrossRef]
- Varin, E.M.; Mulvihill, E.E.; Beaudry, J.L.; Pujadas, G.; Fuchs, S.; Tanti, J.F.; Fazio, S.; Kaur, K.; Cao, X.; Baggio, L.L.; et al. Circulating Levels of Soluble Dipeptidyl Peptidase-4 Are Dissociated from Inflammation and Induced by Enzymatic DPP4 Inhibition. Cell Metab. 2019, 29, 320–334.e5. [Google Scholar] [CrossRef] [PubMed]
- Baggio, L.L.; Varin, E.M.; Koehler, J.A.; Cao, X.; Lokhnygina, Y.; Stevens, S.R.; Holman, R.R.; Drucker, D.J. Plasma levels of DPP4 activity and sDPP4 are dissociated from inflammation in mice and humans. Nat. Commun. 2020, 11, 3766. [Google Scholar] [CrossRef]
- Draznin, B.; Aroda, V.R.; Bakris, G.; Benson, G.; Brown, F.M.; Freeman, R.; Green, J.; Huang, E.; Isaacs, D.; Kahan, S.; et al. 9. Pharmacologic Approaches to Glycemic Treatment: Standards of Medical Care in Diabetes-2022. Diabetes Care 2022, 45, S125–S143. [Google Scholar] [CrossRef] [PubMed]
- Buzzetti, R.; Tuomi, T.; Mauricio, D.; Pietropaolo, M.; Zhou, Z.; Pozzilli, P.; Leslie, R.D. Management of Latent Autoimmune Diabetes in Adults: A Consensus Statement From an International Expert Panel. Diabetes 2020, 69, 2037–2047. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Xu, Y.; Wang, X.; Yang, L.; Zhao, D. Role of Dipeptidyl Peptidase 4 Inhibitors in Antidiabetic Treatment. Molecules 2022, 27, 3055. [Google Scholar] [CrossRef] [PubMed]
- Nagakura, T.; Yasuda, N.; Yamazaki, K.; Ikuta, H.; Yoshikawa, S.; Asano, O.; Tanaka, I. Improved glucose tolerance via enhanced glucose-dependent insulin secretion in dipeptidyl peptidase IV-deficient Fischer rats. Biochem. Biophys. Res. Commun. 2001, 284, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Marguet, D.; Baggio, L.; Kobayashi, T.; Bernard, A.M.; Pierres, M.; Nielsen, P.F.; Ribel, U.; Watanabe, T.; Drucker, D.J.; Wagtmann, N. Enhanced insulin secretion and improved glucose tolerance in mice lacking CD26. Proc. Natl. Acad. Sci. USA 2000, 97, 6874–6879. [Google Scholar] [CrossRef]
- Yasuda, N.; Nagakura, T.; Yamazaki, K.; Inoue, T.; Tanaka, I. Improvement of high fat-diet-induced insulin resistance in dipeptidyl peptidase IV-deficient Fischer rats. Life Sci. 2002, 71, 227–238. [Google Scholar] [CrossRef]
- Conarello, S.L.; Li, Z.; Ronan, J.; Roy, R.S.; Zhu, L.; Jiang, G.; Liu, F.; Woods, J.; Zycband, E.; Moller, D.E.; et al. Mice lacking dipeptidyl peptidase IV are protected against obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2003, 100, 6825–6830. [Google Scholar] [CrossRef]
- Mitani, H.; Takimoto, M.; Hughes, T.E.; Kimura, M. Dipeptidyl peptidase IV inhibition improves impaired glucose tolerance in high-fat diet-fed rats: Study using a Fischer 344 rat substrain deficient in its enzyme activity. Jpn. J. Pharmacol. 2002, 88, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Baerts, L.; Waumans, Y.; Brandt, I.; Jungraithmayr, W.; Van der Veken, P.; Vanderheyden, M.; De Meester, I. Circulating stromal cell-derived factor 1alpha levels in heart failure: A matter of proper sampling. PLoS ONE 2015, 10, e0141408. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Choi, B.K.; Li, W.; Lao, Z.; Lee, A.Y.; Souza, S.C.; Yates, N.A.; Kowalski, T.; Pocai, A.; Cohen, L.H. Quantification of intact and truncated stromal cell-derived factor-1α in circulation by immunoaffinity enrichment and tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2014, 25, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Elmansi, A.M.; Awad, M.E.; Eisa, N.H.; Kondrikov, D.; Hussein, K.A.; Aguilar-Pérez, A.; Herberg, S.; Periyasamy-Thandavan, S.; Fulzele, S.; Hamrick, M.W.; et al. What doesn’t kill you makes you stranger: Dipeptidyl peptidase-4 (CD26) proteolysis differentially modulates the activity of many peptide hormones and cytokines generating novel cryptic bioactive ligands. Pharmacol. Ther. 2019, 198, 90–108. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J. The incretin system in healthy humans: The role of GIP and GLP-1. Metabolism 2019, 96, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Lafferty, R.A.; O’Harte, F.P.M.; Irwin, N.; Gault, V.A.; Flatt, P.R. Proglucagon-Derived Peptides as Therapeutics. Front. Endocrinol. 2021, 12, 689678. [Google Scholar] [CrossRef]
- Kieffer, T.J.; McIntosh, C.H.; Pederson, R.A. Degradation of glucose-dependent insulinotropic polypeptide and truncated glucagon-like peptide 1 in vitro and in vivo by dipeptidyl peptidase IV. Endocrinology 1995, 136, 3585–3596. [Google Scholar] [CrossRef]
- Jonik, S.; Marchel, M.; Grabowski, M.; Opolski, G.; Mazurek, T. Gastrointestinal Incretins-Glucose-Dependent Insulinotropic Polypeptide (GIP) and Glucagon-like Peptide-1 (GLP-1) beyond Pleiotropic Physiological Effects Are Involved in Pathophysiology of Atherosclerosis and Coronary Artery Disease-State of the Art. Biology 2022, 11, 288. [Google Scholar] [CrossRef]
- Osinski, C.; Le Gléau, L.; Poitou, C.; de Toro-Martin, J.; Genser, L.; Fradet, M.; Soula, H.A.; Leturque, A.; Blugeon, C.; Jourdren, L.; et al. Type 2 diabetes is associated with impaired jejunal enteroendocrine GLP-1 cell lineage in human obesity. Int. J. Obes. 2021, 45, 170–183. [Google Scholar] [CrossRef]
- Kounatidis, D.; Vallianou, N.G.; Tsilingiris, D.; Christodoulatos, G.S.; Geladari, E.; Stratigou, T.; Karampela, I.; Dalamaga, M. Therapeutic Potential of GLP-2 Analogs in Gastrointestinal Disorders: Current Knowledge, Nutritional Aspects, and Future Perspectives. Curr. Nutr. Rep. 2022, 1–25. [Google Scholar] [CrossRef]
- Sun, H.; Meng, K.; Hou, L.; Shang, L.; Yan, J. Melanocortin receptor-4 mediates the anorectic effect induced by the nucleus tractus solitarius injection of glucagon-like Peptide-2 in fasted rats. Eur. J. Pharmacol. 2021, 901, 174072. [Google Scholar] [CrossRef]
- Ejarque, M.; Sabadell-Basallote, J.; Beiroa, D.; Calvo, E.; Keiran, N.; Nuñez-Roa, C.; Rodríguez, M.D.M.; Sabench, F.; Del Castillo, D.; Jimenez, V.; et al. Adipose tissue is a key organ for the beneficial effects of GLP-2 metabolic function. Br. J. Pharmacol. 2021, 178, 2131–2145. [Google Scholar] [CrossRef] [PubMed]
- Lu, V.B.; Gribble, F.M.; Reimann, F. Nutrient-Induced Cellular Mechanisms of Gut Hormone Secretion. Nutrients 2021, 13, 883. [Google Scholar] [CrossRef]
- Frerker, N.; Wagner, L.; Wolf, R.; Heiser, U.; Hoffmann, T.; Rahfeld, J.U.; Schade, J.; Karl, T.; Naim, H.Y.; Alfalah, M.; et al. Neuropeptide Y (NPY) cleaving enzymes: Structural and functional homologues of dipeptidyl peptidase 4. Peptides 2007, 28, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Grandt, D.; Schimiczek, M.; Beglinger, C.; Layer, P.; Goebell, H.; Eysselein, V.E.; Reeve, J.R., Jr. Two molecular forms of peptide YY (PYY) are abundant in human blood: Characterization of a radioimmunoassay recognizing PYY 1-36 and PYY 3-36. Regul. Pept. 1994, 51, 151–159. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, C.Y.; Liu, Y.W.; Rao, S.S.; Tan, Y.J.; Qian, Y.X.; Xia, K.; Huang, J.; Liu, X.X.; Hong, C.G.; et al. Neuronal Induction of Bone-Fat Imbalance through Osteocyte Neuropeptide Y. Adv. Sci. 2021, 8, e2100808. [Google Scholar] [CrossRef] [PubMed]
- Rodnoi, P.; Rajkumar, M.; Moin, A.S.M.; Georgia, S.K.; Butler, A.E.; Dhawan, S. Neuropeptide Y expression marks partially differentiated β cells in mice and humans. JCI Insight 2017, 2, e94005. [Google Scholar] [CrossRef] [PubMed]
- Myrsén-Axcrona, U.; Ekblad, E.; Sundler, F. Developmental expression of NPY, PYY and PP in the rat pancreas and their coexistence with islet hormones. Regul. Pept. 1997, 68, 165–175. [Google Scholar] [CrossRef]
- Morgan, D.G.; Kulkarni, R.N.; Hurley, J.D.; Wang, Z.L.; Wang, R.M.; Ghatei, M.A.; Karlsen, A.E.; Bloom, S.R.; Smith, D.M. Inhibition of glucose stimulated insulin secretion by neuropeptide Y is mediated via the Y1 receptor and inhibition of adenylyl cyclase in RIN 5AH rat insulinoma cells. Diabetologia 1998, 41, 1482–1491. [Google Scholar] [CrossRef]
- Crooks, B.; Stamataki, N.S.; McLaughlin, J.T. Appetite, the enteroendocrine system, gastrointestinal disease and obesity. Proc. Nutr. Soc. 2021, 80, 50–58. [Google Scholar] [CrossRef]
- Banks, W.A.; Kastin, A.J. Differential permeability of the blood-brain barrier to two pancreatic peptides: Insulin and amylin. Peptides 1998, 19, 883–889. [Google Scholar] [CrossRef]
- Giustina, A.; Berardelli, R.; Gazzaruso, C.; Mazziotti, G. Insulin and GH-IGF-I axis: Endocrine pacer or endocrine disruptor? Acta Diabetol. 2015, 52, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.T.; Tang, H.Y.; Han, Y.S.; Liu, H.P.; Huang, S.F.; Chien, C.H.; Shyy, J.; Chiu, J.J.; Chen, X. Downregulation of Signaling-active IGF-1 by Dipeptidyl Peptidase IV (DPP-IV). Int. J. Biomed. Sci. 2010, 6, 301–309. [Google Scholar] [PubMed]
- Shin, J.; Fukuhara, A.; Onodera, T.; Kita, S.; Yokoyama, C.; Otsuki, M.; Shimomura, I. SDF-1 Is an Autocrine Insulin-Desensitizing Factor in Adipocytes. Diabetes 2018, 67, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Feickert, M.; Burckhardt, B.B. Substance P in cardiovascular diseases—A bioanalytical review. Clin. Chim. Acta 2019, 495, 501–506. [Google Scholar] [CrossRef]
- Ahmad, S.; Wang, L.; Ward, P.E. Dipeptidyl(amino)peptidase IV and aminopeptidase M metabolize circulating substance P in vivo. J. Pharmacol. Exp. Ther. 1992, 260, 1257–1261. [Google Scholar]
- Guieu, R.; Fenouillet, E.; Devaux, C.; Fajloun, Z.; Carrega, L.; Sabatier, J.M.; Sauze, N.; Marguet, D. CD26 modulates nociception in mice via its dipeptidyl-peptidase IV activity. Behav. Brain Res. 2006, 166, 230–235. [Google Scholar] [CrossRef]
- Ramalho, R.; Almeida, J.; Beltrão, M.; Pirraco, A.; Costa, R.; Sokhatska, O.; Guardão, L.; Palmares, C.; Guimarães, J.T.; Delgado, L.; et al. Substance P antagonist improves both obesity and asthma in a mouse model. Allergy 2013, 68, 48–54. [Google Scholar] [CrossRef]
- Dubon, M.J.; Byeon, Y.; Park, K.S. Substance P enhances the activation of AMPK and cellular lipid accumulation in 3T3 L1 cells in response to high levels of glucose. Mol. Med. Rep. 2015, 12, 8048–8054. [Google Scholar] [CrossRef]
- Um, J.; Jung, N.; Kim, D.; Choi, S.; Lee, S.H.; Son, Y.; Park, K.S. Substance P preserves pancreatic β-cells in type 1 and type 2 diabetic mice. Biochem. Biophys. Res. Commun. 2018, 499, 960–966. [Google Scholar] [CrossRef]
- Sell, H.; Blüher, M.; Klöting, N.; Schlich, R.; Willems, M.; Ruppe, F.; Knoefel, W.T.; Dietrich, A.; Fielding, B.A.; Arner, P.; et al. Adipose dipeptidyl peptidase-4 and obesity: Correlation with insulin resistance and depot-specific release from adipose tissue in vivo and in vitro. Diabetes Care 2013, 36, 4083–4090. [Google Scholar] [CrossRef]
- Rohmann, N.; Schlicht, K.; Geisler, C.; Hollstein, T.; Knappe, C.; Krause, L.; Hagen, S.; Beckmann, A.; Seoudy, A.K.; Wietzke-Braun, P.; et al. Circulating sDPP-4 is Increased in Obesity and Insulin Resistance but Is Not Related to Systemic Metabolic Inflammation. J. Clin. Endocrinol. Metab. 2021, 106, e592–e601. [Google Scholar] [CrossRef] [PubMed]
- Rüter, J.; Hoffmann, T.; Demuth, H.U.; Moschansky, P.; Klapp, B.F.; Hildebrandt, M. Evidence for an interaction between leptin, T cell costimulatory antigens CD28, CTLA-4 and CD26 (dipeptidyl peptidase IV) in BCG-induced immune responses of leptin- and leptin receptor-deficient mice. Biol. Chem. 2004, 385, 537–541. [Google Scholar] [CrossRef]
- Stephan, M.; Radicke, A.; Leutloff, S.; Schmiedl, A.; Pabst, R.; von Hörsten, S.; Dettmer, S.; Lotz, J.; Nave, H. Dipeptidyl peptidase IV (DPP4)-deficiency attenuates diet-induced obesity in rats: Possible implications for the hypothalamic neuropeptidergic system. Behav. Brain Res. 2011, 216, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Romacho, T.; Sell, H.; Indrakusuma, I.; Roehrborn, D.; Castañeda, T.R.; Jelenik, T.; Markgraf, D.; Hartwig, S.; Weiss, J.; Al-Hasani, H.; et al. DPP4 deletion in adipose tissue improves hepatic insulin sensitivity in diet-induced obesity. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E590–E599. [Google Scholar] [CrossRef] [PubMed]
- Röhrborn, D.; Brückner, J.; Sell, H.; Eckel, J. Reduced DPP4 activity improves insulin signaling in primary human adipocytes. Biochem. Biophys. Res. Commun. 2016, 471, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Karl, T.; Hoffmann, T.; Pabst, R.; von Hörsten, S. Extreme reduction of dipeptidyl peptidase IV activity in F344 rat substrains is associated with various behavioral differences. Physiol. Behav. 2003, 80, 123–134. [Google Scholar] [CrossRef]
- Pospisilik, J.A.; Martin, J.; Doty, T.; Ehses, J.A.; Pamir, N.; Lynn, F.C.; Piteau, S.; Demuth, H.U.; McIntosh, C.H.; Pederson, R.A. Dipeptidyl peptidase IV inhibitor treatment stimulates beta-cell survival and islet neogenesis in streptozotocin-induced diabetic rats. Diabetes 2003, 52, 741–750. [Google Scholar] [CrossRef]
- Kohno, D.; Furusawa, K.; Kitamura, T. Anagliptin suppresses diet-induced obesity through enhancing leptin sensitivity and ameliorating hyperphagia in high-fat high-sucrose diet fed mice. Endocr. J. 2020, 67, 523–529. [Google Scholar] [CrossRef]
- Mietlicki-Baase, E.G.; McGrath, L.E.; Koch-Laskowski, K.; Krawczyk, J.; Pham, T.; Lhamo, R.; Reiner, D.J.; Hayes, M.R. Hindbrain DPP-IV inhibition improves glycemic control and promotes negative energy balance. Physiol. Behav. 2017, 173, 9–14. [Google Scholar] [CrossRef]
- Vella, A.; Bock, G.; Giesler, P.D.; Burton, D.B.; Serra, D.B.; Saylan, M.L.; Deacon, C.F.; Foley, J.E.; Rizza, R.A.; Camilleri, M. The effect of dipeptidyl peptidase-4 inhibition on gastric volume, satiation and enteroendocrine secretion in type 2 diabetes: A double-blind, placebo-controlled crossover study. Clin. Endocrinol. 2008, 69, 737–744. [Google Scholar] [CrossRef]
- Rotondo, A.; Masuy, I.; Verbeure, W.; Biesiekierski, J.R.; Deloose, E.; Tack, J. Randomised clinical trial: The DPP-4 inhibitor, vildagliptin, inhibits gastric accommodation and increases glucagon-like peptide-1 plasma levels in healthy volunteers. Aliment. Pharmacol. Ther. 2019, 49, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Heruc, G.A.; Horowitz, M.; Deacon, C.F.; Feinle-Bisset, C.; Rayner, C.K.; Luscombe-Marsh, N.; Little, T.J. Effects of dipeptidyl peptidase IV inhibition on glycemic, gut hormone, triglyceride, energy expenditure, and energy intake responses to fat in healthy males. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E830–E837. [Google Scholar] [CrossRef]
- Maloney, A.; Rosenstock, J.; Fonseca, V. A Model-Based Meta-Analysis of 24 Antihyperglycemic Drugs for Type 2 Diabetes: Comparison of Treatment Effects at Therapeutic Doses. Clin. Pharmacol. Ther. 2019, 105, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Waldrop, G.; Zhong, J.; Peters, M.; Goud, A.; Chen, Y.H.; Davis, S.N.; Mukherjee, B.; Rajagopalan, S. Incretin-based therapy in type 2 diabetes: An evidence based systematic review and meta-analysis. J. Diabetes Complicat. 2018, 32, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Janani, L.; Bamehr, H.; Tanha, K.; Mirzabeigi, P.; Montazeri, H.; Tarighi, P. Effects of Sitagliptin as Monotherapy and Add-On to Metformin on Weight Loss among Overweight and Obese Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis. Drug Res. 2021, 71, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Kirino, Y.; Sato, Y.; Kamimoto, T.; Kawazoe, K.; Minakuchi, K.; Nakahori, Y. Interrelationship of dipeptidyl peptidase IV (DPP4) with the development of diabetes, dyslipidaemia and nephropathy: A streptozotocin-induced model using wild-type and DPP4-deficient rats. J. Endocrinol. 2009, 200, 53–61. [Google Scholar] [CrossRef]
- Poucher, S.M.; Cheetham, S.; Francis, J.; Zinker, B.; Kirby, M.; Vickers, S.P. Effects of saxagliptin and sitagliptin on glycaemic control and pancreatic β-cell mass in a streptozotocin-induced mouse model of type 2 diabetes. Diabetes Obes. Metab. 2012, 14, 918–926. [Google Scholar] [CrossRef]
- Hamamoto, S.; Kanda, Y.; Shimoda, M.; Tatsumi, F.; Kohara, K.; Tawaramoto, K.; Hashiramoto, M.; Kaku, K. Vildagliptin preserves the mass and function of pancreatic β cells via the developmental regulation and suppression of oxidative and endoplasmic reticulum stress in a mouse model of diabetes. Diabetes Obes. Metab. 2013, 15, 153–163. [Google Scholar] [CrossRef]
- Ellis, S.L.; Moser, E.G.; Snell-Bergeon, J.K.; Rodionova, A.S.; Hazenfield, R.M.; Garg, S.K. Effect of sitagliptin on glucose control in adult patients with Type 1 diabetes: A pilot, double-blind, randomized, crossover trial. Diabet. Med. 2011, 28, 1176–1181. [Google Scholar] [CrossRef] [PubMed]
- Seelig, E.; Trinh, B.; Hanssen, H.; Schmid-Trucksäss, A.; Ellingsgaard, H.; Christ-Crain, M.; Donath, M.Y. Exercise and the dipeptidyl-peptidase IV inhibitor sitagliptin do not improve beta-cell function and glucose homeostasis in long-lasting type 1 diabetes-A randomised open-label study. Endocrinol. Diabetes Metab. 2019, 2, e00075. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, L.; Xiang, Y.; Liu, L.; Huang, G.; Long, Z.; Li, X.; Leslie, R.D.; Wang, X.; Zhou, Z. Dipeptidyl peptidase 4 inhibitor sitagliptin maintains β-cell function in patients with recent-onset latent autoimmune diabetes in adults: One year prospective study. J. Clin. Endocrinol. Metab. 2014, 99, E876–E880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Liang, H.; Liu, X.; Wang, X.; Cheng, Y.; Zhao, Y.; Liu, L.; Huang, G.; Wang, X.; Zhou, Z. Islet Function and Insulin Sensitivity in Latent Autoimmune Diabetes in Adults Taking Sitagliptin: A Randomized Trial. J. Clin. Endocrinol. Metab. 2021, 106, e1529–e1541. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, L.; Cheng. Y.; Zheng, P.; Hu, J.; Huang, G.; Zhou, Z. Altered T-cell subsets and transcription factors in latent autoimmune diabetes in adults taking sitagliptin, a dipeptidyl peptidase-4 inhibitor: A 1-year open-label randomized controlled trial. J. Diabetes Investig. 2019, 10, 375–382. [Google Scholar] [CrossRef]
- Farngren, J.; Persson, M.; Schweizer, A.; Foley, J.E.; Ahrén, B. Vildagliptin reduces glucagon during hyperglycemia and sustains glucagon counter regulation during hypoglycemia in type 1 diabetes. J. Clin. Endocrinol. Metab. 2012, 97, 3799–3806. [Google Scholar] [CrossRef] [PubMed]
- Schopman, J.E.; Hoekstra, J.B.L.; Frier, B.M.; Ackermans, M.T.; de Sonnaville, J.J.J.; Stades, A.M.; Zwertbroek, R.; Hartmann, B.; Holst, J.J.; Knop, F.K.; et al. Effects of sitagliptin on counter-regulatory and incretin hormones during acute hypoglycaemia in patients with type 1 diabetes: A randomized double-blind placebo-controlled crossover study. Diabetes Obes. Metab. 2015, 17, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.K.; Moser, E.G.; Bode, B.W.; Klaff, L.J.; Hiatt, W.R.; Beatson, C.; Snell-Bergeon, J.K. Effect of sitagliptin on post-prandial glucagon and GLP-1 levels in patients with type 1 diabetes: Investigator-initiated, double-blind, randomized, placebo-controlled trial. Endocr. Pract. 2013, 19, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Rosmaninho-Salgado, J.; Marques, A.P.; Estrada, M.; Santana, M.; Cortez, V.; Grouzmann, E.; Cavadas, C. Dipeptidyl-peptidase-IV by cleaving neuropeptide y induces lipid accumulation and PPAR-γ expression. Peptides 2012, 37, 49–54. [Google Scholar] [CrossRef]
- Frerker, N.; Raber, K.; Bode, F.; Skripuletz, T.; Nave, H.; Klemann, C.; Pabst, R.; Stephan, M.; Schade, J.; Brabant, G.; et al. Phenotyping of congenic dipeptidyl peptidase 4 (DP4) deficient Dark Agouti (DA) rats suggests involvement of DP4 in neuro-, endocrine, and immune functions. Clin. Chem. Lab. Med. 2009, 47, 275–287. [Google Scholar] [CrossRef]
- Kos, K.; Baker, A.R.; Jernas, M.; Harte, A.L.; Clapham, J.C.; O’Hare, J.P.; Carlsson, L.; Kumar, S.; McTernan, P.G. DPP-IV inhibition enhances the antilipolytic action of NPY in human adipose tissue. Diabetes Obes. Metab. 2009, 11, 285–292. [Google Scholar] [CrossRef]
- Kim, J.; Choi, Y.S.; Lim, S.; Yea, K.; Yoon, J.H.; Jun, D.J.; Ha, S.H.; Kim, J.W.; Kim, J.H.; Suh, P.G.; et al. Comparative analysis of the secretory proteome of human adipose stromal vascular fraction cells during adipogenesis. Proteomics 2010, 10, 394–405. [Google Scholar] [CrossRef]
- Rosenow, A.; Arrey, T.N.; Bouwman, F.G.; Noben, J.P.; Wabitsch, M.; Mariman, E.C.; Karas, M.; Renes, J. Identification of novel human adipocyte secreted proteins by using SGBS cells. J. Proteome Res. 2010, 9, 5389–5401. [Google Scholar] [CrossRef] [PubMed]
- Svensson, H.; Odén, B.; Edén, S.; Lönn, M. Adiponectin, chemerin, cytokines, and dipeptidyl peptidase 4 are released from human adipose tissue in a depot-dependent manner: An in vitro system including human serum albumin. BMC Endocr. Disord. 2014, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Zilleßen, P.; Celner, J.; Kretschmann, A.; Pfeifer, A.; Racké, K.; Mayer, P. Metabolic role of dipeptidyl peptidase 4 (DPP4) in primary human (pre)adipocytes. Sci. Rep. 2016, 6, 23074. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Sawazaki, H.; Takahashi, H.; Yeh, Y.S.; Jheng, H.F.; Nomura, W.; Ara, T.; Takahashi, N.; Seno, S.; Osato, N.; et al. The dipeptidyl peptidase-4 (DPP-4) inhibitor teneligliptin enhances brown adipose tissue function, thereby preventing obesity in mice. FEBS Open Bio. 2018, 8, 1782–1793. [Google Scholar] [CrossRef] [PubMed]
- Fukuda-Tsuru, S.; Kakimoto, T.; Utsumi, H.; Kiuchi, S.; Ishii, S. The novel dipeptidyl peptidase-4 inhibitor teneligliptin prevents high-fat diet-induced obesity accompanied with increased energy expenditure in mice. Eur. J. Pharmacol. 2014, 723, 207–215. [Google Scholar] [CrossRef]
- Shimasaki, T.; Masaki, T.; Mitsutomi, K.; Ueno, D.; Gotoh, K.; Chiba, S.; Kakuma, T.; Yoshimatsu, H. The dipeptidyl peptidase-4 inhibitor des-fluoro-sitagliptin regulates brown adipose tissue uncoupling protein levels in mice with diet-induced obesity. PLoS ONE 2013, 8, e63626. [Google Scholar] [CrossRef]
- Bouchard, L.; Faucher, G.; Tchernof, A.; Deshaies, Y.; Lebel, S.; Hould, F.S.; Marceau, P.; Vohl, M.C. Comprehensive genetic analysis of the dipeptidyl peptidase-4 gene and cardiovascular disease risk factors in obese individuals. Acta Diabetol. 2009, 46, 13–21. [Google Scholar] [CrossRef]
- Turcot, V.; Bouchard, L.; Faucher, G.; Tchernof, A.; Deshaies, Y.; Pérusse, L.; Bélisle, A.; Marceau, S.; Biron, S.; Lescelleur, O.; et al. DPP4 gene DNA methylation in the omentum is associated with its gene expression and plasma lipid profile in severe obesity. Obesity 2011, 19, 388–395. [Google Scholar] [CrossRef]
- Turcot, V.; Tchernof, A.; Deshaies, Y.; Pérusse, L.; Bélisle, A.; Marceau, P.; Hould, F.-S.; Lebel, S.; Vohl, M.-C. Comparison of the dipeptidyl peptidase-4 gene methylation levels between severely obese subjects with and without the metabolic syndrome. Diabetol. Metab. Syndr. 2013, 5, 4. [Google Scholar] [CrossRef]
- Shirakawa, J.; Fujii, H.; Ohnuma, K.; Sato, K.; Ito, Y.; Kaji, M.; Sakamoto, E.; Koganei, M.; Sasaki, H.; Nagashima, Y.; et al. Diet-induced adipose tissue inflammation and liver steatosis are prevented by DPP-4 inhibition in diabetic mice. Diabetes 2011, 60, 1246–1257. [Google Scholar] [CrossRef]
- Shirakawa, J.; Okuyama, T.; Kyohara, M.; Yoshida, E.; Togashi, Y.; Tajima, K.; Yamazaki, S.; Kaji, M.; Koganei, M.; Sasaki, H.; et al. DPP-4 inhibition improves early mortality, β cell function, and adipose tissue inflammation in db/db mice fed a diet containing sucrose and linoleic acid. Diabetol. Metab. Syndr. 2016, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrian, A.D.; Ma, Q.; Lindsay, J.W.; Leone, K.A.; Ma, K.; Coben, J.; Galkina, E.V.; Nadler, J.L. Dipeptidyl peptidase IV inhibitor sitagliptin reduces local inflammation in adipose tissue and in pancreatic islets of obese mice. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E410–E421. [Google Scholar] [CrossRef] [PubMed]
- Zhuge, F.; Ni, Y.; Nagashimada, M.; Nagata, N.; Xu, L.; Mukaida, N.; Kaneko, S.; Ota, T. DPP-4 Inhibition by Linagliptin Attenuates Obesity-Related Inflammation and Insulin Resistance by Regulating M1/M2 Macrophage Polarization. Diabetes 2016, 65, 2966–2979. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Yu, D.M.; McCaughan, G.W.; Gorrell, M.D. Extra-enzymatic roles of DPIV and FAP in cell adhesion and migration on collagen and fibronectin. Adv. Exp. Med. Biol. 2006, 575, 213–222. [Google Scholar] [CrossRef]
- Lessard, J.; Pelletier, M.; Biertho, L.; Biron, S.; Marceau, S.; Hould, F.S.; Lebel, S.; Moustarah, F.; Lescelleur, O.; Marceau, P.; et al. Characterization of dedifferentiating human mature adipocytes from the visceral and subcutaneous fat compartments: Fibroblast-activation protein alpha and dipeptidyl peptidase 4 as major components of matrix remodeling. PLoS ONE 2015, 10, e0122065. [Google Scholar] [CrossRef]
- Marques, A.P.; Cunha-Santos, J.; Leal, H.; Sousa-Ferreira, L.; Pereira de Almeida, L.; Cavadas, C.; Rosmaninho-Salgado, J. Dipeptidyl peptidase IV (DPP-IV) inhibition prevents fibrosis in adipose tissue of obese mice. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 403–413. [Google Scholar] [CrossRef]
- Jung, Y.A.; Choi, Y.K.; Jung, G.S.; Seo, H.Y.; Kim, H.S.; Jang, B.K.; Kim, J.G.; Lee, I.K.; Kim, M.K.; Park, K.G. Sitagliptin attenuates methionine/choline-deficient diet-induced steatohepatitis. Diabetes Res. Clin. Pract. 2014, 105, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Baumeier, C.; Schlüter, L.; Saussenthaler, S.; Laeger, T.; Rödiger, M.; Alaze, S.A.; Fritsche, L.; Häring, H.U.; Stefan, N.; Fritsche, A.; et al. Elevated hepatic DPP4 activity promotes insulin resistance and non-alcoholic fatty liver disease. Mol. Metab. 2017, 6, 1254–1263. [Google Scholar] [CrossRef]
- Ghorpade, D.S.; Ozcan, L.; Zheng, Z.; Nicoloro, S.M.; Shen, Y.; Chen, E.; Blüher, M.; Czech, M.P.; Tabas, I. Hepatocyte-secreted DPP4 in obesity promotes adipose inflammation and insulin resistance. Nature 2018, 555, 673–677. [Google Scholar] [CrossRef]
- Sinitsky, M.Y.; Dyleva, Y.A.; Uchasova, E.G.; Belik, E.V.; Yuzhalin, A.E.; Gruzdeva, O.V.; Matveeva, V.G.; Ponasenko, A.V. Adipokine gene expression in adipocytes isolated from different fat depots of coronary artery disease patients. Arch. Physiol. Biochem. 2022, 128, 261–269. [Google Scholar] [CrossRef]
- Shibasaki, I.; Nakajima, T.; Fukuda, T.; Hasegawa, T.; Ogawa, H.; Tsuchiya, G.; Takei, Y.; Tezuka, M.; Kato, T.; Kanazawa, Y.; et al. Serum and Adipose Dipeptidyl Peptidase 4 in Cardiovascular Surgery Patients: Influence of Dipeptidyl Peptidase 4 Inhibitors. J. Clin. Med. 2022, 11, 4333. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulos, A.S.; Antoniades, C. The role of epicardial adipose tissue in cardiac biology: Classic concepts and emerging roles. J. Physiol. 2017, 595, 3907–3917. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Goebel-Stengel, M.; Teuffel, P.; Hofmann, T.; Buße, P.; Kobelt, P.; Rose, M.; Klapp, B.F. Obese patients have higher circulating protein levels of dipeptidyl peptidase IV. Peptides 2014, 61, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Jang, B.K.; Song, M.K.; Kim, H.S.; Kim, M.K. Association between Serum Dipeptidyl Peptidase-4 Concentration and Obesity-related Factors in Health Screen Examinees. J. Obes. Metab. Syndr. 2017, 26, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Stenlid, R.; Manell, H.; Halldin, M.; Kullberg, J.; Ahlström, H.; Manukyan, L.; Weghuber, D.; Paulmichl, K.; Zsoldos, F.; Bergsten, P.; et al. High DPP-4 Concentrations in Adolescents Are Associated With Low Intact GLP-1. J. Clin. Endocrinol. Metab. 2018, 103, 2958–2966. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.H.; Huri, H.Z.; Al-Hamodi, Z.; Salem, S.D.; Muniandy, S. Serum levels of soluble CD26/dipeptidyl peptidase-IV in type 2 diabetes mellitus and its association with metabolic syndrome and therapy with antidiabetic agents in Malaysian subjects. PLoS ONE 2015, 10, e0140618. [Google Scholar] [CrossRef]
- Yamauchi, M.; Tsuchiya, T.; Takebayashi, K.; Inukai, T.; Hashimoto, K. Effects of short-term calorie restriction on circulating DPP-4/sCD26 concentrations and body composition in patients with type 2 diabetes. Diabetol. Int. 2021, 12, 286–292. [Google Scholar] [CrossRef]
- Malin, S.K.; Huang, H.; Mulya, A.; Kashyap, S.R.; Kirwan, J.P. Lower dipeptidyl peptidase-4 following exercise training plus weight loss is related to increased insulin sensitivity in adults with metabolic syndrome. Peptides 2013, 47, 142–147. [Google Scholar] [CrossRef]
- Barchetta, I.; Chiappetta, C.; Ceccarelli, V.; Cimini, F.A.; Bertoccini, L.; Gaggini, M.; Cristofano, C.D.; Silecchia, G.; Lenzi, A.; Leonetti, F.; et al. Angiopoietin-Like Protein 4 Overexpression in Visceral Adipose Tissue from Obese Subjects with Impaired Glucose Metabolism and Relationship with Lipoprotein Lipase. Int. J. Mol. Sci. 2020, 21, 7197. [Google Scholar] [CrossRef]
- Cimini, F.A.; Barchetta, I.; Ciccarelli, G.; Leonetti, F.; Silecchia, G.; Chiappetta, C.; Di Cristofano, C.; Capoccia, D.; Bertoccini, L.; Ceccarelli, V.; et al. Adipose tissue remodelling in obese subjects is a determinant of presence and severity of fatty liver disease. Diabetes Metab. Res. Rev. 2021, 37, e3358. [Google Scholar] [CrossRef]
- Barchetta, I.; Angelico, F.; Del Ben, M.; Di Martino, M.; Cimini, F.A.; Bertoccini, L.; Polimeni, L.; Catalano, C.; Fraioli, A.; Del Vescovo, R.; et al. Phenotypical heterogeneity linked to adipose tissue dysfunction in patients with Type 2 diabetes. Clin. Sci. 2016, 130, 1753–1762. [Google Scholar] [CrossRef] [PubMed]
- Barchetta, I.; Cimini, F.A.; Capoccia, D.; De Gioannis, R.; Porzia, A.; Mainiero, F.; Di Martino, M.; Bertoccini, L.; De Bernardinis, M.; Leonetti, F.; et al. WISP1 Is a Marker of Systemic and Adipose Tissue Inflammation in Dysmetabolic Subjects With or Without Type 2 Diabetes. J. Endocr. Soc. 2017, 1, 660–670. [Google Scholar] [CrossRef]
- Barchetta, I.; Ciccarelli, G.; Barone, E.; Cimini, F.A.; Ceccarelli, V.; Bertoccini, L.; Sentinelli, F.; Tramutola, A.; Del Ben, M.; Angelico, F.; et al. Greater circulating DPP4 activity is associated with impaired flow-mediated dilatation in adults with type 2 diabetes mellitus. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 1087–1094. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Nargis, T.; Tantia, O.; Ghosh, S.; Chakrabarti, P. Increased Plasma Dipeptidyl Peptidase-4 (DPP4) Activity Is an Obesity-Independent Parameter for Glycemic Deregulation in Type 2 Diabetes Patients. Front. Endocrinol. 2019, 10, 505. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Ozeki, N.; Terasawa, T.; Naruse, R.; Hara, K.; Suetsugu, M.; Takebayashi, K.; Shibazaki, M.; Haruki, K.; Morita, K.; et al. Serum level of soluble CD26/dipeptidyl peptidase-4 (DPP-4) predicts the response to sitagliptin, a DPP-4 inhibitor, in patients with type 2 diabetes controlled inadequately by metformin and/or sulfonylurea. Transl. Res. 2012, 159, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Patarrão, R.S.; Duarte, N.; Coelho, I.; Ward, J.; Ribeiro, R.T.; Meneses, M.J.; Andrade, R.; Costa, J.; Correia, I.; Boavida, J.M.; et al. Prediabetes blunts DPP4 genetic control of postprandial glycaemia and insulin secretion. Diabetologia 2022, 65, 861–871. [Google Scholar] [CrossRef]
- Rau, M.; Schmitt, J.; Berg, T.; Kremer, A.E.; Stieger, B.; Spanaus, K.; Bengsch, B.; Romero, M.R.; Marin, J.J.; Keitel, V.; et al. Serum IP-10 levels and increased DPPIV activity are linked to circulating CXCR3+ T cells in cholestatic HCV patients. PLoS ONE 2018, 13, e0208225. [Google Scholar] [CrossRef]
- Itou, M.; Kawaguchi, T.; Taniguchi, E.; Sata, M. Dipeptidyl peptidase-4: A key player in chronic liver disease. World J. Gastroenterol. 2013, 19, 2298–2306. [Google Scholar] [CrossRef]
- Balaban, Y.H.; Korkusuz, P.; Simsek, H.; Gokcan, H.; Gedikoglu, G.; Pinar, A.; Hascelik, G.; Asan, E.; Hamaloglu, E.; Tatar, G. Dipeptidyl peptidase IV (DDP IV) in NASH patients. Ann. Hepatol. 2007, 6, 242–250. [Google Scholar] [CrossRef]
- Williams, K.H.; Vieira De Ribeiro, A.J.; Prakoso, E.; Veillard, A.S.; Shackel, N.A.; Brooks, B.; Bu, Y.; Cavanagh, E.; Raleigh, J.; McLennan, S.V.; et al. Circulating dipeptidyl peptidase-4 activity correlates with measures of hepatocyte apoptosis and fibrosis in non-alcoholic fatty liver disease in type 2 diabetes mellitus and obesity: A dual cohort cross-sectional study. J. Diabetes 2015, 7, 809–819. [Google Scholar] [CrossRef]
- Zheng, T.; Chen, B.; Yang, L.; Hu, X.; Zhang, X.; Liu, H.; Qin, L. Association of plasma dipeptidyl peptidase-4 activity with non-alcoholic fatty liver disease in nondiabetic Chinese population. Metabolism 2017, 73, 125–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firneisz, G.; Varga, T.; Lengyel, G.; Fehér, J.; Ghyczy, D.; Wichmann, B.; Selmeci, L.; Tulassay, Z.; Rácz, K.; Somogyi, A. Serum dipeptidyl peptidase-4 activity in insulin resistant patients with non-alcoholic fatty liver disease: A novel liver disease biomarker. PLoS ONE 2010, 5, e12226. [Google Scholar] [CrossRef] [PubMed]
- Sagara, M.; Iijima, T.; Kase, M.; Kato, K.; Sakurai, S.; Tomaru, T.; Jojima, T.; Usui, I.; Aso, Y. Serum levels of soluble dipeptidyl peptidase-4 in type 2 diabetes are associated with severity of liver fibrosis evaluated by transient elastography (FibroScan) and the FAST (FibroScan-AST) score, a novel index of non-alcoholic steatohepatitis with significant fibrosis. J. Diabetes Complicat. 2021, 35, 107885. [Google Scholar] [CrossRef]
- Lin, D.S.; Lee, J.K.; Hung, C.S.; Chen, W.J. The efficacy and safety of novel classes of glucose-lowering drugs for cardiovascular outcomes: A network meta-analysis of randomised clinical trials. Diabetologia 2021, 64, 2676–2686. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barchetta, I.; Cimini, F.A.; Dule, S.; Cavallo, M.G. Dipeptidyl Peptidase 4 (DPP4) as A Novel Adipokine: Role in Metabolism and Fat Homeostasis. Biomedicines 2022, 10, 2306. https://doi.org/10.3390/biomedicines10092306
Barchetta I, Cimini FA, Dule S, Cavallo MG. Dipeptidyl Peptidase 4 (DPP4) as A Novel Adipokine: Role in Metabolism and Fat Homeostasis. Biomedicines. 2022; 10(9):2306. https://doi.org/10.3390/biomedicines10092306
Chicago/Turabian StyleBarchetta, Ilaria, Flavia Agata Cimini, Sara Dule, and Maria Gisella Cavallo. 2022. "Dipeptidyl Peptidase 4 (DPP4) as A Novel Adipokine: Role in Metabolism and Fat Homeostasis" Biomedicines 10, no. 9: 2306. https://doi.org/10.3390/biomedicines10092306
APA StyleBarchetta, I., Cimini, F. A., Dule, S., & Cavallo, M. G. (2022). Dipeptidyl Peptidase 4 (DPP4) as A Novel Adipokine: Role in Metabolism and Fat Homeostasis. Biomedicines, 10(9), 2306. https://doi.org/10.3390/biomedicines10092306