
Evaluation of CAR-T Cells’ Cytotoxicity against Modified Solid Tumor Cell Lines

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtaining CAR-T Cells
2.2. Obtaining a Solid Tumor Cell Line Expressing the Red Fluorescent Protein Katushka2S and CD19 Antigen
2.2.1. Obtaining Lentiviral Particles
2.2.2. Cell Transduction
2.3. Evaluation of CAR-T Cells’ Cytotoxicity against Generated CD19+ Tumor Cells Using xCELLigence Biosensor Analyzer
2.4. Evaluation of CAR-T Cells Efficacy against H522 and PC-3M Tumor Cells
2.5. Evaluation of the Effectiveness of CAR-T Cells against 3D Tumor Cell Cultures
2.6. Multiplex Analysis of Supernatants for Cytokines
3. Results
3.1. Preparation and Phenotyping of CAR-T Cells
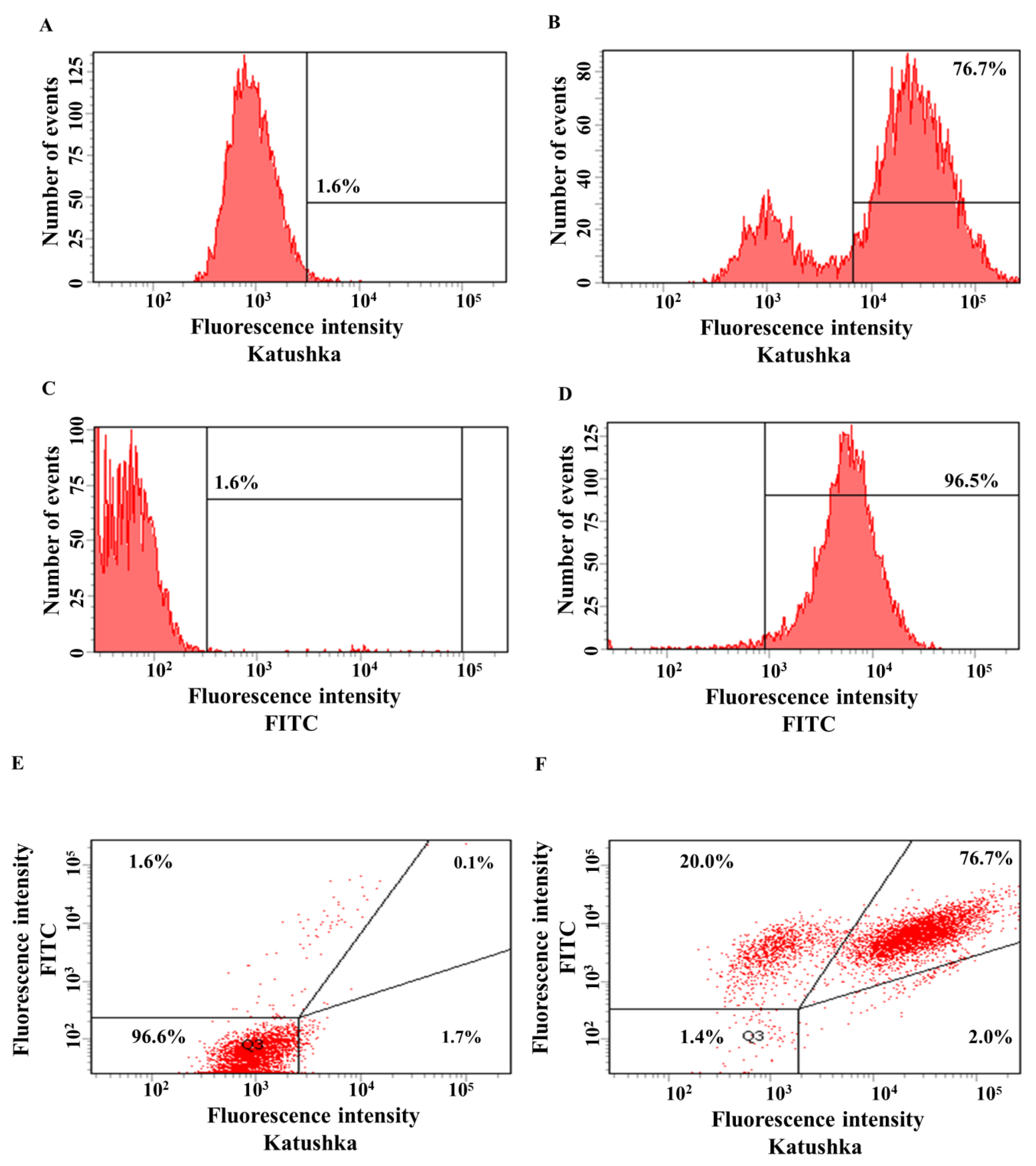
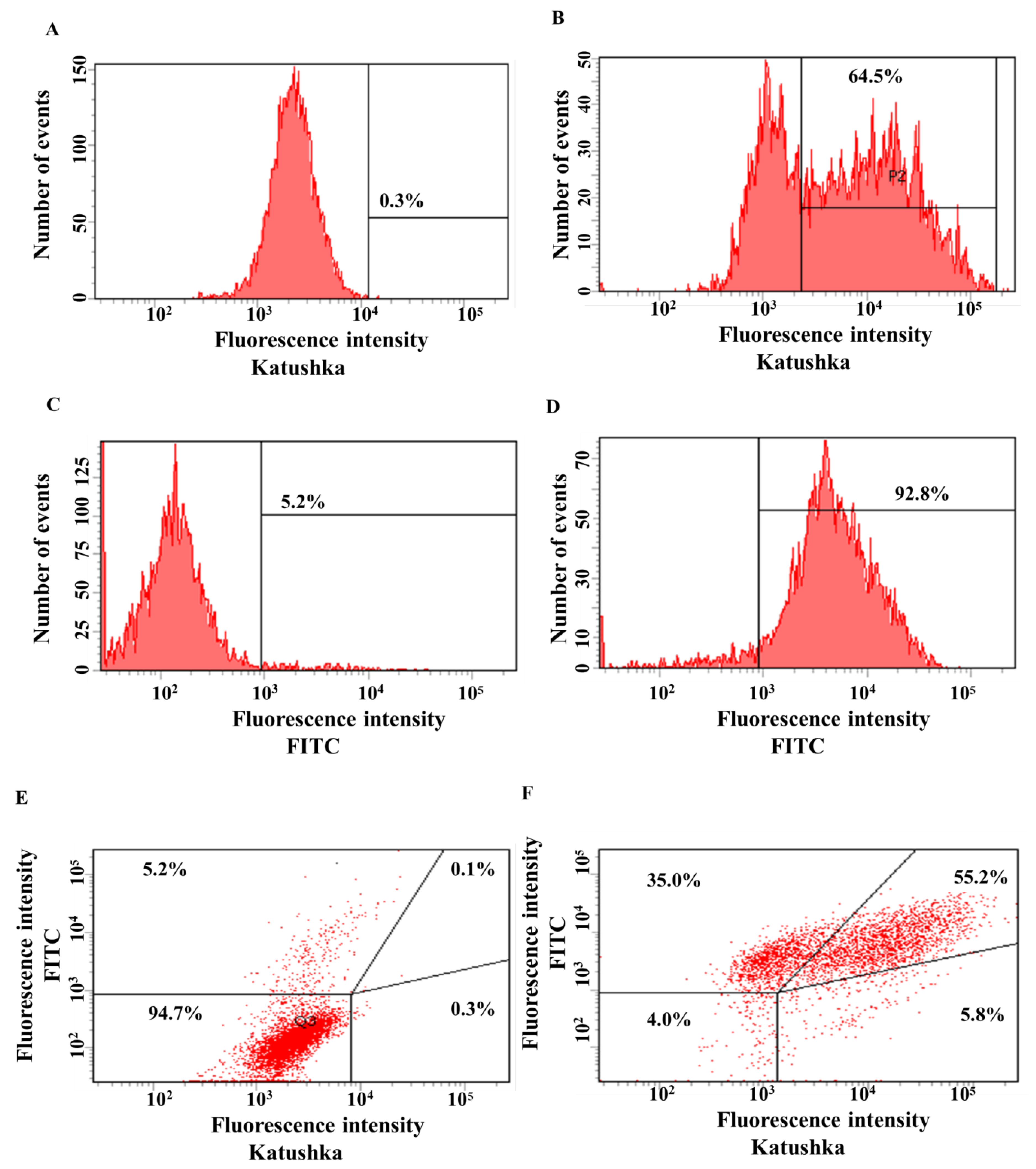
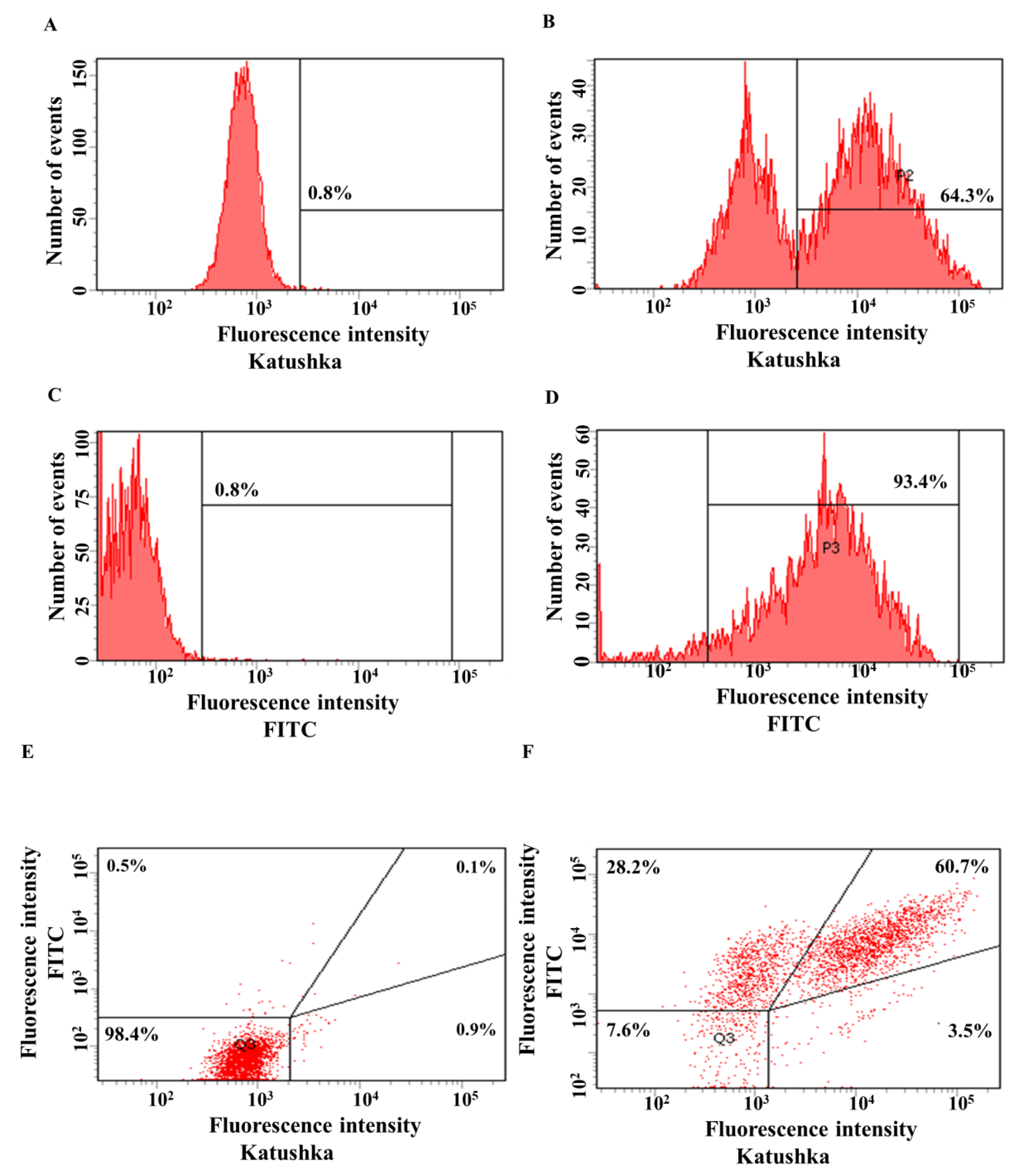
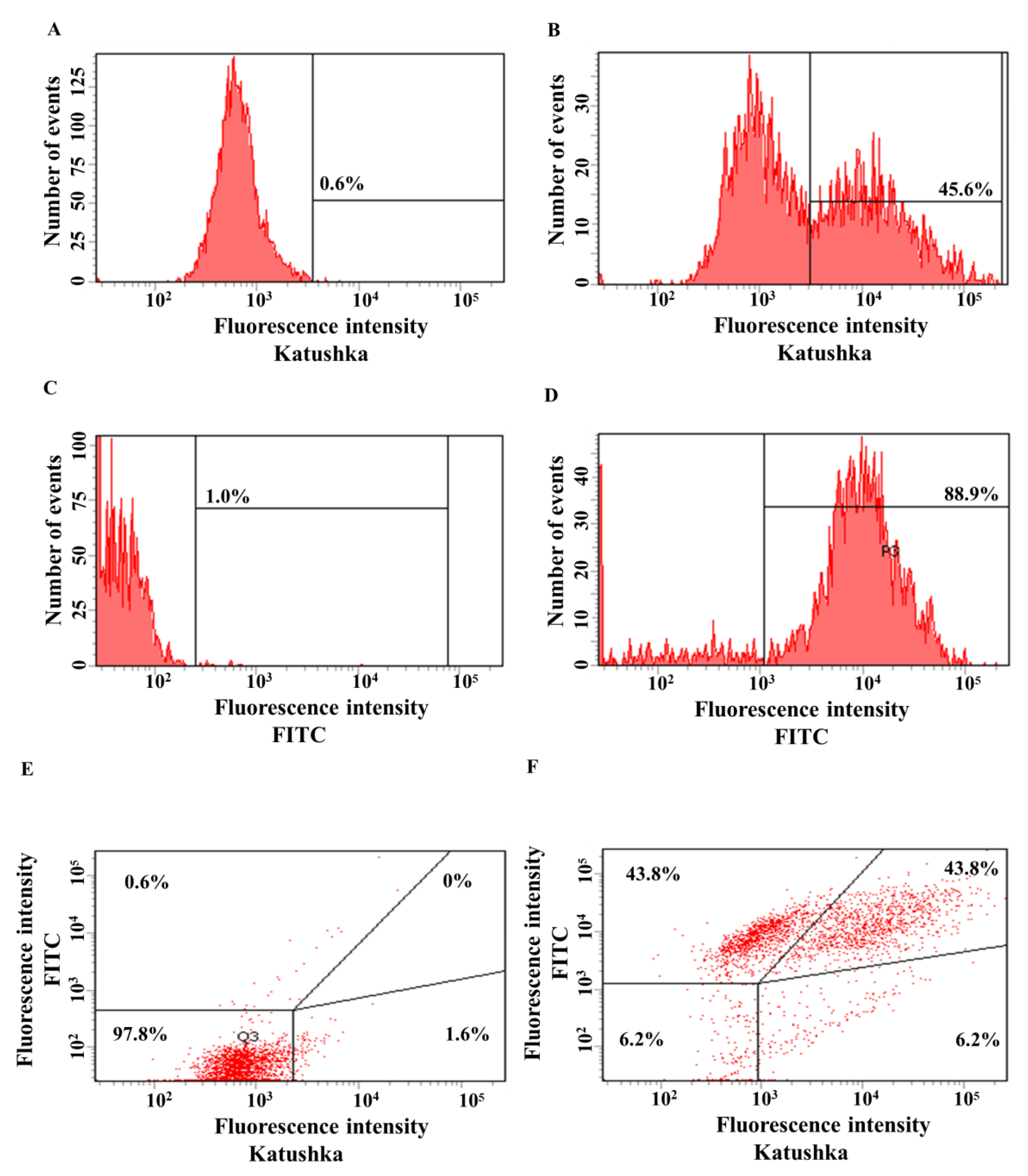
3.2. Generating Solid Tumor Cell Lines Expressing Red Fluorescent Protein Katushka2S and CD19 Antigen
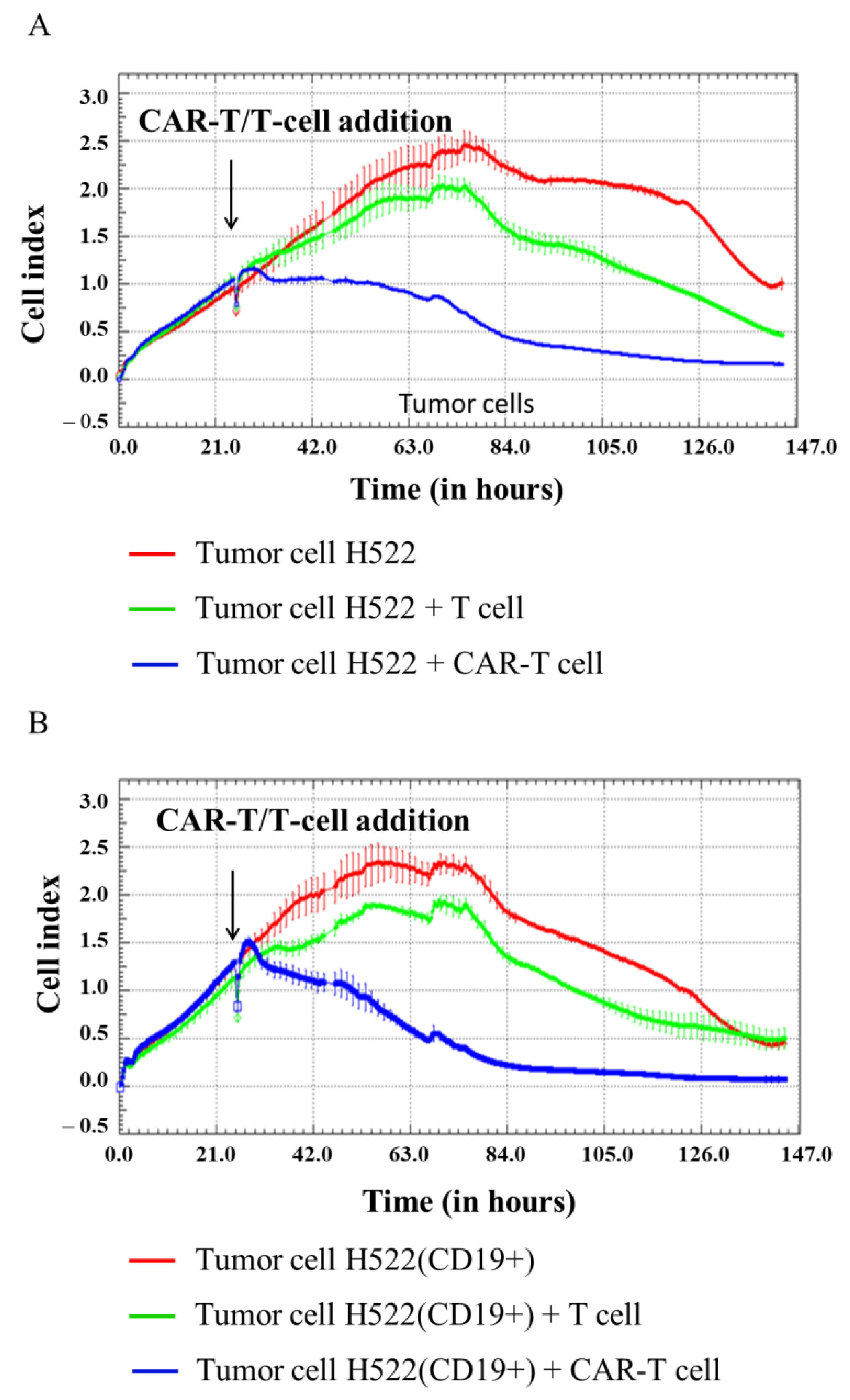
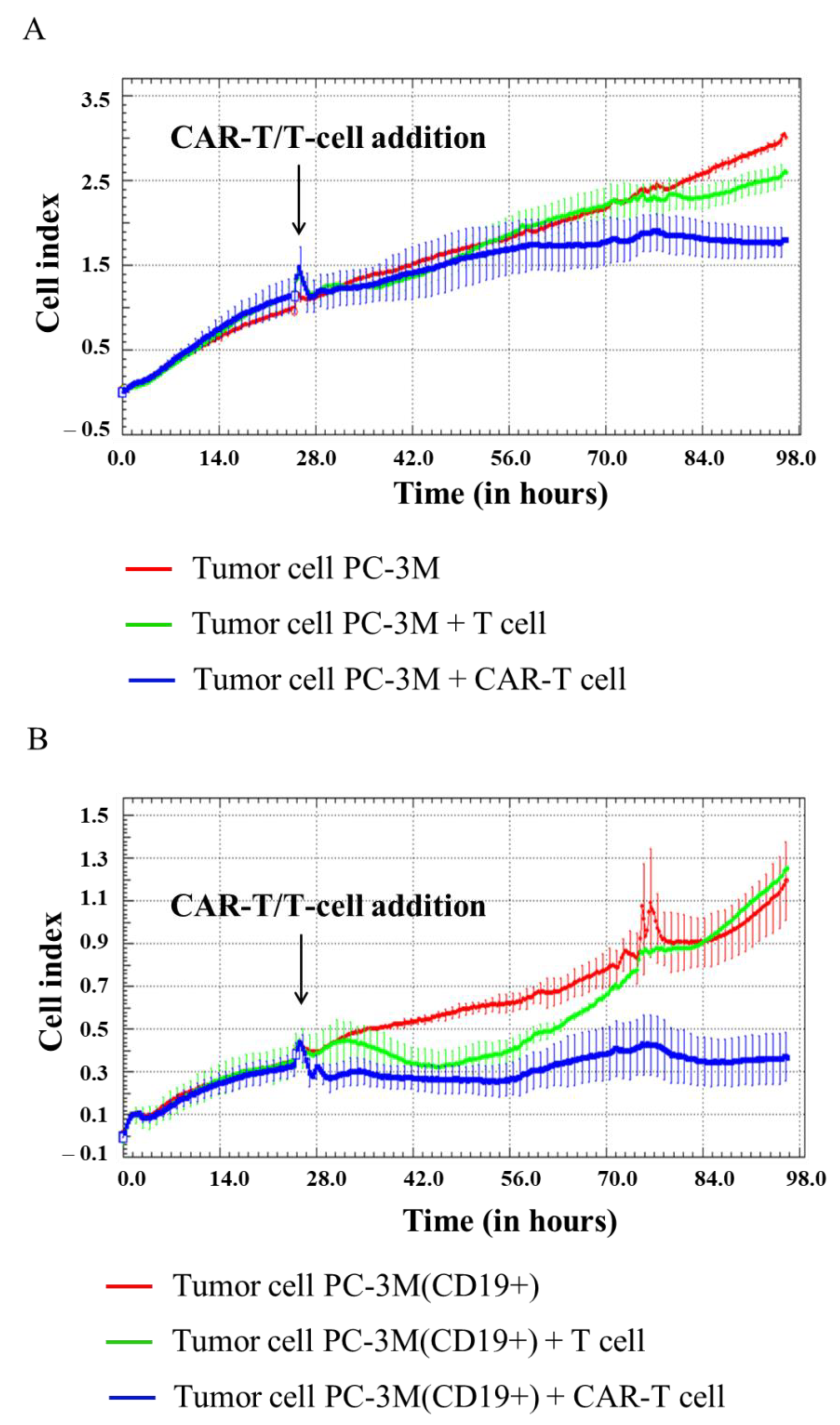
3.3. Evaluation of the Cytotoxicity of CAR-T Cells against Modified CD19+ Solid Tumor Cells Using xCELLigence Real-Time Cell Analyzer
3.4. Evaluation of the Effectiveness of CAR-T Cells against Tumor Cell Lines
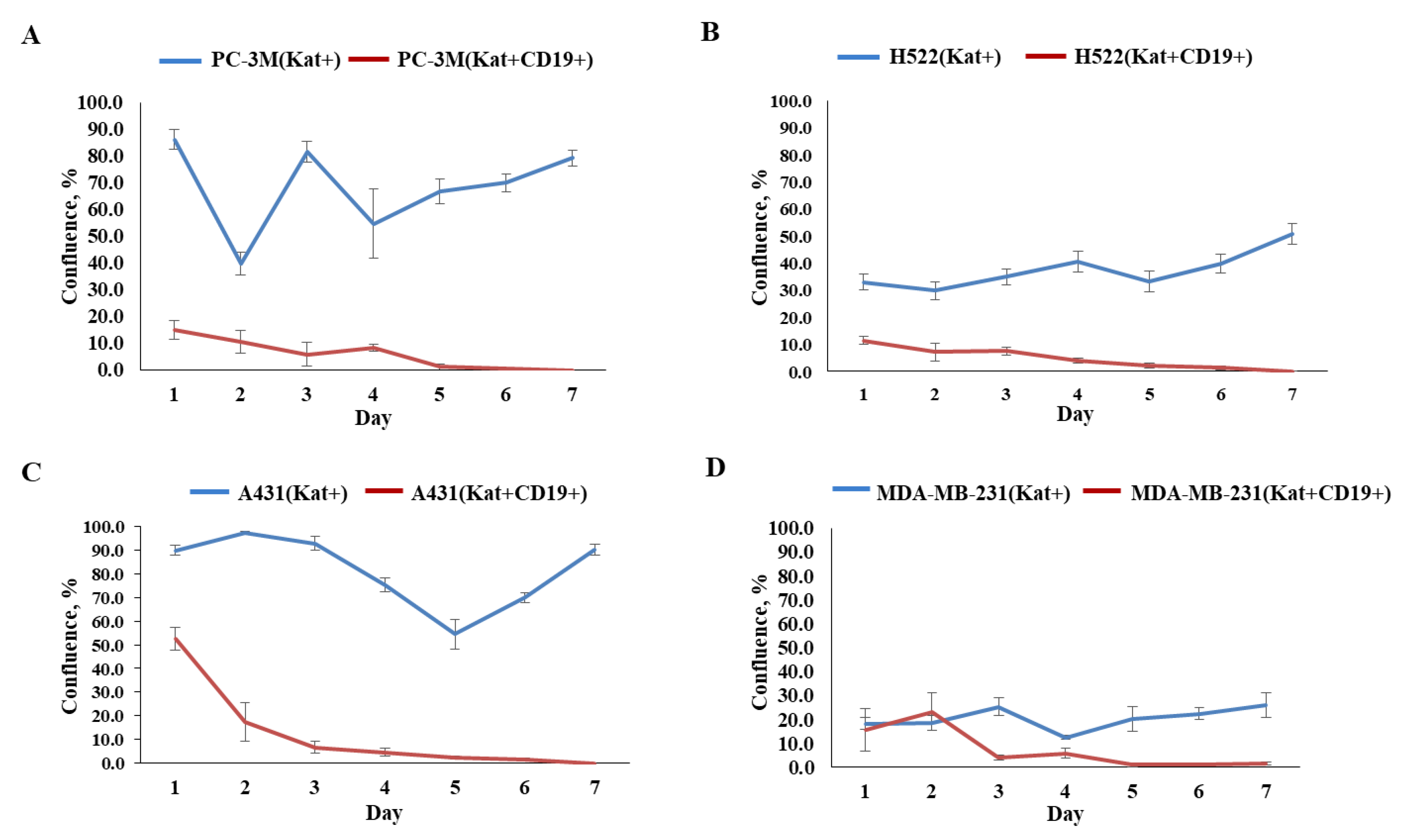
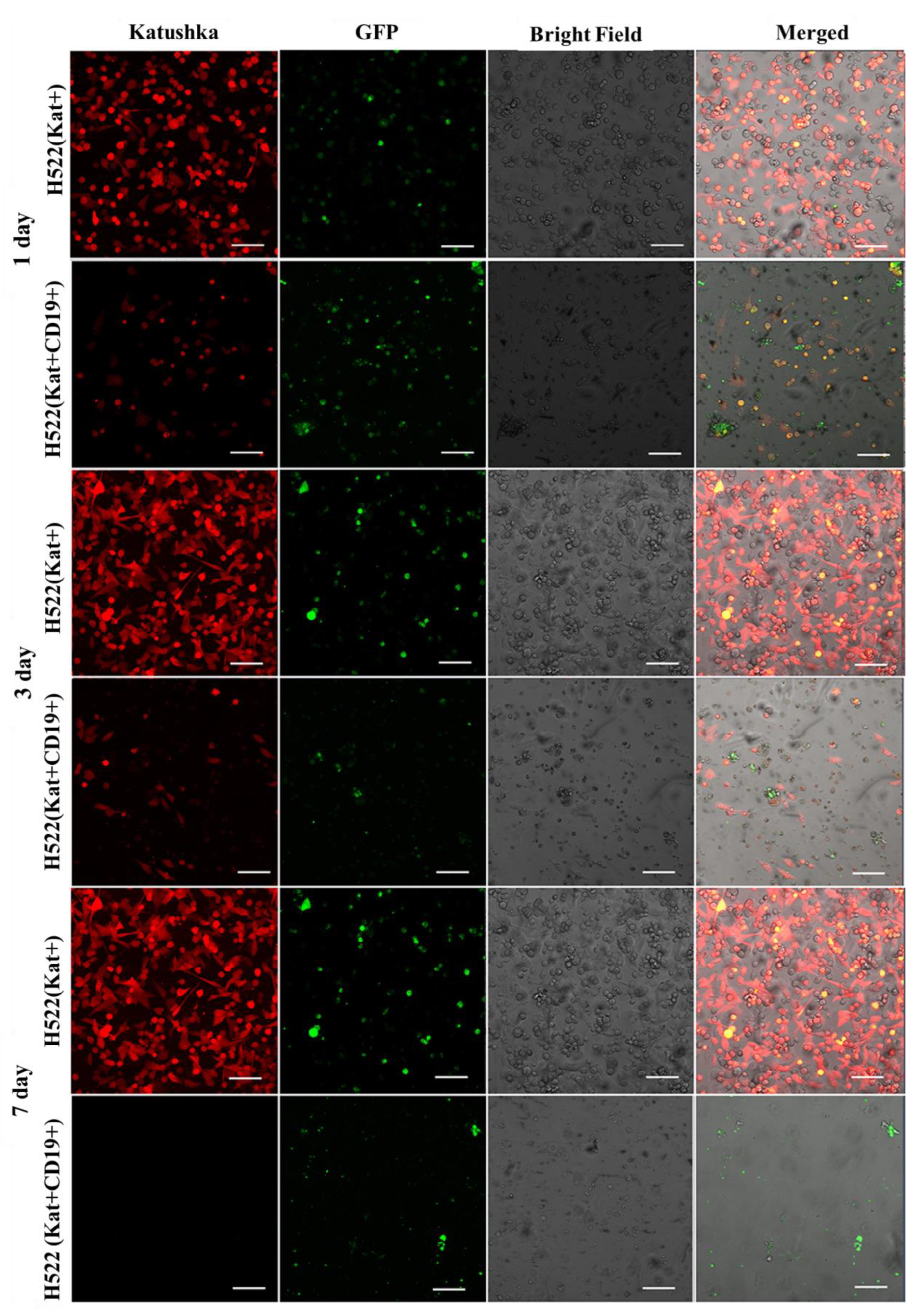
3.5. Evaluation of the Efficacy of CAR-T Cells against 3D Tumor Cell Cultures
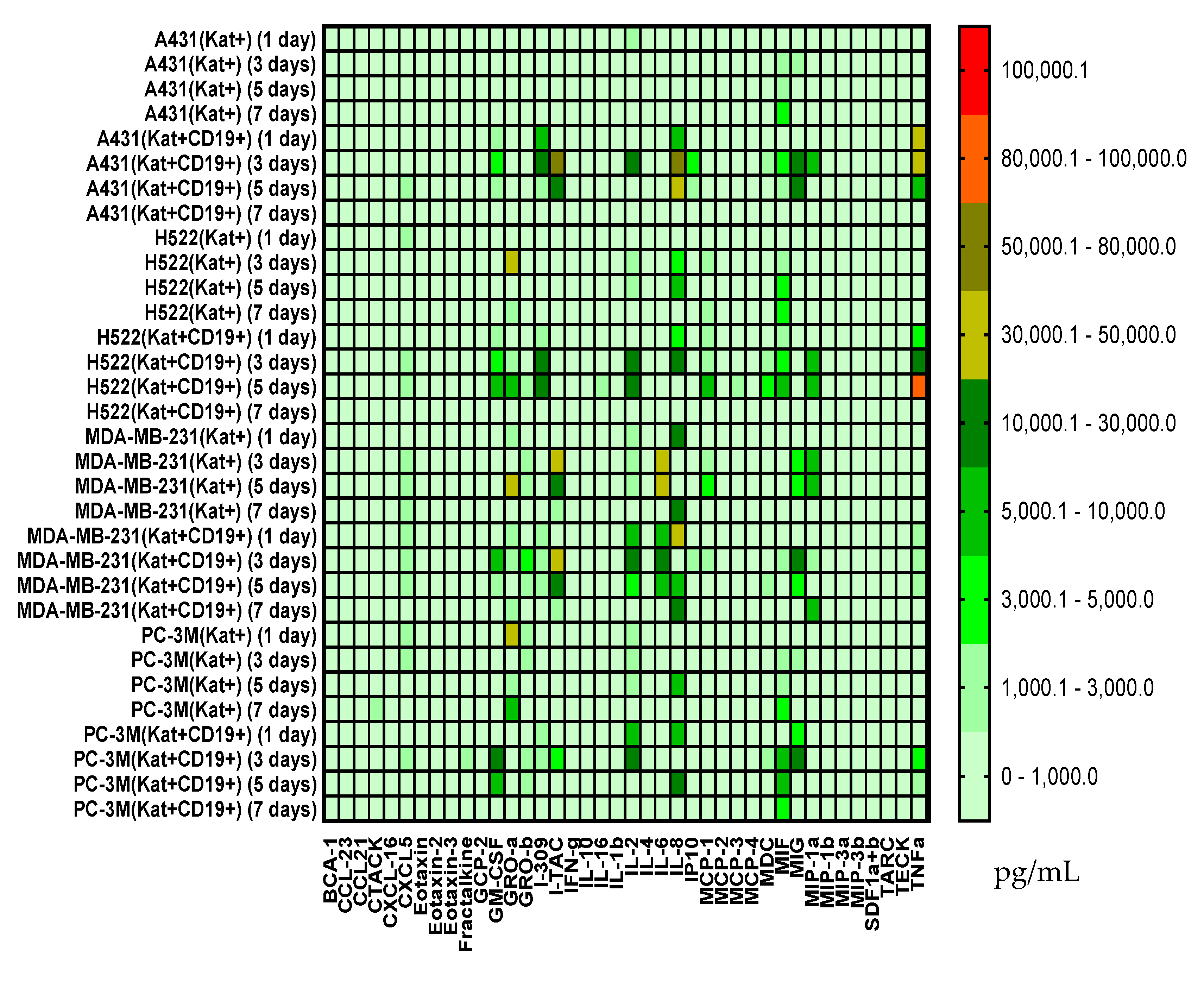
3.6. Evaluation of Cytokines and Chemokines in Supernatants via Multiplex Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Titov, A.; Zmievskaya, E.; Ganeeva, I.; Valiullina, A.; Petukhov, A.; Rakhmatullina, A.; Miftakhova, R.; Fainshtein, M.; Rizvanov, A.; Bulatov, E. Adoptive Immunotherapy beyond CAR T-Cells. Cancers 2021, 13, 743. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, M.; Du Rusquec, P.; Romano, E. Current Clinical Evidence and Potential Solutions to Increase Benefit of CAR T-Cell Therapy for Patients with Solid Tumors. Oncoimmunology 2020, 9, 1777064. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.; Wickman, E.; DeRenzo, C.; Gottschalk, S. CAR T Cell Therapy for Solid Tumors: Bright Future or Dark Reality? Mol. Ther. 2020, 28, 2320–2339. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Li, R.; Meng, L.; Han, Z.; Hong, Z. Macrophage, the potential key mediator in CAR-T related CRS. Exp. Hematol. Oncol. 2020, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Scholler, J.; Schalk, D.L.; June, C.H.; Lum, L.G. Enhanced cytotoxicity against solid tumors by bispecific antibody-armed CD19 CAR T cells: A proof-of-concept study. J. Cancer Res. Clin. Oncol. 2020, 146, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Donnadieu, E.; Dupré, L.; Pinho, L.G.; Cotta-De-Almeida, V. Surmounting the obstacles that impede effective CAR T cell trafficking to solid tumors. J. Leukoc. Biol. 2020, 108, 1067–1079. [Google Scholar] [CrossRef] [PubMed]
- Petukhov, A.; Markova, V.; Motorin, D.; Titov, A.; Belozerova, N.; Gershovich, P.; Karabel’Skii, A.; Ivanov, R.; Zaikova, E.; Smirnov, E.; et al. Manufacturing of CD19 Specific CAR T-Cells and Evaluation of their Functional Activity in Vitro. Clin. oncohematology 2018, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zaytsev, D.V.; Zaikova, E.; Golovkin, A.; Bulatov, E.; Valiullina, A.; Mirgayazova, R.; Daks, A.; Zaritskey, A.; Petukhov, A. Granulocyte-Macrophage Colony-Stimulating Factor and CAR-T Technology for Solid Tumors in Experiment. Clin. Oncohematology 2020, 13, 115–122. [Google Scholar] [CrossRef]
- Cullen, S.P.; Brunet, M.; Martin, S.J. Granzymes in cancer and immunity. Cell Death Differ. 2010, 17, 616–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Yang, J.F.; Deng, B.P.; Zhao, X.J.; Zhang, X.; Lin, Y.H.; Wu, Y.N.; Deng, Z.L.; Zhang, Y.L.; Liu, S.H.; et al. High efficacy and safety of low-dose CD19-directed CAR-T cell therapy in 51 refractory or relapsed B acute lymphoblastic leukemia patients. Leukemia 2017, 31, 2587–2593. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, J.; Schüßler-Lenz, M.; Bondanza, A.; Buchholz, C.J. Clinical development of CAR T cells—Challenges and opportunities in translating innovative treatment concepts. EMBO Mol. Med. 2017, 9, 1183–1197. [Google Scholar] [CrossRef] [PubMed]
- Guedan, S.; Madar, A.; Casado-Medrano, V.; Shaw, C.E.; Wing, A.; Liu, F.; Young, R.M.; June, C.H.; Posey, A.D., Jr. Single residue in CD28-costimulated CAR-T cells limits long-term persistence and antitumor durability. J. Clin. Investig. 2020, 130, 3087–3097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagyu, S.; Mochizuki, H.; Yamashima, K.; Kubo, H.; Saito, S.; Tanaka, M.; Sakamoto, K.; Shimoi, A.; Nakazawa, Y. A lymphodepleted non-human primate model for the assessment of acute on-target and off-tumor toxicity of human chimeric antigen receptor-T cells. Clin. Transl. Immunol. 2021, 10, e1291. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, M.; Ohminami, H.; Arai, J.; Kasahara, Y.; Ishida, Y.; Fujita, S. Granule exocytosis, and not the Fas/Fas ligand system, is the main pathway of cytotoxicity mediated by alloantigen-specific CD4+ as well as CD8+ cytotoxic T lymphocytes in humans. Blood 2000, 95, 2352–2355. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, J.; Ruella, M.; Houot, R. Overcoming Intrinsic Resistance of Cancer Cells to CAR T-Cell Killing. Clin. Cancer Res. 2021, 27, 6298–6306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Jordan, K.R.; Schulte, B.; Purev, E. Characterization of clinical grade CD19 chimeric antigen receptor T cells produced using automated CliniMACS prodigy system. Drug Des. Dev. Ther. 2018, 12, 3343–3356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, K.A.; Hanafi, L.-A.; Li, D.; Gust, J.; Liles, W.C.; Wurfel, M.M.; López, J.A.; Chen, J.; Chung, D.; Harju-Baker, S.; et al. Kinetics and biomarkers of severe cytokine release syndrome after CD19 chimeric antigen receptor–modified T-cell therapy. Blood 2017, 130, 2295–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valiullina, A.K.; Zmievskaya, E.A.; Ganeeva, I.A.; Zhuravleva, M.N.; Garanina, E.E.; Rizvanov, A.A.; Petukhov, A.V.; Bulatov, E.R. Evaluation of CAR-T Cells’ Cytotoxicity against Modified Solid Tumor Cell Lines. Biomedicines 2023, 11, 626. https://doi.org/10.3390/biomedicines11020626
Valiullina AK, Zmievskaya EA, Ganeeva IA, Zhuravleva MN, Garanina EE, Rizvanov AA, Petukhov AV, Bulatov ER. Evaluation of CAR-T Cells’ Cytotoxicity against Modified Solid Tumor Cell Lines. Biomedicines. 2023; 11(2):626. https://doi.org/10.3390/biomedicines11020626
Chicago/Turabian StyleValiullina, Aigul Kh., Ekaterina A. Zmievskaya, Irina A. Ganeeva, Margarita N. Zhuravleva, Ekaterina E. Garanina, Albert A. Rizvanov, Alexey V. Petukhov, and Emil R. Bulatov. 2023. "Evaluation of CAR-T Cells’ Cytotoxicity against Modified Solid Tumor Cell Lines" Biomedicines 11, no. 2: 626. https://doi.org/10.3390/biomedicines11020626
APA StyleValiullina, A. K., Zmievskaya, E. A., Ganeeva, I. A., Zhuravleva, M. N., Garanina, E. E., Rizvanov, A. A., Petukhov, A. V., & Bulatov, E. R. (2023). Evaluation of CAR-T Cells’ Cytotoxicity against Modified Solid Tumor Cell Lines. Biomedicines, 11(2), 626. https://doi.org/10.3390/biomedicines11020626